
Lactoferrin Binding to SARS-CoV-2 Spike Glycoprotein Blocks Pseudoviral Entry and Relieves Iron Protein Dysregulation in Several In Vitro Models

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
, 
 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bovine and Human Lactoferrin
2.2. Cell Culture and Pseudovirus
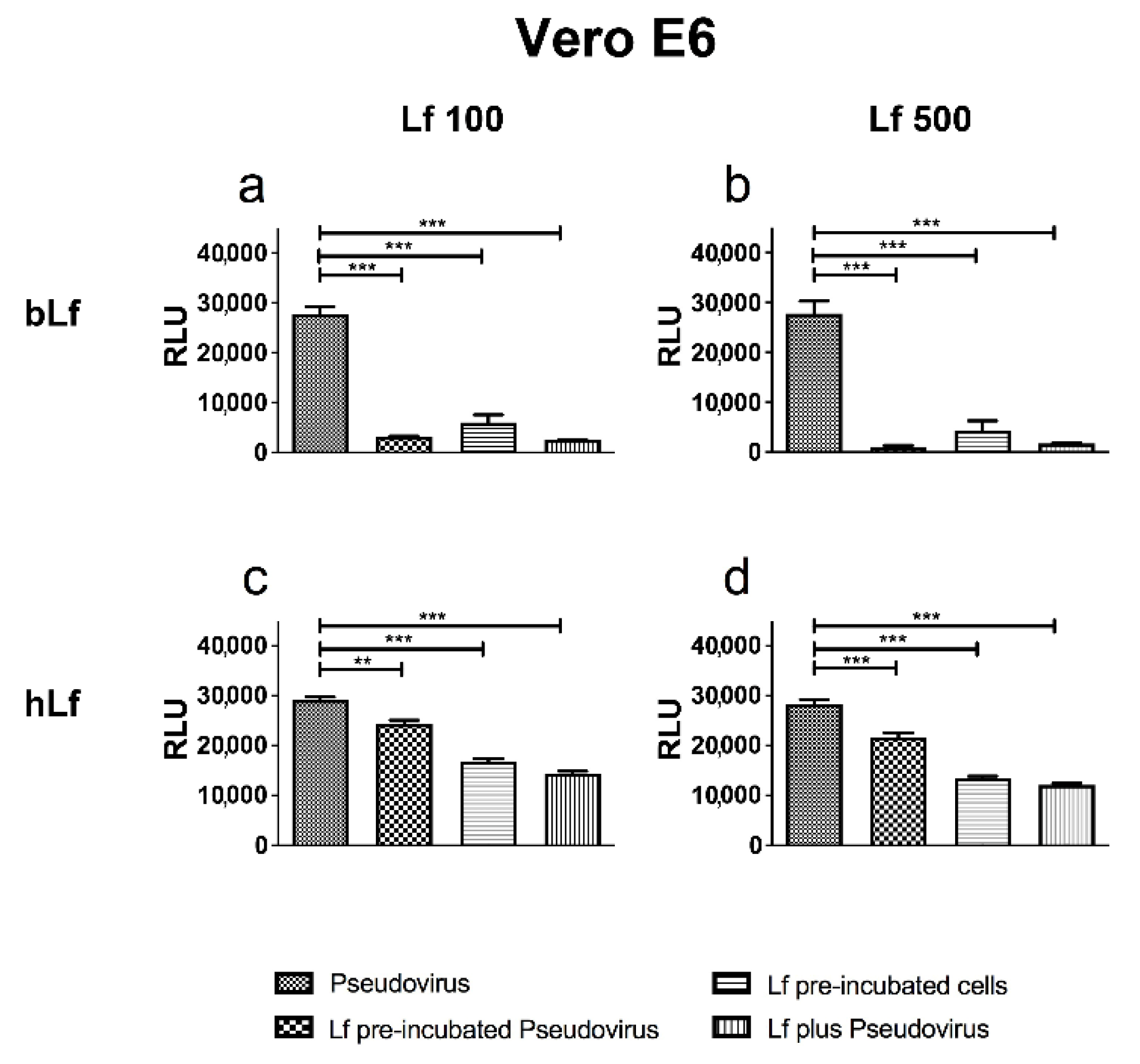
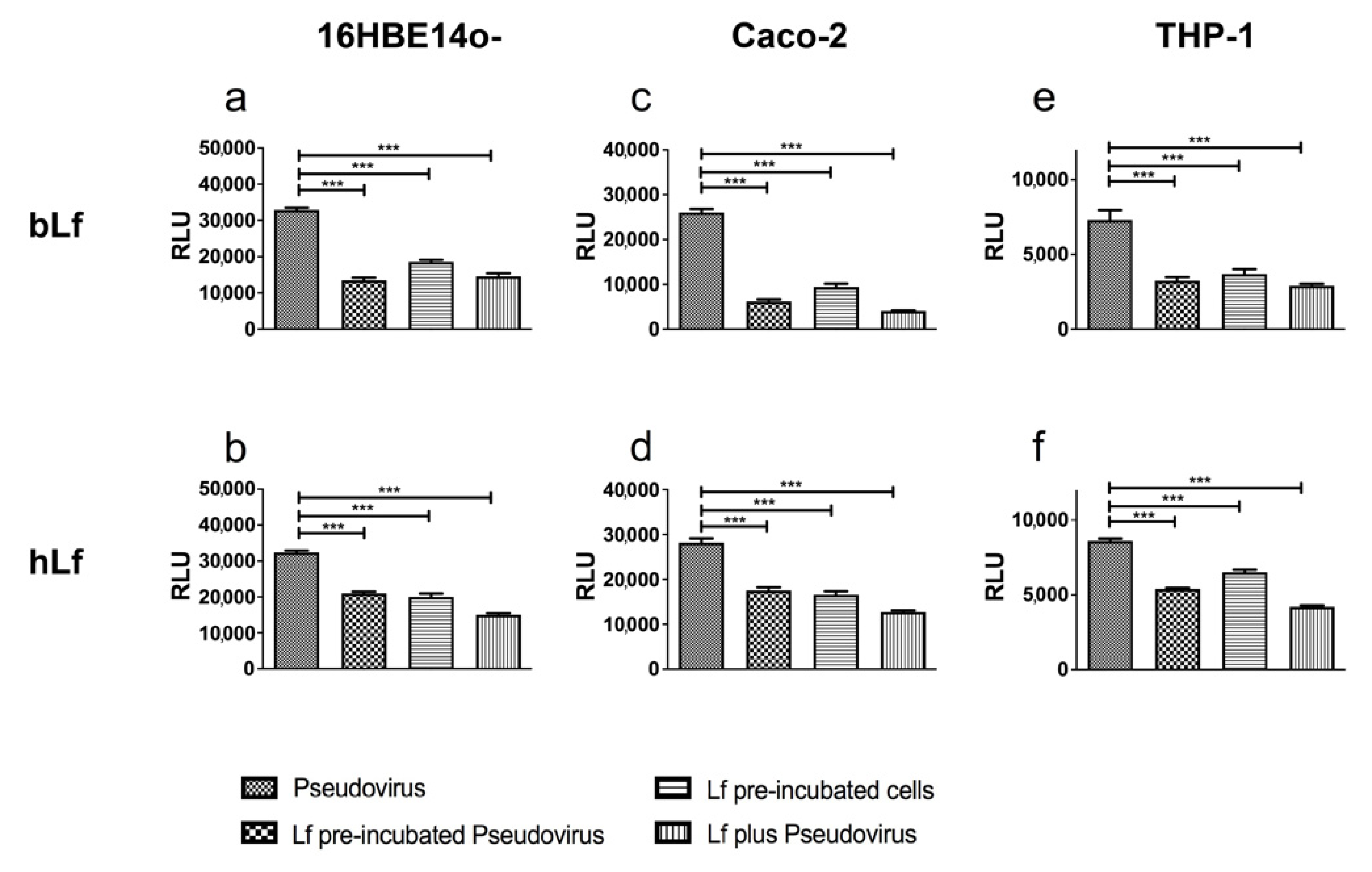
2.3. Pseudovirus Neutralization Assay
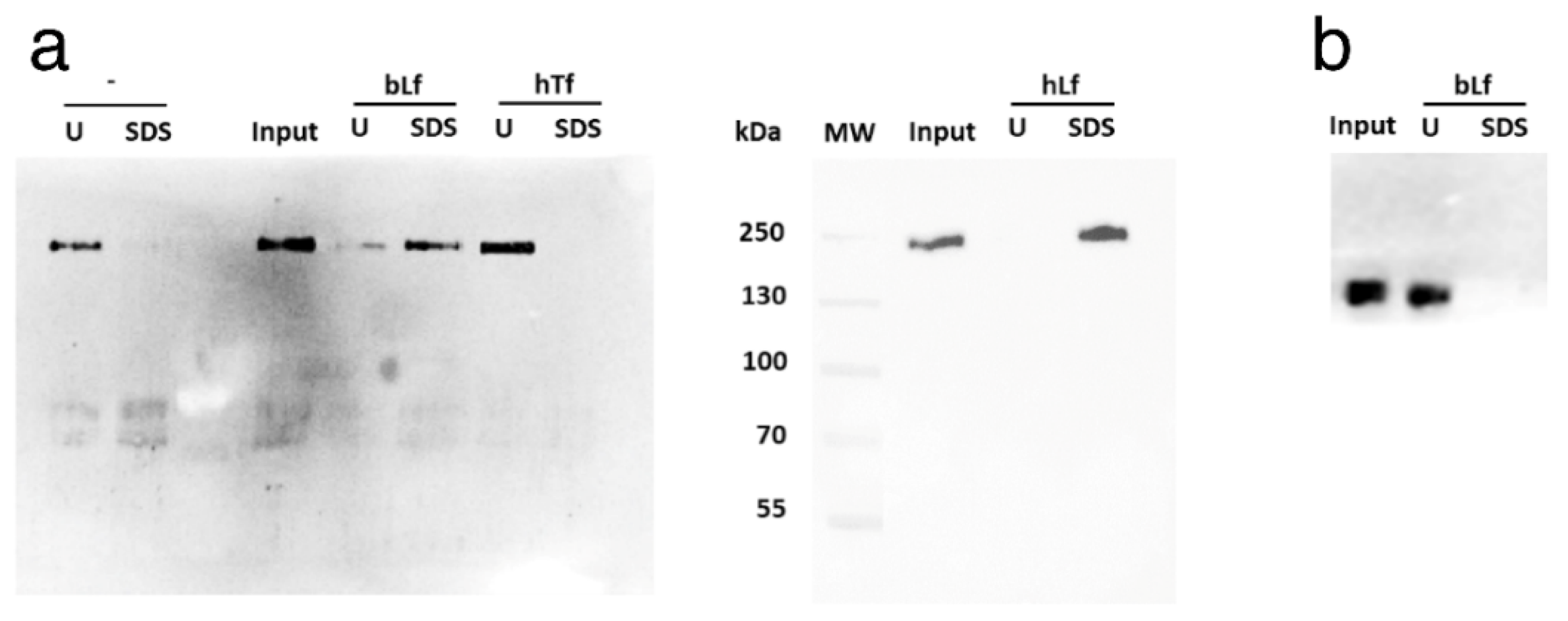
2.4. Sepharose 6B Pull-Down
2.5. Stimulation of Caco-2 and Differentiated THP-1 Cells with Spike
2.6. Cytokine Analysis
2.7. Western Blots
2.8. Structures Preparations and Molecular Docking Simulations of the TfR1-Lfs Complexes
2.9. Modelling of the SARS-CoV-2 Variant Structures
2.10. Protein–Protein Docking Methods
2.11. Molecular Dynamics
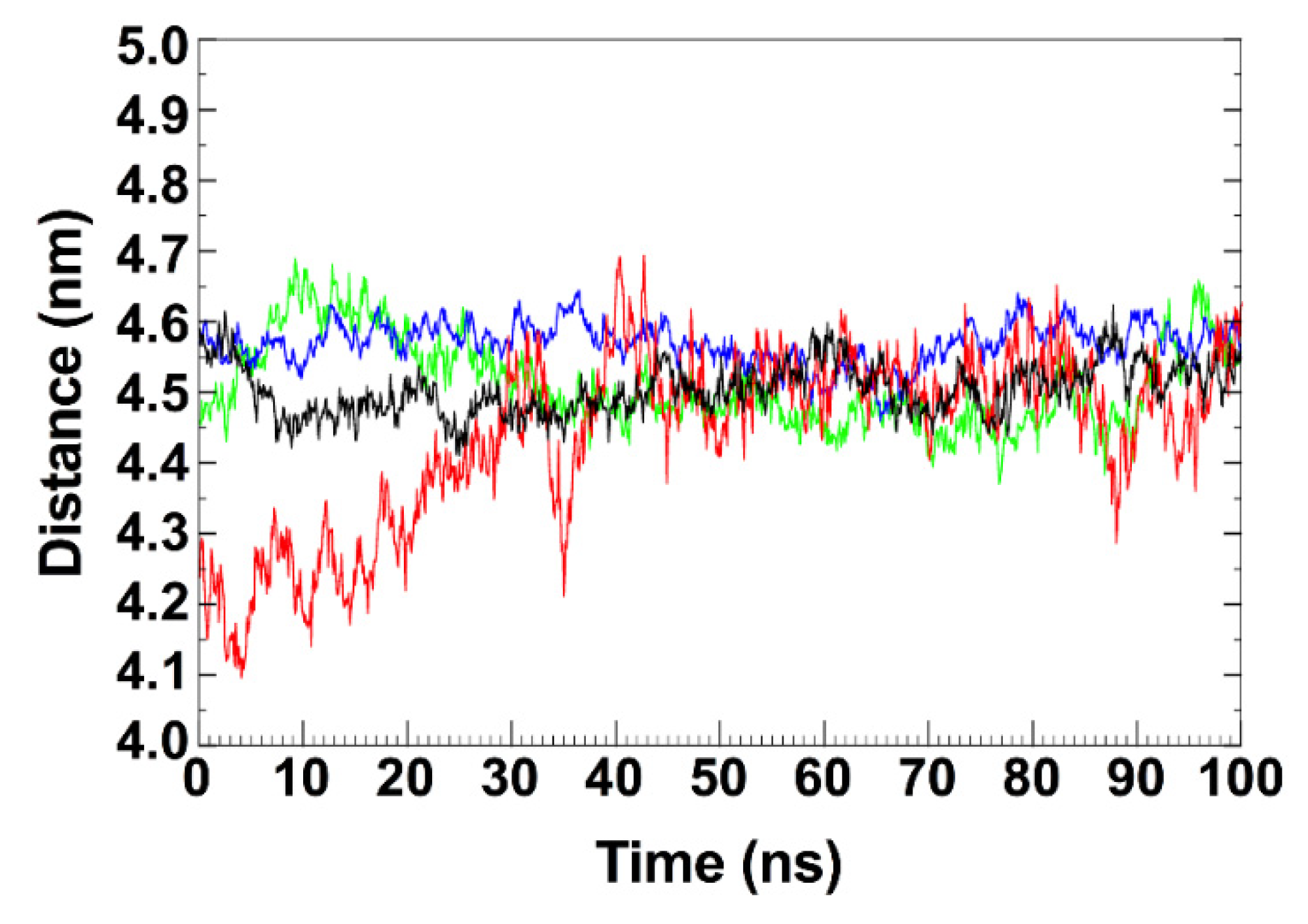
2.12. Trajectory Analysis
2.13. Statistical Analysis
3. Results
3.1. Lactoferrins Exert Neutralizing Activity against SARS-CoV-2 Spike Pseudovirus
3.2. Lactoferrins Bind to SARS-CoV-2 Spike
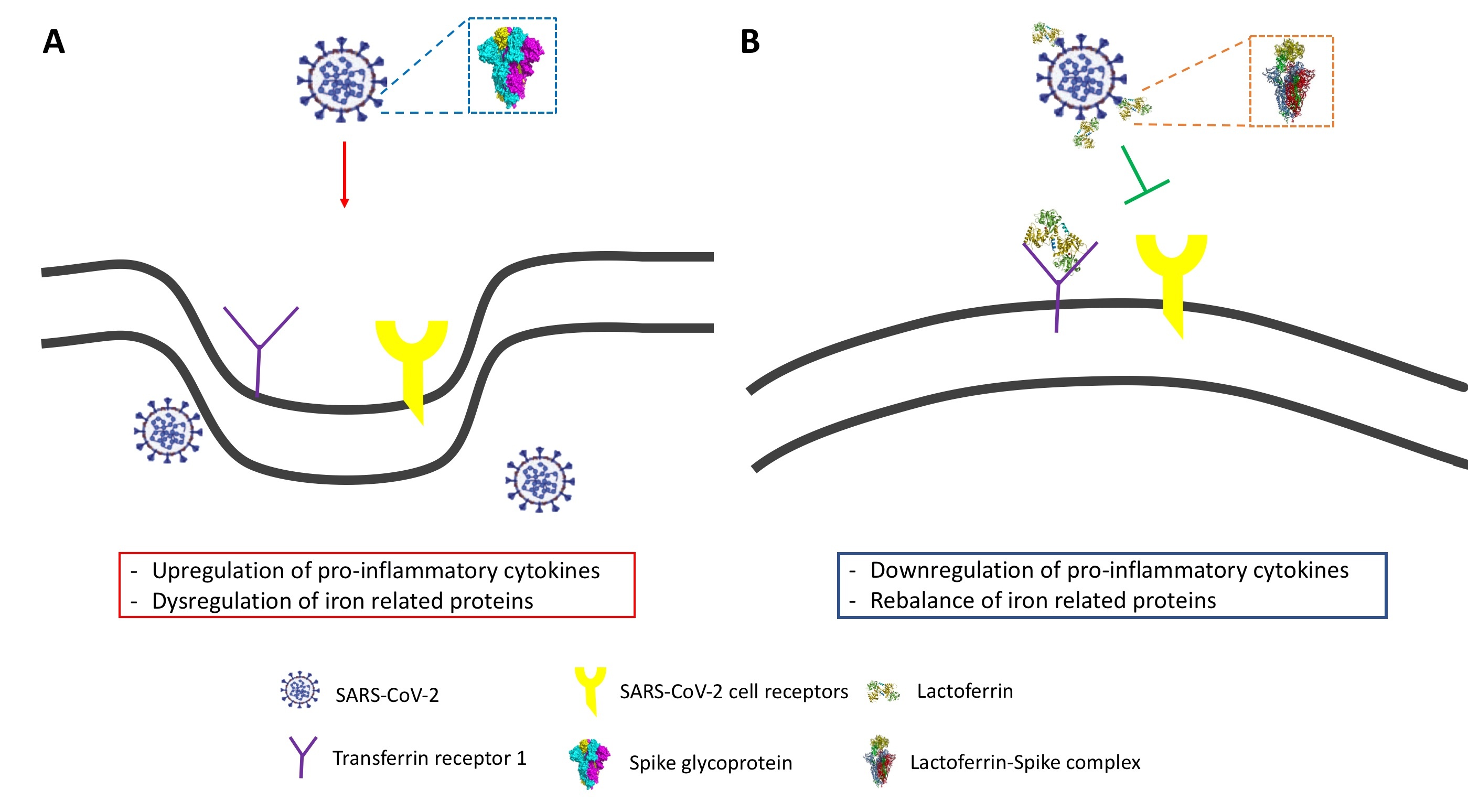
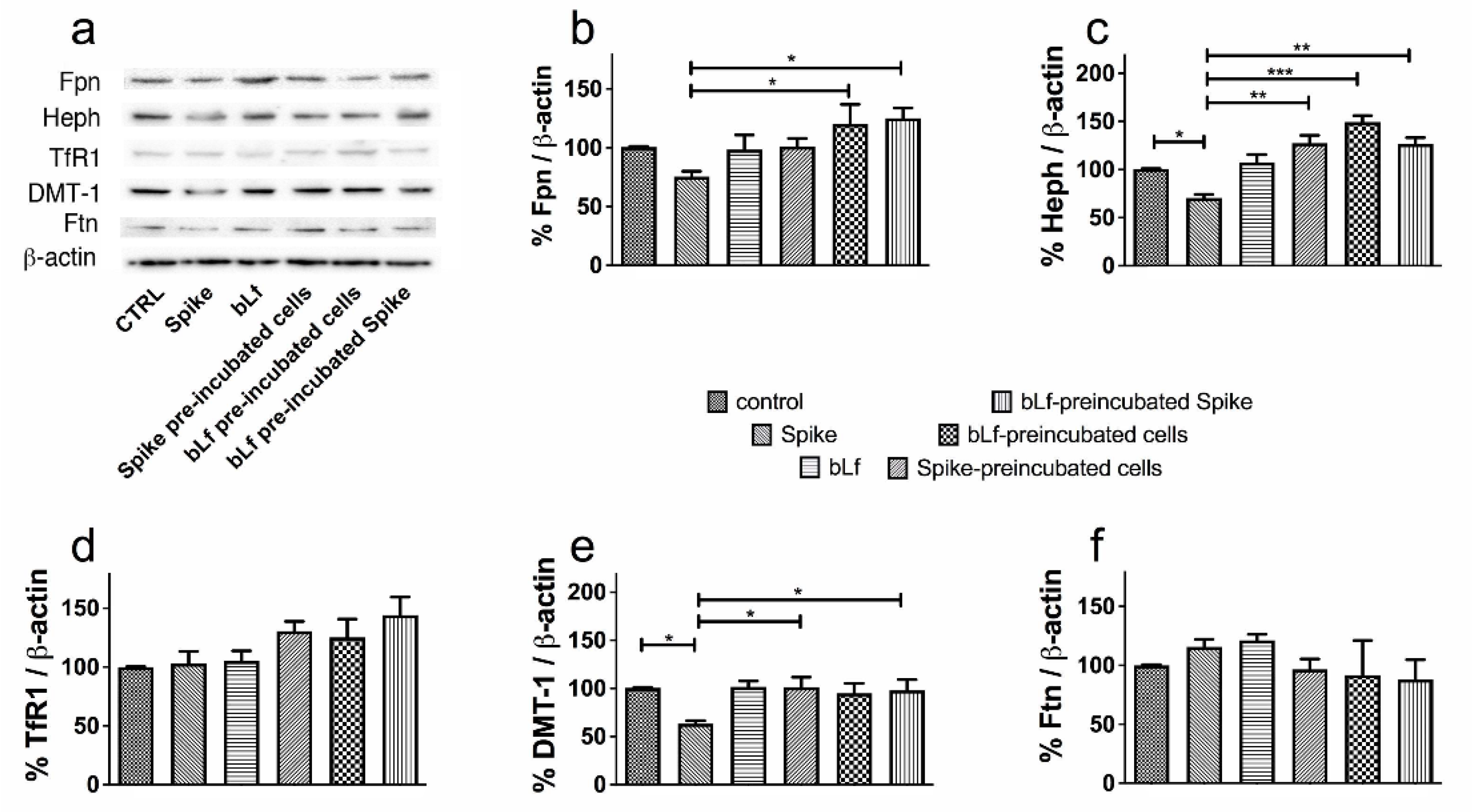
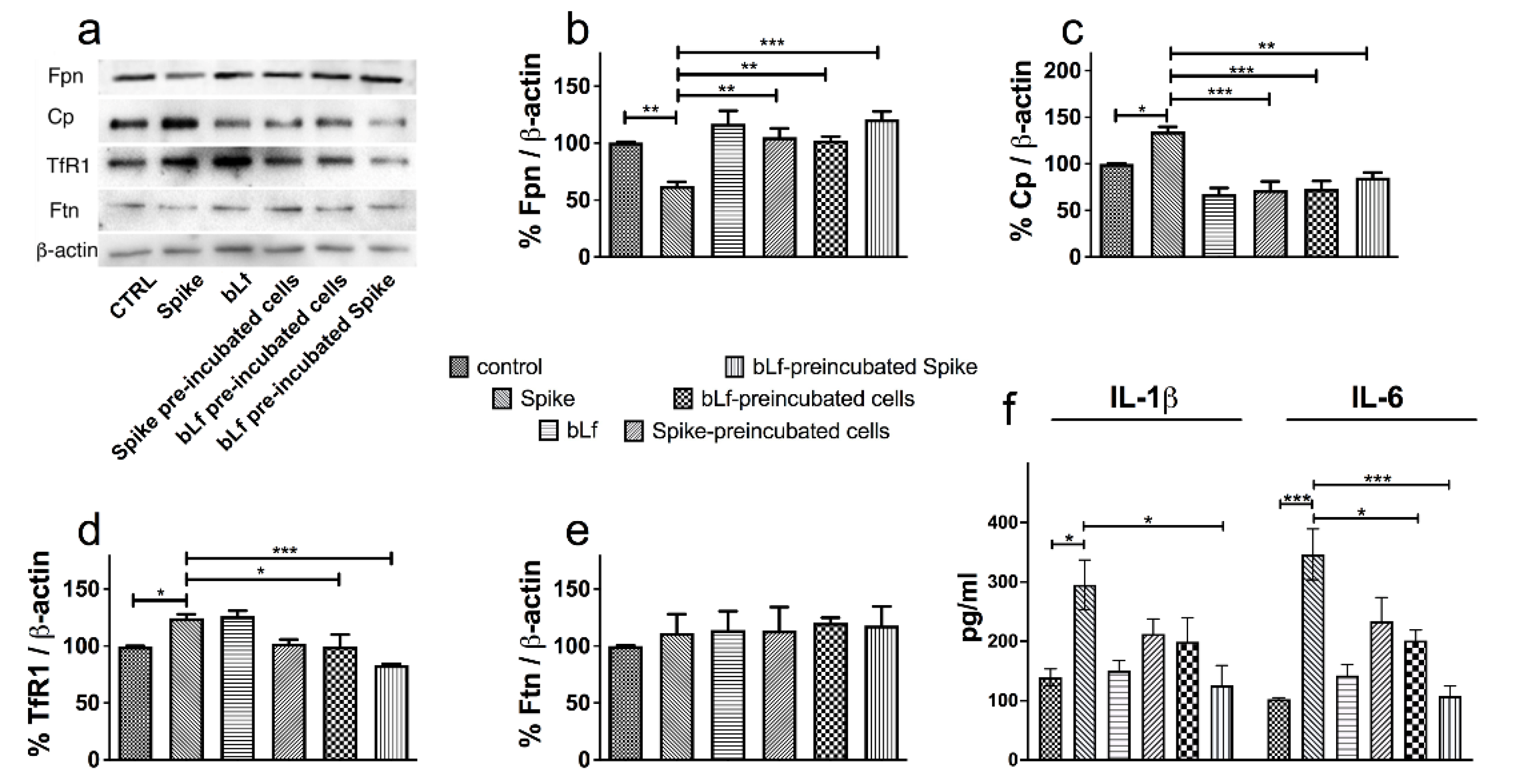
3.3. Bovine Lactoferrin Counteracts the Dysregulation of Iron Proteins Induced by SARS-CoV-2 Spike
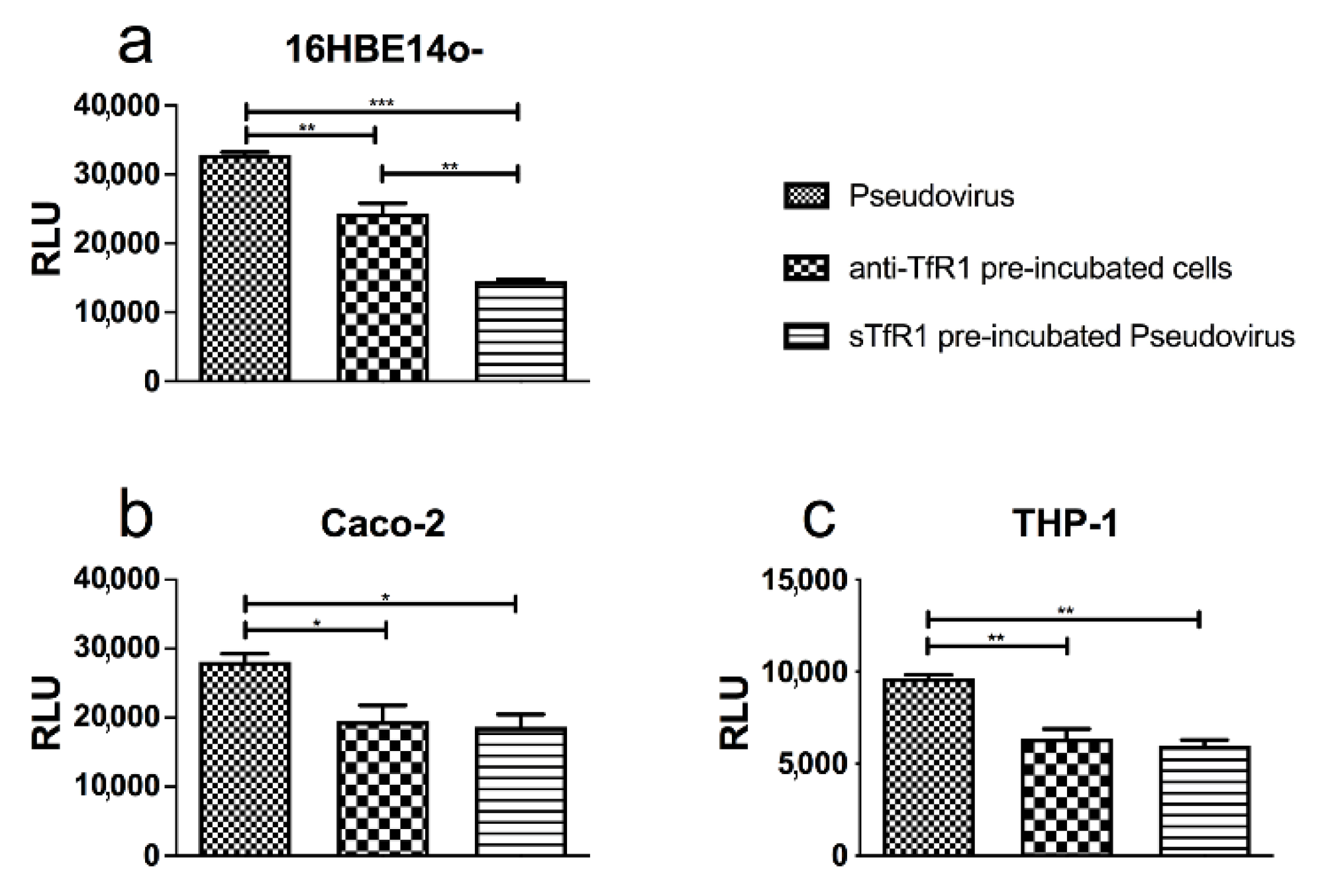
3.4. TfR1 Contributes to SARS-CoV-2 Spike Pseudovirus Fusion to Cell Membrane
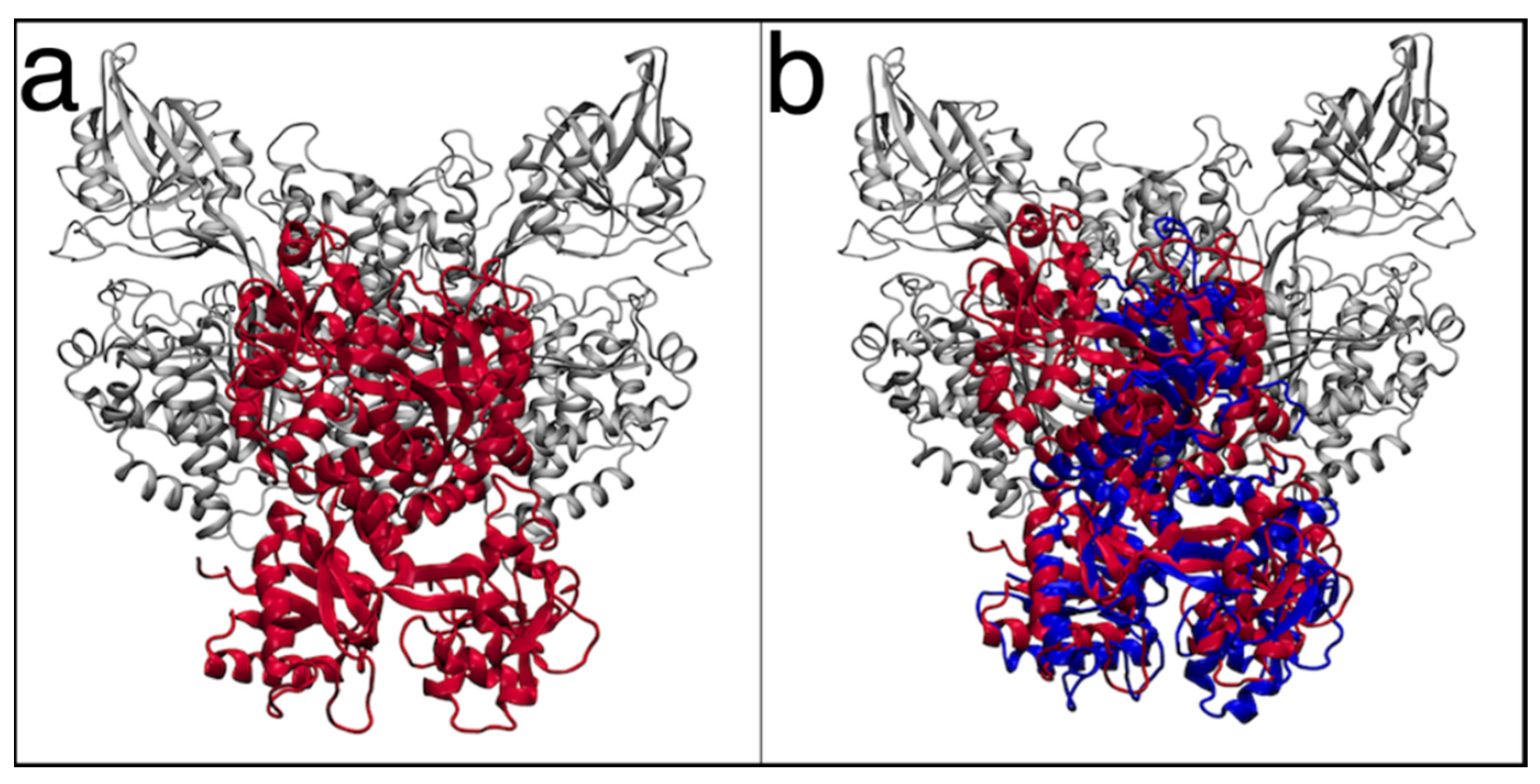
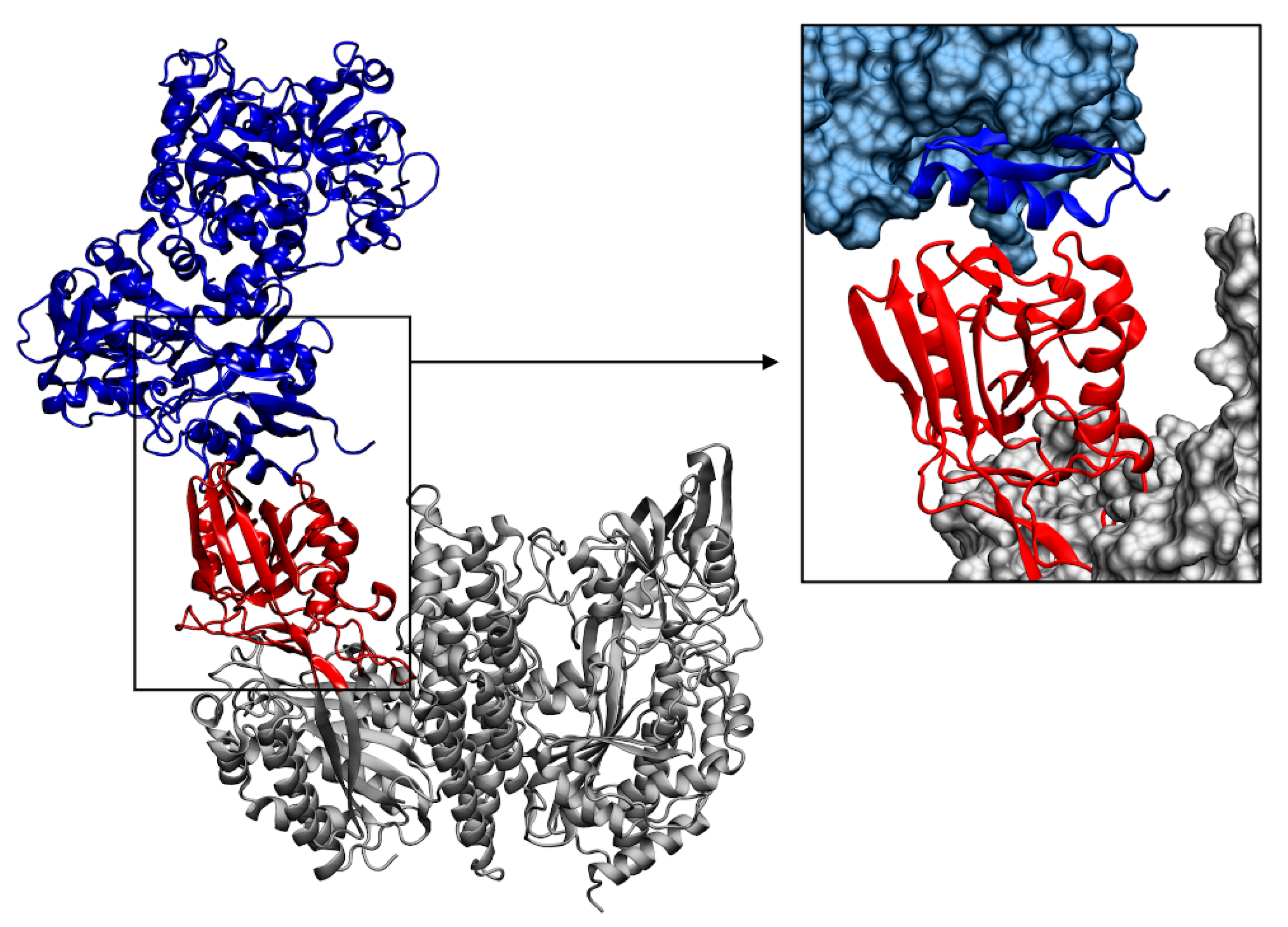
3.5. Molecular Docking Simulations of TfR-1 in Complex with Lactoferrins
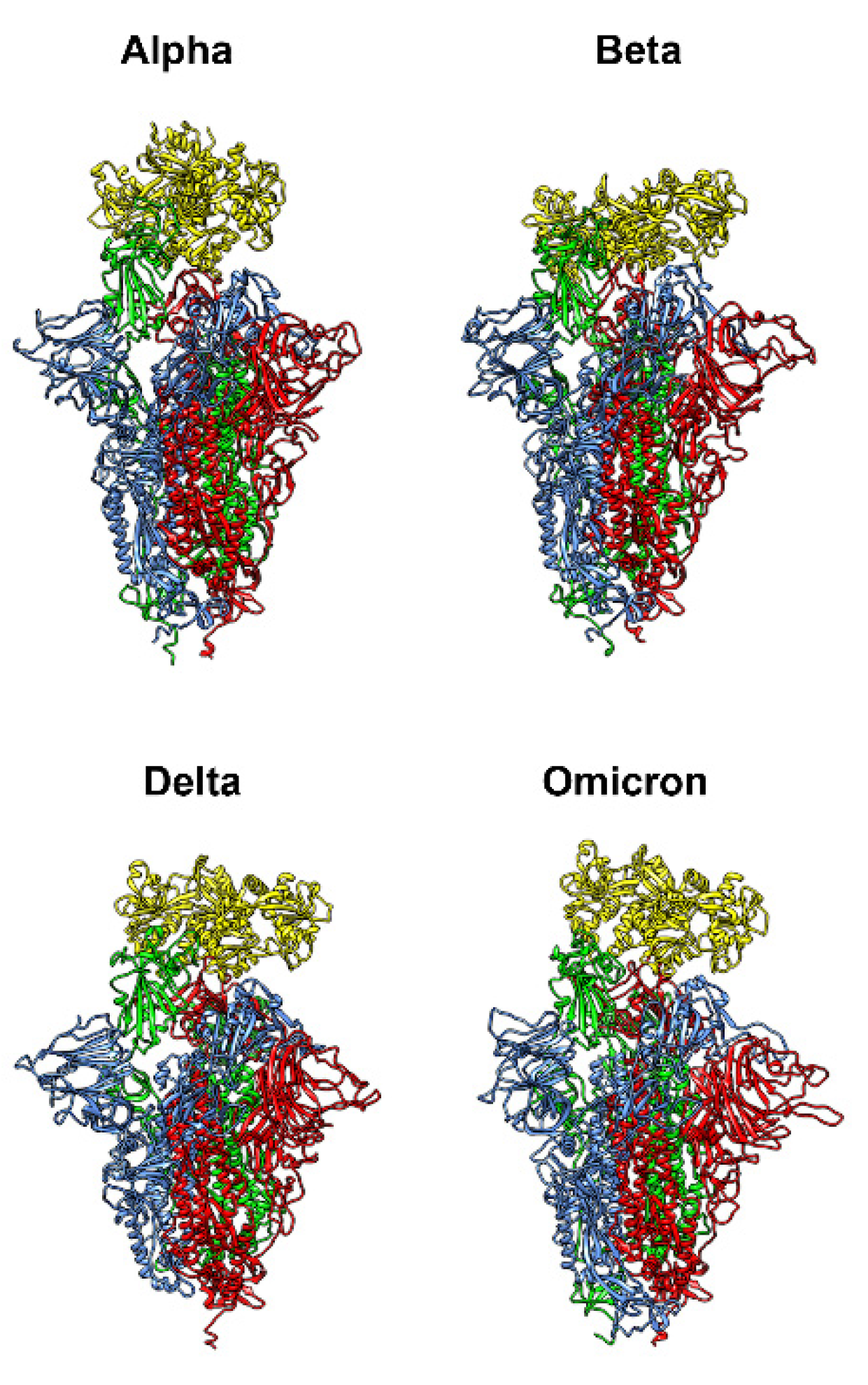
3.6. Computational Results on Bovine Lactoferrin and Spike Variants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic Characterisation and Epidemiology of 2019 Novel Coronavirus: Implications for Virus Origins and Receptor Binding. The Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS- CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, L.; Xu, M.; Wu, J.; Luo, D.; Zhu, Y.; Li, B.; Song, X.; Zhou, X. Prognostic value of interleukin-6, C-reactive protein, and procalcitonin in patients with COVID-19. J. Clin. Virol. 2020, 127, 104370. [Google Scholar] [CrossRef]
- Patra, T.; Meyer, K.; Geerling, L.; Isbell, T.S.; Hoft, D.F.; Brien, J.; Pinto, A.K.; Ray, R.B.; Ray, R. SARS-CoV-2 Spike protein promotes IL-6 trans-signaling by activation of angiotensin II receptor signaling in epithelial cells. PLoS Pathog. 2020, 16, e1009128. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Nemeth, E. Iron homeostasis in host defence and inflammation. Nat. Rev. Immunol. 2015, 15, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazer, D.M.; Anderson, G.J. The orchestration of body iron intake: How and where do enterocytes receive their cues? Blood Cells Mol. Dis. 2003, 30, 288–297. [Google Scholar] [CrossRef]
- Mackenzie, B.; Garrick, M.D. Iron imports: II. Iron uptake at the apical membrane in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G981–G986. [Google Scholar] [CrossRef] [Green Version]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Bonaccorsi di Patti, M.C.; Cutone, A.; Polticelli, F.; Rosa, L.; Lepanto, M.S.; Valenti, P.; Musci, G. The ferroportin-ceruloplasmin system and the mammalian iron homeostasis machine: Regulatory pathways and the role of lactoferrin. Biometals 2018, 31, 399–414. [Google Scholar] [CrossRef]
- Mancinelli, R.; Rosa, L.; Cutone, A.; Lepanto, M.S.; Franchitto, A.; Onori, P.; Gaudio, E.; Valenti, P. Viral Hepatitis and Iron Dysregulation: Molecular Pathways and the Role of Lactoferrin. Molecules 2020, 25, 1997. [Google Scholar] [CrossRef] [PubMed]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence against microbial and viral attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.; Cutone, A.; Lepanto, M.S.; Paesano, R.; Valenti, P. Lactoferrin: A Natural Glycoprotein Involved in Iron and Inflammatory Homeostasis. Int. J. Mol. Sci. 2017, 18, 1985. [Google Scholar] [CrossRef]
- Hu, Y.; Meng, X.; Zhang, F.; Xiang, Y.; Wang, J. The in vitro antiviral activity of lactoferrin against common human coronaviruses and SARS-CoV-2 is mediated by targeting the heparan sulfate co-receptor. Emerg. Microbes Infect. 2021, 10, 317–330. [Google Scholar] [CrossRef]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin against SARS-CoV-2: In vitro and in silico evidences. Front. Pharmacol. 2021, 12, 666600. [Google Scholar] [CrossRef]
- Mirabelli, C.; Wotring, J.W.; Zhang, C.J.; McCarty, S.M.; Fursmidt, R.; Pretto, C.D.; Qiao, Y.; Zhang, Y.; Frum, T.; Kadambi, N.S.; et al. Morphological cell profiling of SARS-CoV-2 infection identifies drug repurposing candidates for COVID-19. Proc. Natl. Acad. Sci. USA 2021, 118, e2105815118. [Google Scholar] [CrossRef]
- Tsuji, Y. Transmembrane protein western blotting: Impact of sample preparation on detection of SLC11A2 (DMT1) and SLC40A1 (ferroportin). PLoS ONE 2020, 15, e0235563. [Google Scholar] [CrossRef]
- Crook, Z.R.; Girard, E.; Sevilla, G.P.; Merrill, M.; Friend, D.; Rupert, P.B.; Pakiam, F.; Nguyen, E.; Yin, C.; Ruff, R.O.; et al. A TfR-Binding Cystine-Dense Peptide Promotes Blood–Brain Barrier Penetration of Bioactive Molecules. J. Mol. Biol. 2020, 432, 3989–4009. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y.; et al. Potent Binding of 2019 Novel Coronavirus Spike Protein by a SARS Coronavirus-specific Human Monoclonal Antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, D.; Betz, R.; Cerutti, D.; Cheatham, T.; Darden, T.; Duke, R. Amber 2016; University of California: San Francisco, CA, USA, 2016. [Google Scholar]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein-protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef]
- McGibbon, R.T.; Beauchamp, K.A.; Harrigan, M.P.; Klein, C.; Swails, J.M.; Hernández, C.X.; Schwantes, C.R.; Wang, L.P.; Lane, T.J.; Pande, V.S. MDTraj: A Modern Open Library for the Analysis of Molecular Dynamics Trajectories. Biophys. J. 2015, 109, 1528–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckenroth, B.E.; Steere, A.N.; Chasteen, N.D.; Everse, S.J.; Mason, A.B. How the binding of human transferrin primes the transferrin receptor potentiating iron release at endosomal pH. Proc. Natl. Acad. Sci. USA 2011, 108, 13089–13094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romeo, A.; Iacovelli, F.; Falconi, M. Targeting the SARS-CoV-2 Spike Glycoprotein Prefusion Conformation: Virtual Screening and Molecular Dynamics Simulations Applied to the Identification of Potential Fusion Inhibitors. Virus Res. 2020, 286, 198068. [Google Scholar] [CrossRef]
- Moore, S.A.; Anderson, B.F.; Groom, C.R.; Haridas, M.; Baker, E.N. Three-dimensional Structure of Diferric Bovine Lactoferrin at 2.8 Å Resolution. J. Mol. Biol. 1997, 274, 222–236. [Google Scholar] [CrossRef]
- Ramírez-Aportela, E.; López-Blanco, J.R.; Chacón, P. FRODOCK 2.0: Fast Protein-Protein Docking Server. Bioinformatics 2016, 32, 2386–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garzon, J.I.; Lopéz-Blanco, J.R.; Pons, C.; Kovacs, J.; Abagyan, R.; Fernandez-Recio, J.; Chacon, P. FRODOCK: A New Approach for Fast Rotational Protein-Protein Docking. Bioinformatics 2009, 25, 2544–2551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An Overview of the Amber Biomolecular Simulation Package. WIREs Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J.C. Numerical Integration of the Cartesian Equations of Motion of a System with Constraints: Molecular Dynamics of N-Alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable Molecular Dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Iannone, F.; Ambrosino, F.; Bracco, G.; De Rosa, M.; Funel, A.; Guarnieri, G.; Migliori, S.; Palombi, F.; Ponti, G.; Santomauro, G.; et al. CRESCO ENEA HPC Clusters: A Working Example of a Multifabric GPFS Spectrum Scale Layout. In Proceedings of the International Conference on High Performance Computing & Simulation (HPCS), Dublin, Ireland, 15–19 July 2019; pp. 1051–1052. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elass, E.; Masson, M.; Mazurier, J.; Legrand, D. Lactoferrin inhibits the lipopolysaccharide-induced expression and proteoglycan-binding ability of interleukin-8 in human endothelial cells. Infect. Immun. 2002, 70, 1860–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.A.; Wilk, K.M.; Bangale, Y.A.; Kruzel, M.L.; Actor, J.K. Lactoferrin modulation of IL-12 and IL-10 response from activated murine leukocytes. Med. Microbiol. Immunol. 2007, 196, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inubushi, T.; Kawazoe, A.; Miyauchi, M.; Kudo, Y.; Ao, M.; Ishikado, A.; Makino, T.; Takata, T. Molecular mechanisms of the inhibitory effects of bovine lactoferrin on lipopolysaccharide-mediated osteoclastogenesis. J. Biol. Chem. 2012, 287, 23527–23536. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, K.; Motsuchi, W.; Tanaka, S.; Dosako, S. Inhibition with lactoferrin of in vitro infection with human herpes virus. Jpn. J. Med. Sci. Biol. 1994, 47, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Oda, H.; Kolawole, A.O.; Mirabelli, C.; Wakabayashi, H.; Tanaka, M.; Yamauchi, K.; Abe, F.; Wobus, C.E. Antiviral effects of bovine lactoferrin on human norovirus. Biochem. Cell Biol. 2021, 99, 166–172. [Google Scholar] [CrossRef]
- Wotring, J.W.; Fursmidt, R.; Ward, L.; Sexton, J.Z. Evaluating the in vitro efficacy of bovine lactoferrin products against SARS-CoV-2 variants of concern. J. Dairy Sci. 2022, 105, 2791–2802. [Google Scholar] [CrossRef]
- Bennett, R.M.; Mohla, C. A solid-phase radioimmunoassay for the measurement of lactoferrin in human plasma: Variations with age, sex and disease. J. Lab. Clin. Med. 1976, 88, 156–166. [Google Scholar]
- Dosch, S.F.; Mahajan, S.D.; Collins, A.R. SARS coronavirus spike protein-induced innate immune response occurs via activation of the NF-kappaB pathway in human monocyte macrophages in vitro. Virus Res. 2009, 142, 19–27. [Google Scholar] [CrossRef]
- Tang, X.; Yang, M.; Duan, Z.; Liao, Z.; Liu, L.; Cheng, R.; Fang, M.; Wang, G.; Liu, H.; Xu, J.; et al. Transferrin receptor is another receptor for SARS-CoV-2 entry. bioRxiv 2020. [Google Scholar] [CrossRef]
- Abraham, J.; Corbett, K.D.; Farzan, M.; Choe, H.; Harrison, S.C. Structural basis for receptor recognition by New World hemorrhagic fever arenaviruses. Nat. Struct. Mol. Biol. 2010, 17, 438–444. [Google Scholar] [CrossRef]
- Saadi, F.; Pal, D.; Sarma, J.D. Spike Glycoprotein Is Central to Coronavirus Pathogenesis-Parallel Between m-CoV and SARS-CoV-2. Ann. Neurosci. 2021, 28, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Tacchini, L.; Gammella, E.; De Ponti, C.; Recalcati, S.; Cairo, G. Role of HIF-1 and NF-kappaB transcription factors in the modulation of transferrin receptor by inflammatory and anti-inflammatory signals. J. Biol. Chem. 2008, 283, 20674–20686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, S.M. The role of iron in viral infections. Front. Biosci. (Landmark Ed) 2020, 25, 893–911. [Google Scholar] [CrossRef] [PubMed]
- Romeo, A.M.; Christen, L.; Niles, E.G.; Kosman, D.J. Intracellular chelation of iron by bipyridyl inhibits DNA virus replication: Ribonucleotide reductase maturation as a probe of intracellular iron pools. J. Biol. Chem. 2001, 276, 24301–24308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, S.; She, H.; Takeuchi, H.; Han, B.; Engelhardt, J.F.; Barton, C.H.; Zandi, E.; Giulivi, C.; Tsukamoto, H. Signaling role of intracellular iron in NF-kappaB activation. J. Biol. Chem. 2003, 278, 17646–17654. [Google Scholar] [CrossRef] [Green Version]
- Sienkiewicz, M.; Jaśkiewicz, A.; Tarasiuk, A.; Fichna, J. Lactoferrin: An overview of its main functions, immunomodulatory and antimicrobial role, and clinical significance. Crit. Rev. Food Sci. Nutr. 2021, 8, 1–18. [Google Scholar] [CrossRef]
- Recalcati, S.; Locati, M.; Marini, A.; Santambrogio, P.; Zaninotto, F.; De Pizzol, M.; Zammataro, L.; Girelli, D.; Cairo, G. Differential regulation of iron homeostasis during human macrophage polarized activation. Eur. J. Immunol. 2010, 40, 824–835. [Google Scholar] [CrossRef]
- Cutone, A.; Rosa, L.; Lepanto, M.S.; Scotti, M.J.; Berlutti, F.; Bonaccorsi di Patti, M.C.; Musci, G.; Valenti, P. Lactoferrin Efficiently Counteracts the Inflammation-Induced Changes of the Iron Homeostasis System in Macrophages. Front. Immunol. 2017, 8, 705. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.B.; Nguyen, N.B.; Marquess, K.D.; Yang, F.; Haile, D.J. Regulation of hepcidin and ferroportin expression by lipopolysaccharide in splenic macrophages. Blood Cells Mol. Dis. 2005, 35, 47–56. [Google Scholar] [CrossRef]
- Willemetz, A.; Beatty, S.; Richer, E.; Rubio, A.; Auriac, A.; Milkereit, R.J.; Thibaudeau, O.; Vaulont, S.; Malo, D.; Canonne-Hergaux, F. Iron- and Hepcidin-Independent Downregulation of the Iron Exporter Ferroportin in Macrophages during Salmonella Infection. Front. Immunol. 2017, 8, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepanto, M.S.; Rosa, L.; Cutone, A.; Conte, M.P.; Paesano, R.; Valenti, P. Efficacy of Lactoferrin Oral Administration in the Treatment of Anemia and Anemia of Inflammation in Pregnant and Non-pregnant Women: An Interventional Study. Front. Immunol. 2018, 9, 2123. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, B.; Sampath, P.; Fox, P.L. Translational control of ceruloplasmin gene expression: Beyond the IRE. Biol. Res. 2006, 39, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Marques, L.; Auriac, A.; Willemetz, A.; Banha, J.; Silva, B.; Canonne-Hergaux, F.; Costa, L. Immune cells and hepatocytes express glycosylphosphatidylinositol-anchored ceruloplasmin at their cell surface. Blood Cells Mol. Dis. 2012, 48, 110–120. [Google Scholar] [CrossRef]
- Tisato, V.; Gallo, S.; Melloni, E.; Celeghini, C.; Passaro, A.; Zauli, G.; Secchiero, P.; Bergamini, C.; Trentini, A.; Bonaccorsi, G.; et al. TRAIL and Ceruloplasmin Inverse Correlation as a Representative Crosstalk between Inflammation and Oxidative Stress. Mediators Inflamm. 2018, 2018, 9629537. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, B.; Mukhopadhyay, C.K.; Prok, A.; Cathcart, M.K.; Fox, P.L. Induction of ceruloplasmin synthesis by IFN-gamma in human monocytic cells. J. Immunol. 1997, 159, 1938–1944. [Google Scholar] [PubMed]
- Persichini, T.; Maio, N.; di Patti, M.C.; Rizzo, G.; Toscano, S.; Colasanti, M.; Musci, G. Interleukin-1β induces ceruloplasmin and ferroportin-1 gene expression via MAP kinases and C/EBPβ, AP-1, and NF-κB activation. Neurosci. Lett. 2010, 484, 133–138. [Google Scholar] [CrossRef]
- Sheikh, N.; Dudas, J.; Ramadori, G. Changes of gene expression of iron regulatory proteins during turpentine oil-induced acute-phase response in the rat. Lab. Investig. 2007, 87, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Bergamaschi, G.; Di Sabatino, A.; Pasini, A.; Ubezio, C.; Costanzo, F.; Grataroli, D.; Masotti, M.; Alvisi, C.; Corazza, G.R. Intestinal expression of genes implicated in iron absorption and their regulation by hepcidin. Clin. Nutr. 2017, 36, 1427–1433. [Google Scholar] [CrossRef]
- Malik, I.A.; Naz, N.; Sheikh, N.; Khan, S.; Moriconi, F.; Blaschke., M.; Ramadori, G. Comparison of changes in gene expression of transferrin receptor-1 and other iron-regulatory proteins in rat liver and brain during acute-phase response. Cell. Tissue Res. 2011, 344, 299–312. [Google Scholar] [CrossRef]
- Wesselius, L.J.; Nelson, M.E.; Skikne, B.S. Increased release of ferritin and iron by iron-loaded alveolar macrophages in cigarette smokers. Am. J. Respir. Crit. Care Med. 1994, 150, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.N.; Eubanks, S.K.; Schaffer, K.J. Zhou, C.Y.; Linder, M.C. Secretion of ferritin by rat hepatoma cells and its regulation by inflammatory cytokines and iron. Blood 1997, 90, 4979–4986. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Phua, J.; Weng, L.; Ling, L.; Egi, M.; Lim, C.M.; Divatia, J.V.; Shrestha, B.R.; Arabi, Y.M.; Ng, J.; Gomersall, C.D.; et al. Intensive care management of coronavirus disease 2019 (COVID-19): Challenges and recommendations. Lancet Respir. Med. 2020, 8, 506–517. [Google Scholar] [CrossRef]
- Frioni, A.; Conte, M.P.; Cutone, A.; Longhi, C.; Musci, G.; di Patti, M.C.; Natalizi, T.; Marazzato, M.; Lepanto, M.S.; Puddu, P.; et al. Lactoferrin differently modulates the inflammatory response in epithelial models mimicking human inflammatory and infectious diseases. Biometals 2014, 27, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Cutone, A.; Lepanto, M.S.; Rosa, L.; Scotti, M.J.; Rossi, A.; Ranucci, S.; De Fino, I.; Bragonzi, A.; Valenti, P.; Musci, G.; et al. Aerosolized Bovine Lactoferrin Counteracts Infection, Inflammation and Iron Dysbalance in A Cystic Fibrosis Mouse Model of Pseudomonas aeruginosa Chronic Lung Infection. Int. J. Mol. Sci. 2019, 20, 2128. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, H.; Tong, X.; Kawanami, T.; Wano, Y.; Hirose, Y.; Sugai, S.; Koeffler, H.P. Analyses for binding of the transferrin family of proteins to the transferrin receptor 2. Br. J. Haematol. 2004, 127, 464–473. [Google Scholar] [CrossRef]
- Rosa, L.; Tripepi, G.; Naldi, E.; Aimati, M.; Santangeli, S.; Venditto, F.; Caldarelli, M.; Valenti, P. Ambulatory COVID-19 Patients Treated with Lactoferrin as a Supplementary Antiviral Agent: A Preliminary Study. J. Clin. Med. 2021, 10, 4276. [Google Scholar] [CrossRef]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin as Antiviral Treatment in COVID-19 Management: Preliminary Evidence. Int. J. Environ. Res. Public Health 2021, 18, 10985. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant of Concern | Defining Mutations |
|---|---|
| B.1.1.7 (Alpha) | ΔH69-V70, ΔY144, N501Y, A570D, D614G, P681H, T716I, S982A, D1118H |
| B.1.351 (Beta) | D80A, D215G, ΔL241-L242-A243, K417N, E484K, N501Y, D614G, A701V |
| B.1.617.2 (Delta) | T19R, ΔE156-F157, R158G, L452R, T478K, D614G, P681R, D950N |
| B.1.1.529 (Omicron) | A67V, ΔH69-V70, T95I, ΔG142-V143-Y144, Y145D, ΔN211, L212I, G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F |
| TfR1-hLf | TfR1-bLf |
|---|---|
| Hydrogen bonds | Hydrogen bonds |
| K385.B–Q512 | / |
| R646.A–C371 | / |
| S654.A–E388 | / |
| R121.A–Q165 | / |
| G661.A–Y65 | / |
| Y123.A–Q165 | / |
| Salt bridges | Salt bridges |
| D755.A–K73 | D245.B–K27 |
| K508.A–E335 | D352.B–R21 |
| K508.A–E336 | D356.B–K28 |
| E623.A -E366 | K205.B–E176 |
| R183.B–E514 | R208.B–E178 |
| K385.B–E514 | E369.B–R186 |
| E606.A–R332 | E369.B–R38 |
| E612.A–K333 | / |
| R629.A–E637 | / |
| K633.A–E637 | / |
| E664.A–R120 | / |
| Non-polar contacts | Non-polar contacts |
| S120.A: P167 | D245: K28 |
| Y123.A: P144, E146, A147, F166 | Y247: R20, Q23, W24, K27 |
| W124.A: T139, E143 | T248: R20, W24 |
| D125.A: E143, R151 | E350: Q13, W16, F17, R20 |
| K508.A: T139, S334 | G351: F17, W24 |
| Q511.A: E336 | D352: R25, S285 |
| K600.A: P142 | C353: R25 |
| N608.A: P142 | P354: R25, K28 |
| L619.A: N359, S362, G363, G367, T370 | S355: R25 |
| R623.A: Q360, G363, L364 | M365: W24 |
| D626.A: Q360 | V366: R20, W24 |
| R629.A: S636 | E369: W16 |
| Q640.A: E352, E353, R356 | / |
| Y643.A: L355, R356, N359 | / |
| R646.A: L355, N359 | / |
| G647.A: L355 | / |
| F650.A: V346, T370, C371, S372 | / |
| R651.A: S373 | / |
| T658.A: E388 | / |
| F660.A: R332 | / |
| G661.A: I328 | / |
| D662.A: Y65, L69 | / |
| A663.A: L69 | / |
| E664.A: L69 | / |
| K665.A: R332 | / |
| V670.A: A70 | / |
| E163.B: Q512, G513, E514 | / |
| K177.B: N52, N261 | / |
| Q185.B: E514 | / |
| F187.B: Q512 | / |
| K394.B: P71, Y72 | / |
| MM/GBSA Results | |||||
|---|---|---|---|---|---|
| Variant | VdW (kcal/mol) | Electrostatic (kcal/mol) | Nonpolar Solvation (kcal/mol) | Polar Solvation (kcal/mol) | ΔGbinding (kcal/mol) |
| B.1.1.7 | −181.5 ± 11.1 | −22.9 ± 47.6 | −22.6 ± 1.8 | 190.7 ± 49.5 | −36.2 ± 8.8 |
| B.1.351 | −175.3 ± 12.4 | 280.1 ± 56.0 | −22.5 ± 1.8 | −151.4 ± 54.8 | −69.1 ± 13.5 |
| B.1.617.2 | −164.9 ± 9.2 | 473.13 ± 56.5 | −22.1 ± 1.2 | −332.4 ± 53.1 | −46.4 ± 8.3 |
| B.1.1.529 | −156.7 ± 10.1 | 836.9 ± 51.6 | −21.6 ± 1.1 | −758.8 ± 47.7 | −45.8 ± 11.0 |
| B.1.1.7 (Alpha) | B.1.351 (Beta) | B.1.617.2 (Delta) | B.1.1.529 (Omicron) |
|---|---|---|---|
| Salt bridges | |||
| D443–K458A | K358–D398A | K358–D405A | D646–K440B |
| D126–K444A | E352–K444C | K642–D467A | E355–R498C |
| E574–K444B | E407–K444C | E355–R409A | E356–R498C |
| D646–K444B | D646–K444B | E654–R498B | |
| E355–K378A | K358–D405A | ||
| Hydrogen bonds | |||
| N349–Y501A | E356- N449C | Q386–N439C | K358–D405A |
| Q628–E406A | S160–F490C | N387–T500C | E356–R498C |
| H439–Y439A | S381–N440C | D646–K444B | E355–R498C |
| Q386–N465C | Q378–N440C | Q386–Q506C | E356–S496C |
| C390–N440C | D646–K440B | ||
| E355–R408A | R363–F497C | ||
| Q386–Q506C | |||
| B.1.1.7 (Alpha) | B.1.351 (Beta) | B.1.617.2 (Delta) | B.1.1.529 (Omicron) |
|---|---|---|---|
| Non-Polar Contacts | |||
| K627: D402A, R405A, Q411A, T412A, G413A | C377: W436C, N437C, S438C, N439C, N440C | G641: F462A, R464A | T576: F494B |
| L630: Q411A | S381: W436C, S438C, Y505C | K642: N437B | C644: N436B |
| L631: P409A, G410A, Q411A, P497C | W380: S438C, N439C | C644: S441B, P497B | P645: N436B |
| H632: P497C | G385: S438C, G496C, F497C, Q498C, P499C, T500C, Y505C | P645: S436B, N437B, D440B, S441B, F495B, Q496B, P497B, T498B | S653: F494B, R495B, P496B |
| Q634: Q411A | K376: N439C | D646: S436B, N437B, P497B, T498B, Y503B | L151: G499A |
| A635: P497C, T498C | N387: N439C | K647: P497B | R152: R495A, P496A, T497A, Y498A, G499A |
| L636: T498C | V388: N439C, K444C, G496C | F648: P497B | P153: R495A, P496A, T497A |
| K642: T498C | T389: N439C, K444C | F651: F495B, Q496B, P497B | L155: R495A |
| P645: G499B | E373: N440C | S653: F495B, Q496B | S156: G499A |
| D646: T498C, G500C | E355: S443C, K444C, G446C, G447C, Y495C, G496C | L125: Q496A | W157: H502A |
| S653: P496B | T362: S443C, K444C, V445C, G446C, G447C, G496C, F497C, Q498C | L151: G500A | E159: C477C |
| A668: D417A | A359: K444C, V445C, G446C | R152: N499A, G500A, G502A, Y503A | S160: P476C, C477C, N484C |
| Q124: Q495A | V364: K444C, V445C | P153: Q496A, G502A, Y503A | T353: G499A, V500A |
| L125: S440A | C390: K444C | L155: W434A | A354: G401A, F494C |
| M148: P496A, T497A | G406: K444C, V445C | W157: C486C | E355: Y492C, F494C |
| G149: Q495A, P496A, T497A, G501A | L347: V445C | E159: C486C, L490C | V357: I399A, G401A, G499A, V500A, G501A |
| R152: G501A, Y502A | R351: V445C, G446C, G447C, N448C | S160: S369A, N485C, C486C | K358: F494C, R495C |
| P153: S435A, Q495A, G501A | E352: N448C | L251: P497A | Y361: R495C |
| C250: F494A, Q495A, P496A | A184: G476C | N252: Q496A, P497A | T362: S493C, F494C, R495C |
| L251: F494A, Q495A | S160: P479C, C480C, F486C, N487C, C488C, Y489C, S371A | L347: V443C | Q374: N436C |
| N252: Q495A | E159: C488C, Y489C, L492C | R351: G444C, N446C | C377: S435C, N436C, D439C |
| S341: T497A | K358: G496C, Q498C | E352: N446C | Q378: G499B |
| L344: T497A | S384: F497C, Q498C, P499C, T500C | T353: G500A | W380: S493C |
| T345: T497A | Y361: Q498C | A354: V405A | S381: Y498C |
| K348: T497A, Y498A | Q386: Q498C, P499C | E355: V405A, N446C, G494C | S384: F494C, R495C, P496C, T497C, Y498C |
| E356: G499A | Q383: P499C | V357: G402A, V405A | G385: S493C, F494C, R495C, P496C, T497C, H502C |
| V357: G499A | W157: S371A | K358: V405A, Q496C | Q386: R495C, P496C |
| K358: K442C, G494C | A354: F374A, V407A | Y361: P497C | N387: S493C |
| R360: T497A, Y498A, G499A | V357: V407A | T362: G445C, G494C, Q496C | V388: S493C |
| Y361: V404A, G494C, Q496C | P645: W437B, S439B, N440B, Y506B | E373: N437C | T389: D439C, S440C, K441C |
| T362: K442C, G494C | C644: S439B, N440B, S444B, Q499B | K376: N437C | C390: K441C |
| C377: N437C | D646: S439B, T501B, Y506B | C377: N435C, S436C, N437C, N438C | A391: K441C |
| S381: S436C, N437C, D440C, Y499C, Y503C | S575: F498B | Q378: P497B, T498B, N499B | V401: K441C |
| Q382: P497C, T498C, Y499C | S653: F498B, T501B | W380: S436C, N437C | G406: K441C |
| Q383: Q496C, P497C, T498C, Y499C | F648: Q499B, P500B | S381: G500B, W434C, S436C, Y503C | S575: V442B, F494B |
| S384: G494C, F495C, Q496C, P497C, T498C, Y499C | F651: Q499B, P500B | Q382: G500B, Y503B | |
| G385: S436C, D440C, K442C, G494C, Q496C | K375: P500B | S384: Q496C, P497C, T498C, N499C | |
| Q386: Q496C, P497C | Q378: P500B, T501B, Y502B, G503B | G385: W434C, S436C, G494C, F495C, Q496C, P497C, T498C, Y503C | |
| S437: E468A, I469A | K647: P500B | Q386: Q496C, P497C, T498C | |
| K438: E468A, I469A | K652: P500B | V388: N437C, G494C | |
| Q382: G503B, Y506B | T389: N437C | ||
| C390: K442C | |||
| V401: K442C | |||
| G406: K442C, V443C | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cutone, A.; Rosa, L.; Bonaccorsi di Patti, M.C.; Iacovelli, F.; Conte, M.P.; Ianiro, G.; Romeo, A.; Campione, E.; Bianchi, L.; Valenti, P.; et al. Lactoferrin Binding to SARS-CoV-2 Spike Glycoprotein Blocks Pseudoviral Entry and Relieves Iron Protein Dysregulation in Several In Vitro Models. Pharmaceutics 2022, 14, 2111. https://doi.org/10.3390/pharmaceutics14102111
Cutone A, Rosa L, Bonaccorsi di Patti MC, Iacovelli F, Conte MP, Ianiro G, Romeo A, Campione E, Bianchi L, Valenti P, et al. Lactoferrin Binding to SARS-CoV-2 Spike Glycoprotein Blocks Pseudoviral Entry and Relieves Iron Protein Dysregulation in Several In Vitro Models. Pharmaceutics. 2022; 14(10):2111. https://doi.org/10.3390/pharmaceutics14102111
Chicago/Turabian StyleCutone, Antimo, Luigi Rosa, Maria Carmela Bonaccorsi di Patti, Federico Iacovelli, Maria Pia Conte, Giusi Ianiro, Alice Romeo, Elena Campione, Luca Bianchi, Piera Valenti, and et al. 2022. "Lactoferrin Binding to SARS-CoV-2 Spike Glycoprotein Blocks Pseudoviral Entry and Relieves Iron Protein Dysregulation in Several In Vitro Models" Pharmaceutics 14, no. 10: 2111. https://doi.org/10.3390/pharmaceutics14102111
APA StyleCutone, A., Rosa, L., Bonaccorsi di Patti, M. C., Iacovelli, F., Conte, M. P., Ianiro, G., Romeo, A., Campione, E., Bianchi, L., Valenti, P., Falconi, M., & Musci, G. (2022). Lactoferrin Binding to SARS-CoV-2 Spike Glycoprotein Blocks Pseudoviral Entry and Relieves Iron Protein Dysregulation in Several In Vitro Models. Pharmaceutics, 14(10), 2111. https://doi.org/10.3390/pharmaceutics14102111