
Polydatin Alleviates Diabetes-Induced Hyposalivation through Anti-Glycation Activity in db/db Mouse



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. In Vivo Experiment Design in Normoglycemic Mice
2.3. In Vivo Experiment Design in Hyperglycemic Mice
2.4. Collection of Saliva
2.5. Hematoxylin and Eosin (H&E) Staining
2.6. Immunohistochemical Staining
2.7. Oxidative Stress Assay in Salivary Gland
2.8. Periodic Acid-Schiff (PAS) Staining
2.9. TUNEL Assay
2.10. Statistical Analysis
3. Results
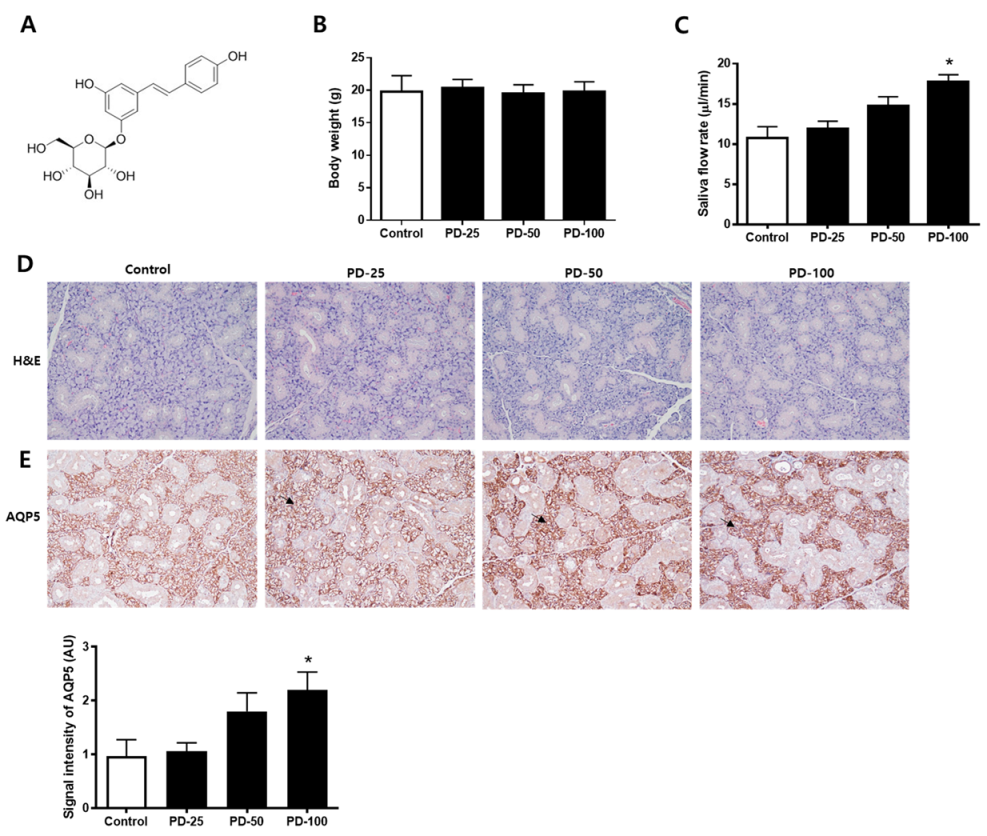
3.1. Increased Salivation in Normoglycemic Mice by Polydatin and Selection of the Oral Dose of Polydatin
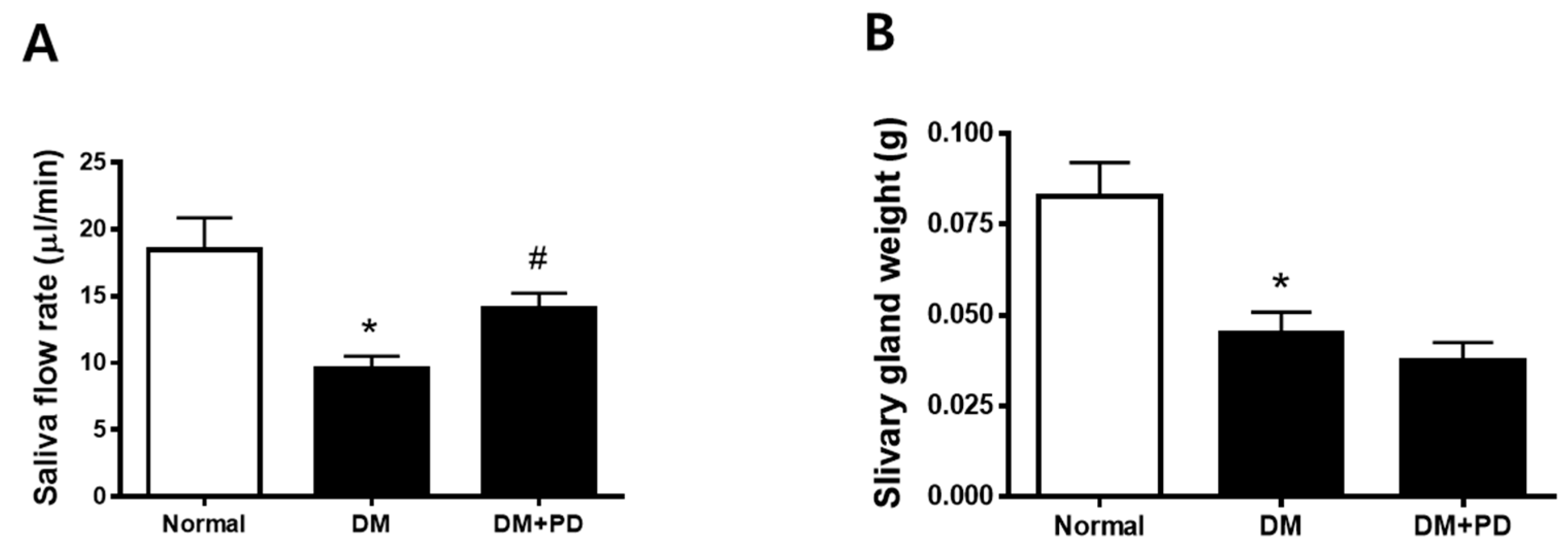
3.2. Polydatin Improves Hyposalivation in Type 2 Diabetic Mice
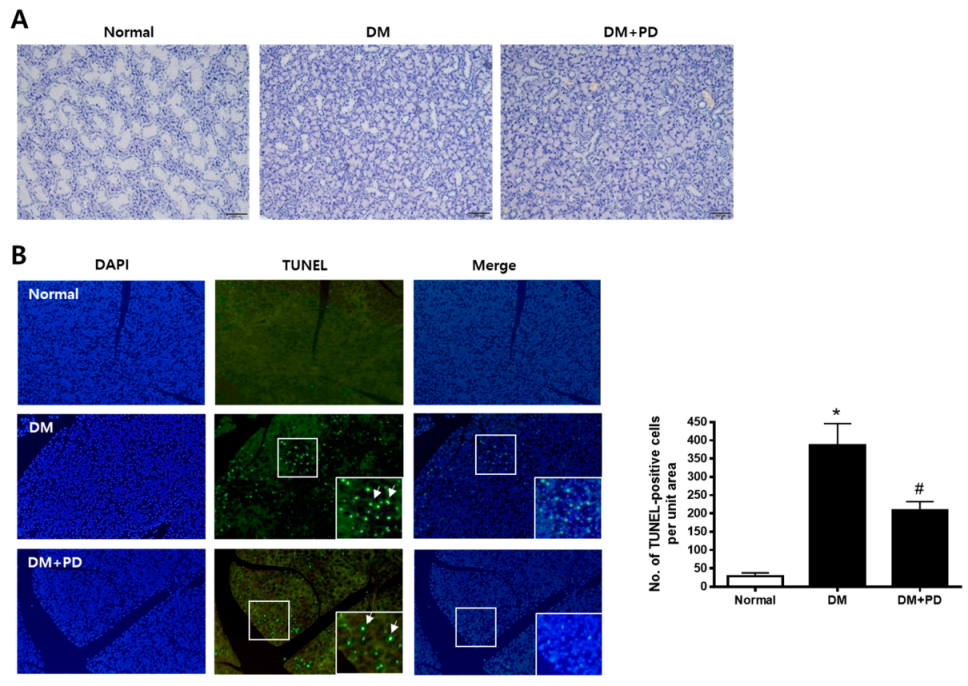
3.3. Polydatin Inhibits Apoptosis in the Submandibular Gland in Type 2 Diabetic Mice
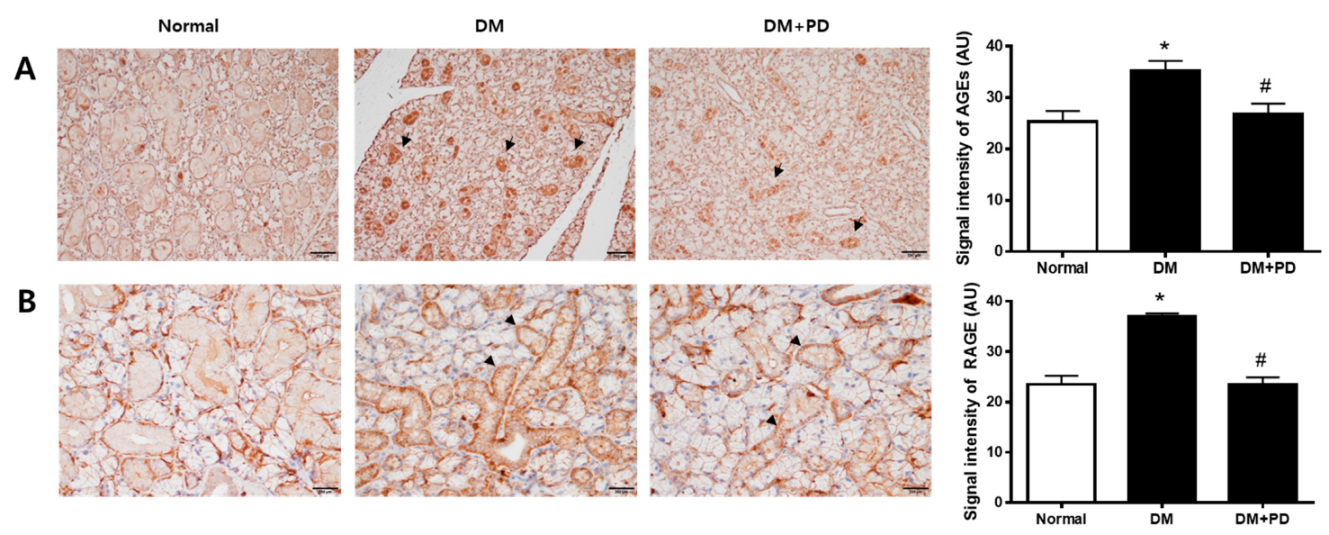
3.4. Polydatin Decreases AGEs Accumulations and RAGE Expression in the Submandibular Gland in Type 2 Diabetic Mice
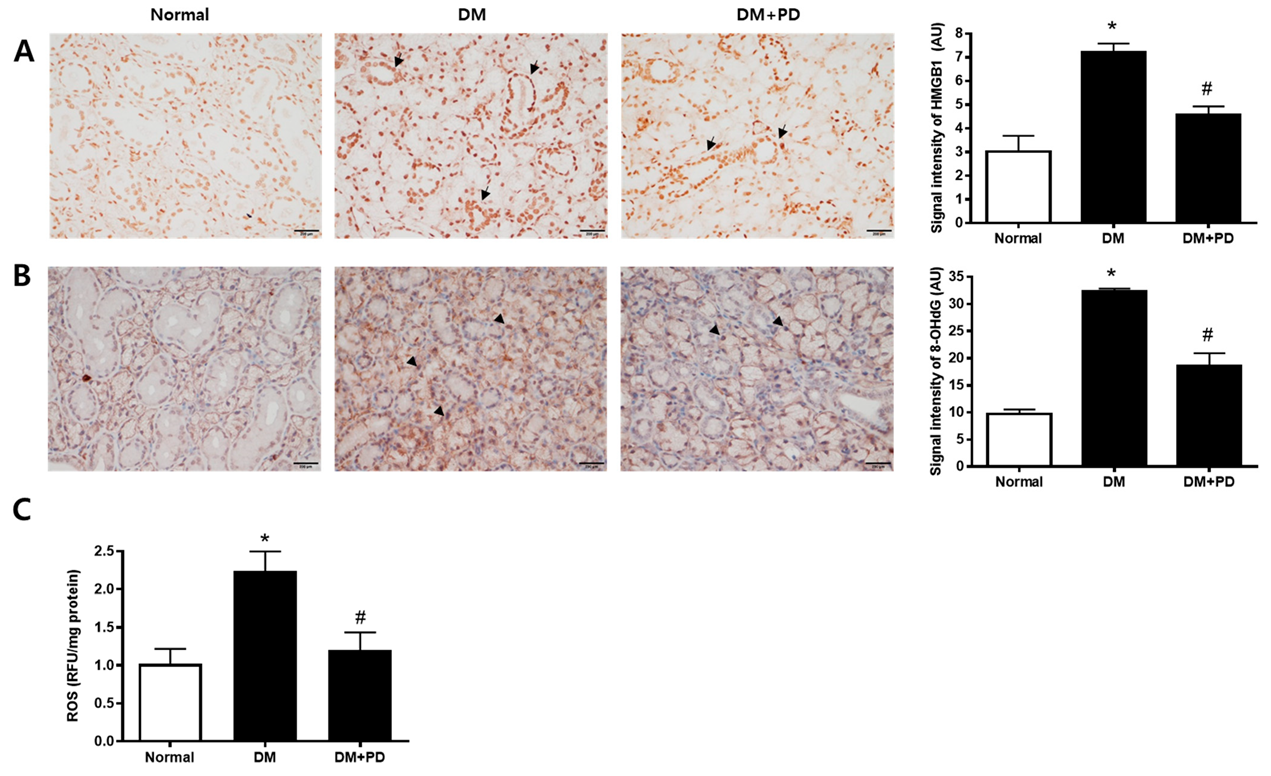
3.5. Polydatin Decreases the Expressions of HMGB1 and 8-OHdG in the Submandibular Gland in Type 2 Diabetic Mice
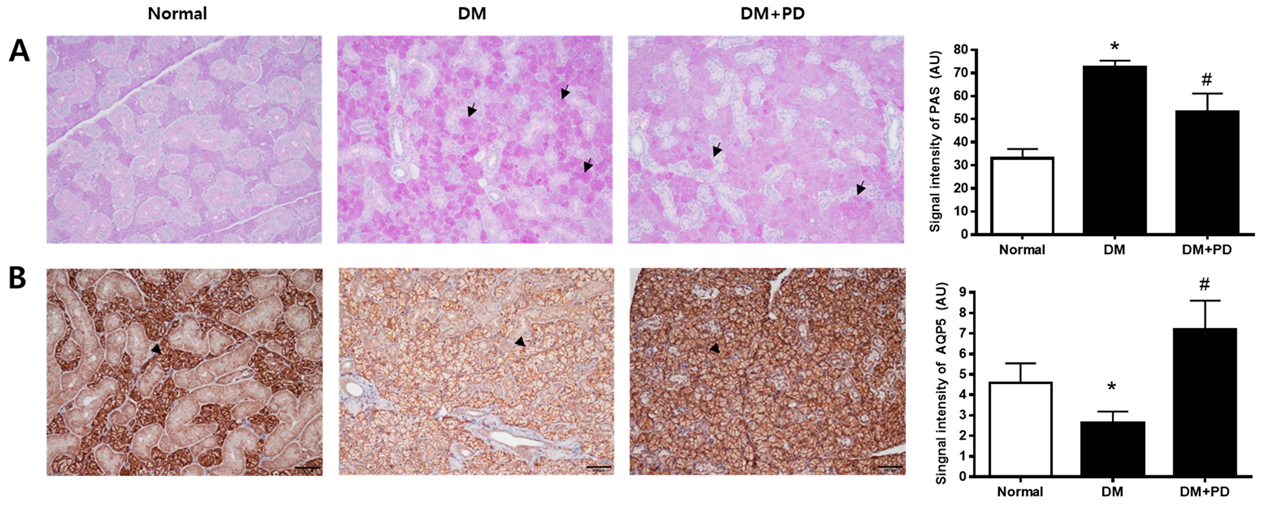
3.6. Polydatin Decreases Mucin Accumulation and the Increased AQP5 Channel in the Submandibular Gland in Type 2 Diabetic Mice
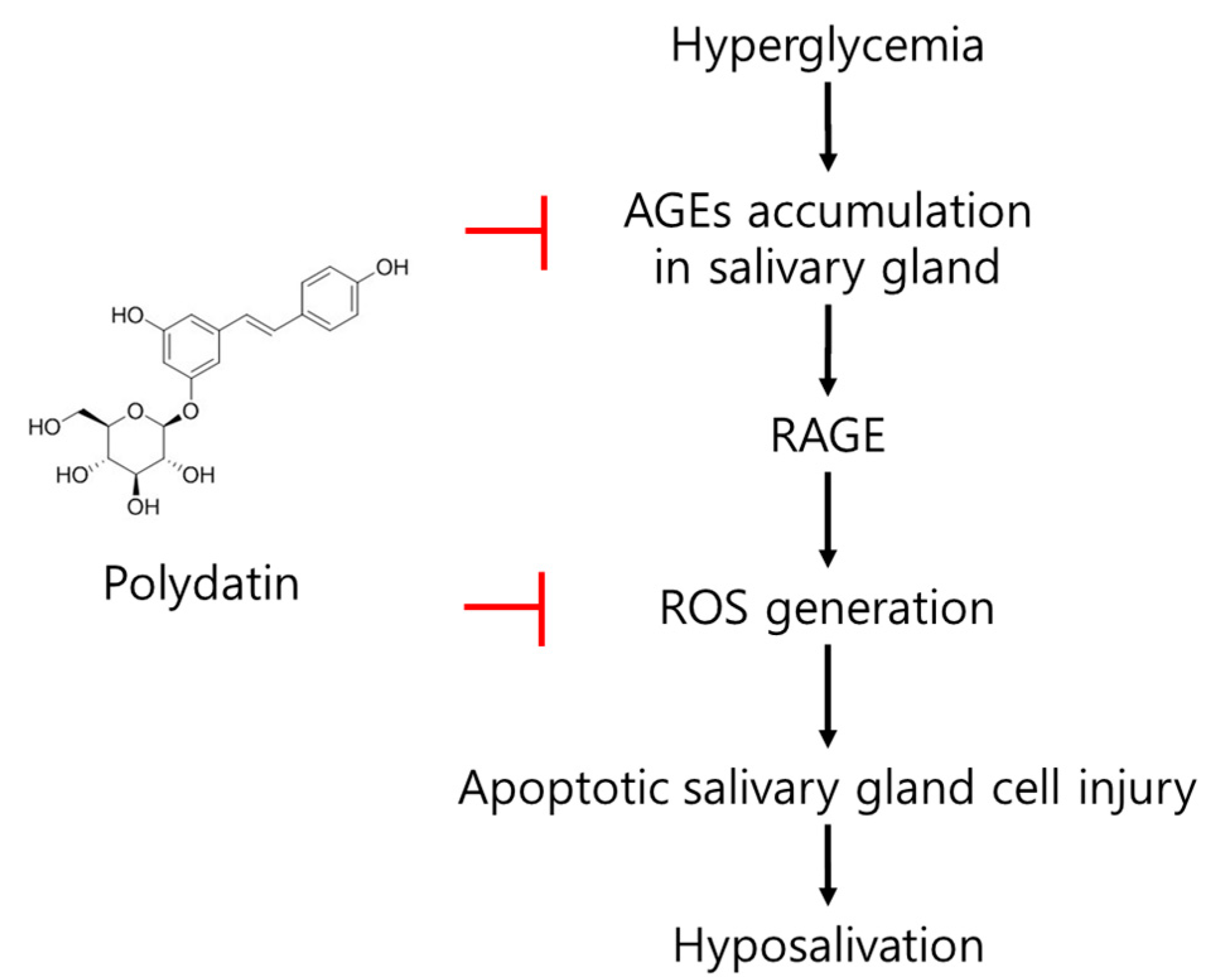
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miranda-Rius, J.; Brunet-Llobet, L.; Lahor-Soler, E.; Farré, M. Salivary secretory disorders, inducing drugs, and clinical management. Int. J. Med. Sci. 2015, 12, 811–824. [Google Scholar] [CrossRef] [Green Version]
- Proctor, G.B. The physiology of salivary secretion. Periodontology 2000 2016, 70, 11–25. [Google Scholar] [CrossRef] [Green Version]
- Mese, H.; Matsuo, R. Salivary secretion, taste and hyposalivation. J. Oral Rehabil. 2007, 34, 711–723. [Google Scholar] [CrossRef] [PubMed]
- van der Putten, G.J.; Brand, H.S.; Schols, J.M.; de Baat, C. The diagnostic suitability of a xerostomia questionnaire and the association between xerostomia, hyposalivation and medication use in a group of nursing home residents. Clin. Oral Investig. 2011, 15, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guggenheimer, J.; Moore, P.A. Xerostomia: Etiology, recognition and treatment. J. Am. Dent. Assoc. 2003, 134, 61–69. [Google Scholar] [CrossRef]
- Atkinson, J.C.; Grisius, M.; Massey, W. Salivary hypofunction and xerostomia: Diagnosis and treatment. Dent. Clin. N. Am. 2005, 49, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.A.; Guggenheimer, J.; Etzel, K.R.; Weyant, R.J.; Orchard, T. Type 1 diabetes mellitus, xerostomia, and salivary flow rates. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2001, 92, 281–291. [Google Scholar] [CrossRef]
- Feng, J.K.; Lu, Y.F.; Li, J.; Qi, Y.H.; Yi, M.L.; Ma, D.Y. Upregulation of salivary α2 macroglobulin in patients with type 2 diabetes mellitus. Genet. Mol. Res. 2015, 14, 2268–2274. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sreebny, L.M.; Yu, A.; Green, A.; Valdini, A. Xerostomia in diabetes mellitus. Diabetes Care 1992, 15, 900–904. [Google Scholar] [CrossRef]
- Rohani, B. Oral manifestations in patients with diabetes mellitus. World J. Diabetes 2019, 10, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Cicmil, S.; Mladenović, I.; Krunić, J.; Ivanović, D.; Stojanovic, N. Oral alterations in diabetes mdellitus. Balk. J. Dent. Med. 2018, 22, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Visvanathan, V.; Nix, P. Managing the patient presenting with xerostomia: A review. Int. J. Clin. Pract. 2010, 64, 404–407. [Google Scholar] [CrossRef]
- Bascones-Martínez, A.; Muñoz-Corcuera, M.; Bascones-Ilundain, C. Side effects of drugs on the oral cavity. Med. Clin. 2015, 144, 126–131. [Google Scholar] [CrossRef]
- Selvan, S.R.; Venugopalan, S. Effect of oral hypoglycemic drugs on salivary flow—A review. Int. J. Pharmtech. Res. 2013, 5, 1608–1610. [Google Scholar]
- Xie, X.; Peng, J.; Huang, K.; Huang, J.; Shen, X.; Liu, P.; Huang, H. Polydatin ameliorates experimental diabetes-induced fibronectin through inhibiting the activation of NF-κB signaling pathway in rat glomerular mesangial cells. Mol. Cell Endocrinol. 2012, 362, 183–193. [Google Scholar] [CrossRef]
- Du, Q.H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm. Biol. 2013, 51, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ye, J.; Li, J.; Chen, C.; Huang, J.; Liu, P.; Huang, H. Polydatin ameliorates lipid and glucose metabolism in type 2 diabetes mellitus by downregulating proprotein convertase subtilisin/kexin type 9 (PCSK9). Cardiovasc. Diabetol. 2016, 15, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şimşek, G.; Gürocak, S.; Karadaǧ, N.; Karabulut, A.B.; Demirtaş, E.; Karataş, E.; Pepele, E. Protective effects of resveratrol on salivary gland damage induced by total body irradiation in rats. Laryngoscope 2012, 122, 2743–2748. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yang, X.; Cai, J.; Ma, J.; Cheng, H.; Zhao, K.; Yang, L.; Cao, Y.; Qin, Q.; Zhang, C.; et al. Resveratrol attenuates radiation-induced salivary gland dysfunction in mice. Laryngoscope 2013, 123, 23–29. [Google Scholar] [CrossRef]
- Inoue, H.; Kishimoto, A.; Ushikoshi-Nakayama, R.; Hasaka, A.; Takahashi, A.; Ryo, K.; Muramatsu, T.; Ide, F.; Mishima, K.; Saito, I. Resveratrol improves salivary dysfunction in a non-obese diabetic (NOD) mouse model of Sjögren’s syndrome. J. Clin. Biochem. Nutr. 2016, 59, 107–112. [Google Scholar] [CrossRef]
- Yan, S.F.; Ramasamy, R.; Schmidt, A.M. Mechanisms of disease: Advanced glycation end-products and their receptor in inflammation and diabetes complications. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Brownlee, M. Hyperglycemia-induced reactive oxygen species increase expression of the receptor for advanced glycation end products (RAGE) and RAGE ligands. Diabetes 2010, 59, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.; Kang, R.; Zeh, H.J., 3rd; Lotze, M.T. High-mobility group box 1, oxidative stress, and disease. Antioxid. Redox. Signal. 2011, 14, 1315–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2’ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Xiang, R.-L.; Huang, Y.; Zhang, Y.; Cong, X.; Zhang, Z.-J.; Wu, L.-L.; Yu, G.-Y. Type 2 diabetes-induced hyposalivation of the submandibular gland through PINK1/Parkin-mediated mitophagy. J. Cell. Physiol. 2020, 235, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Allushi, B.; Bagavant, H.; Papinska, J.; Deshmukh, U.S. Hyperglycemia and salivary gland dysfunction in the non-obese diabetic mouse: Caveats for preclinical studies in Sjögren’s syndrome. Sci. Rep. 2019, 9, 17969. [Google Scholar] [CrossRef]
- Fedirko, N.V.; Kruglikov, I.A.; Kopach, O.V.; Vats, J.A.; Kostyuk, P.G.; Voitenko, N.V. Changes in functioning of rat submandibular salivary gland under streptozotocin-induced diabetes are associated with alterations of Ca2+ signaling and Ca2+ transporting pumps. Biochim. Biophys. Acta 2006, 1762, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Huang, L.; Li, N.; Miao, J.; Liu, W.; Yu, J. Ameliorative effect of polydatin on hyperglycemia and renal injury in streptozotocin-induced diabetic rats. Cell Mol. Biol. 2019, 65, 55–59. [Google Scholar] [CrossRef]
- Yousef, A.; Shawki, H.; El-Shahawy, A.; El-Twab, S.; Abdel Moneim, A.; Oishi, H. Polydatin mitigates pancreatic β-cell damage through its antioxidant activity. Biomed. Pharmacother. 2021, 133, 111027. [Google Scholar] [CrossRef]
- Gong, W.; Li, J.; Chen, Z.; Huang, J.; Chen, Q.; Cai, W.; Liu, P.; Huang, H. Polydatin promotes Nrf2-ARE anti-oxidative pathway through activating CKIP-1 to resist HG-induced up-regulation of FN and ICAM-1 in GMCs and diabetic mice kidneys. Free Radic. Biol. Med. 2017, 106, 393–405. [Google Scholar] [CrossRef]
- Chen, C.; Huang, K.; Hao, J.; Huang, J.; Yang, Z.; Xiong, F.; Liu, P.; Huang, H. Polydatin attenuates AGEs-induced upregulation of fibronectin and ICAM-1 in rat glomerular mesangial cells and db/db diabetic mice kidneys by inhibiting the activation of the SphK1-S1P signaling pathway. Mol. Cell Endocrinol. 2016, 427, 45–56. [Google Scholar] [CrossRef]
- Zheng, L.; Wu, J.; Mo, J.; Guo, L.; Wu, X.; Bao, Y. Polydatin inhibits adipose tissue inflammation and ameliorates lipid metabolism in high-fat-fed mice. Biomed Res. Int. 2019, 2019, 7196535. [Google Scholar] [CrossRef]
- Tramunt, B.; Smati, S.; Grandgeorge, N.; Lenfant, F.; Arnal, J.F.; Montagner, A.; Gourdy, P. Sex differences in metabolic regulation and diabetes susceptibility. Diabetologia 2020, 63, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Elzinga, S.E.; Savelieff, M.G.; O’Brien, P.D.; Mendelson, F.E.; Hayes, J.M.; Feldman, E.L. Sex differences in insulin resistance, but not peripheral neuropathy, in a diet-induced prediabetes mouse model. Dis. Model Mech. 2021, 14, dmm048909. [Google Scholar] [CrossRef] [PubMed]
- Salsali, A.; Nathan, M. A review of types 1 and 2 diabetes mellitus and their treatment with insulin. Am. J. Ther. 2006, 13, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.S.; Jung, W.K.; Park, S.-B.; Kim, H.R.; Kim, J. Gemigliptin suppresses salivary dysfunction in streptozotocin-induced diabetic rats. Biomed. Pharmacother. 2021, 137, 111297. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-L.; Edelstein, D.; Rossetti, L.; Fantus, I.G.; Goldberg, H.; Ziyadeh, F.; Wu, J.; Brownlee, M. Hyperglycemia-induced mitochondrial superoxide overproduction activates the hexosamine pathway and induces plasminogen activator inhibitor-1 expression by increasing Sp1 glycosylation. Proc. Natl. Acad. Sci. USA 2000, 97, 12222–12226. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.J. Redox imbalance stress in diabetes mellitus: Role of the polyol pathway. Anim. Model Exp. Med. 2018, 1, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Auten, R.L.; Davis, J.M. Oxygen toxicity and reactive oxygen species: The devil is in the details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Kranstuber, A.L.; Del Rio, C.; Biesiadecki, B.J.; Hamlin, R.L.; Ottobre, J.; Gyorke, S.; Lacombe, V.A. Advanced glycation end product cross-link breaker attenuates diabetes-induced cardiac dysfunction by improving sarcoplasmic reticulum calcium handling. Front. Physiol. 2012, 3, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanajou, D.; Ghorbani Haghjo, A.; Argani, H.; Aslani, S. AGE-RAGE axis blockade in diabetic nephropathy: Current status and future directions. Eur. J. Pharmacol. 2018, 833, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Negre-Salvayre, A.; Salvayre, R.; Augé, N.; Pamplona, R.; Portero-Otín, M. Hyperglycemia and glycation in diabetic complications. Antioxid. Redox. Signal. 2009, 11, 3071–3109. [Google Scholar] [CrossRef]
- Matsumoto, N.; Omagari, D.; Ushikoshi-Nakayama, R.; Yamazaki, T.; Inoue, H.; Saito, I. Hyperglycemia induces generation of reactive oxygen species and accelerates apoptotic cell death in salivary gland cells. Pathobiology 2021, 88, 234–241. [Google Scholar] [CrossRef]
- Katz, J.; Stavropoulos, F.; Bhattacharyya, I.; Stewart, C.; Perez, F.M.; Caudle, R.M. Receptor of advanced glycation end product (RAGE) expression in the minor salivary glands of patients with Sjögren’s syndrome: A preliminary study. Scand. J. Rheumatol. 2004, 33, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Maleki, V.; Foroumandi, E.; Hajizadeh-Sharafabad, F.; Kheirouri, S.; Alizadeh, M. The effect of resveratrol on advanced glycation end products in diabetes mellitus: A systematic review. Arch. Physiol. Biochem. 2020. [Google Scholar] [CrossRef]
- Sheng, Z.; Ai, B.; Zheng, L.; Zheng, X.; Xu, Z.; Shen, Y.; Jin, Z. Inhibitory activities of kaempferol, galangin, carnosic acid and polydatin against glycation and α-amylase and α-glucosidase enzymes. Int. J. Food Sci. Technol. 2018, 53, 755–766. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Z.; Xu, Z.; Feng, W.; Yang, X.; Qi, Z. Polydatin protects Schwann cells from methylglyoxal induced cytotoxicity and promotes crushed sciatic nerves regeneration of diabetic rats. Phytother. Res. 2021, 35, 4592–4604. [Google Scholar] [CrossRef]
- Yu, Y.; Tang, D.; Kang, R. Oxidative stress-mediated HMGB1 biology. Front. Physiol. 2015, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.; Liu, Y.; Deterding, L.J.; Poltoratsky, V.P.; Kedar, P.S.; Horton, J.K.; Kanno, S.; Asagoshi, K.; Hou, E.W.; Khodyreva, S.N.; et al. HMGB1 is a cofactor in mammalian base excision repair. Mol. Cell 2007, 27, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.S.; Jo, K.; Kim, J.S.; Pyo, M.K.; Kim, J. GS-E3D, a new pectin lyase-modified red ginseng extract, inhibited diabetes-related renal dysfunction in streptozotocin-induced diabetic rats. BMC Complement. Altern. Med. 2017, 17, 430. [Google Scholar] [CrossRef]
- Fukuoka, C.Y.; Simões, A.; Uchiyama, T.; Arana-Chavez, V.E.; Abiko, Y.; Kuboyama, N.; Bhawal, U.K. The effects of low-power laser irradiation on inflammation and apoptosis in submandibular glands of diabetes-induced rats. PLoS ONE 2017, 12, e0169443. [Google Scholar] [CrossRef]
- Ryo, K.; Yamada, H.; Nakagawa, Y.; Tai, Y.; Obara, K.; Inoue, H.; Mishima, K.; Saito, I. Possible involvement of oxidative stress in salivary gland of patients with Sjogren’s syndrome. Pathobiology 2006, 73, 252–260. [Google Scholar] [CrossRef]
- Martín, A.R.; Villegas, I.; La Casa, C.; de la Lastra, C.A. Resveratrol, a polyphenol found in grapes, suppresses oxidative damage and stimulates apoptosis during early colonic inflammation in rats. Biochem. Pharmacol. 2004, 67, 1399–1410. [Google Scholar]
- Wang, H.L.; Gao, J.P.; Han, Y.L.; Xu, X.; Wu, R.; Gao, Y.; Cui, X.H. Comparative studies of polydatin and resveratrol on mutual transformation and antioxidative effect in vivo. Phytomedicine 2015, 22, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Regev-Shoshani, G.; Shoseyov, O.; Bilkis, I.; Kerem, Z. Glycosylation of resveratrol protects it from enzymic oxidation. Biochem. J. 2003, 374, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Perrella, F.; Coppola, F.; Petrone, A.; Platella, C.; Montesarchio, D.; Stringaro, A.; Ravagnan, G.; Fuggetta, M.P.; Rega, N.; Musumeci, D. Interference of polydatin/resveratrol in the ACE2: Spike recognition during COVID-19 infection. A focus on their potential mechanism of action through computational and biochemical assays. Biomolecules 2021, 11, 1048. [Google Scholar] [CrossRef]
- Huang, Y.; Mao, Q.Y.; Shi, X.J.; Cong, X.; Zhang, Y.; Wu, L.L.; Yu, G.Y.; Xiang, R.L. Disruption of tight junctions contributes to hyposalivation of salivary glands in a mouse model of type 2 diabetes. J. Anat. 2020, 237, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Castro, I.; Barrera, M.-J.; González, S.; Aguilera, S.; Urzúa, U.; Cortés, J.; González, M.-J. Mucins in salivary gland development, regeneration, and disease. In Salivary Gland Development and Regeneration: Advances in Research and Clinical Approaches to Functional Restoration; Springer: New York, NY, USA, 2017; pp. 45–71. [Google Scholar]
- Maciejczyk, M.; Skutnik-Radziszewska, A.; Zieniewska, I.; Matczuk, J.; Domel, E.; Waszkiel, D.; Żendzian-Piotrowska, M.; Szarmach, I.; Zalewska, A. Antioxidant defense, oxidative modification, and salivary gland function in an early phase of cerulein pancreatitis. Oxid. Med. Cell. Longev. 2019, 2019, 8403578. [Google Scholar] [CrossRef]
- Ambudkar, I.S. Ca2+ signaling and regulation of fluid secretion in salivary gland acinar cells. Cell Calcium 2014, 55, 297–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agre, P.; King, L.S.; Yasui, M.; Guggino, W.B.; Ottersen, O.P.; Fujiyoshi, Y.; Engel, A.; Nielsen, S. Aquaporin water channels--from atomic structure to clinical medicine. J. Physiol. 2002, 542, 3–16. [Google Scholar] [CrossRef]
- Satoh, K.; Narita, T.; Matsuki-Fukushima, M.; Okabayashi, K.; Ito, T.; Senpuku, H.; Sugiya, H. E2f1-deficient NOD/SCID mice have dry mouth due to a change of acinar/duct structure and the down-regulation of AQP5 in the salivary gland. Pflugers Arch. 2013, 465, 271–281. [Google Scholar] [CrossRef]
- Ma, T.; Song, Y.; Gillespie, A.; Carlson, E.J.; Epstein, C.J.; Verkman, A.S. Defective secretion of saliva in transgenic mice lacking aquaporin-5 water channels. J. Biol. Chem. 1999, 274, 20071–20074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Mori, S.; Date, F.; Hong, G. Epigallocatechin gallate stimulates the neuroreactive salivary secretomotor system in autoimmune sialadenitis of MRL-Fas(lpr) mice via activation of cAMP-dependent protein kinase A and inactivation of nuclear factor κB. Autoimmunity 2015, 48, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, A.; Posa, F.; De Maria, S.; Ravagnan, G.; Ballini, A.; Porro, C.; Trotta, T.; Grano, M.; Muzio, L.L.; Mori, G. Polydatin, natural precursor of resveratrol, promotes osteogenic differentiation of mesenchymal stem cells. Int. J. Med. Sci. 2018, 15, 944–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Tindall, D.J. Dynamic FoxO transcription factors. J. Cell Sci. 2007, 120, 2479–2487. [Google Scholar] [CrossRef] [Green Version]
- Motta, M.C.; Divecha, N.; Lemieux, M.; Kamel, C.; Chen, D.; Gu, W.; Bultsma, Y.; McBurney, M.; Guarente, L. Mammalian SIRT1 represses forkhead transcription factors. Cell 2004, 116, 551–563. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Lee, S.W.; Kang, M.; Choi, J.K.; Park, K.; Byun, J.S.; Kim, D.Y. FoxO1 as a Regulator of Aquaporin 5 Expression in the Salivary Gland. J. Dent. Res. 2021, 100, 1281–1288. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Suzuki, T.; Koyama, H.; Tanaka, S.; Takata, K. Aquaporin-5 (AQP5), a water channel protein, in the rat salivary and lacrimal glands: Immunolocalization and effect of secretory stimulation. Cell Tissue Res. 1999, 295, 513–521. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal | DM | DM + PD | |
|---|---|---|---|
| Body weight (g) | 26.46 ± 0.37 | 30.74 ± 1.30 * | 30.06 ± 1.86 * |
| Blood glucose (mg/dL) | 179.29 ± 17.61 | 571.33 ± 70.22 * | 562.86 ± 98.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.R.; Jung, W.K.; Park, S.-B.; Ryu, H.Y.; Kim, Y.H.; Kim, J. Polydatin Alleviates Diabetes-Induced Hyposalivation through Anti-Glycation Activity in db/db Mouse. Pharmaceutics 2022, 14, 51. https://doi.org/10.3390/pharmaceutics14010051
Kim HR, Jung WK, Park S-B, Ryu HY, Kim YH, Kim J. Polydatin Alleviates Diabetes-Induced Hyposalivation through Anti-Glycation Activity in db/db Mouse. Pharmaceutics. 2022; 14(1):51. https://doi.org/10.3390/pharmaceutics14010051
Chicago/Turabian StyleKim, Hyung Rae, Woo Kwon Jung, Su-Bin Park, Hwa Young Ryu, Yong Hwan Kim, and Junghyun Kim. 2022. "Polydatin Alleviates Diabetes-Induced Hyposalivation through Anti-Glycation Activity in db/db Mouse" Pharmaceutics 14, no. 1: 51. https://doi.org/10.3390/pharmaceutics14010051
APA StyleKim, H. R., Jung, W. K., Park, S. -B., Ryu, H. Y., Kim, Y. H., & Kim, J. (2022). Polydatin Alleviates Diabetes-Induced Hyposalivation through Anti-Glycation Activity in db/db Mouse. Pharmaceutics, 14(1), 51. https://doi.org/10.3390/pharmaceutics14010051