
Ramosin: The First Antibacterial Peptide Identified on Bolitoglossa ramosi Colombian Salamander

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis
2.1.1. Transcriptome
2.1.2. Protein Prediction from the De Novo Transcriptome of Bolitoglossa ramosi
2.1.3. APD3 Peptide Database
2.1.4. Alignments to Identify Candidate Peptides
2.1.5. In Silico Prediction of the Antimicrobial Activity of Candidate Peptides
2.1.6. Molecular Confirmation of the Candidate Peptides
2.2. Peptide Synthesis and Characterization
2.2.1. Determination of the Main Fraction Containing the Expected Peptide
2.2.2. Circular Dichroism
2.3. Antimicrobial Activity
2.4. Hemolytic Assay
2.5. Scanning Electron Microscopy (SEM)
2.6. Cell Lines
2.7. In Vitro Cytotoxicity Assay
2.8. Statistic Test
3. Results
3.1. In Silico Identification of AMPs from B. ramosi Salamander
3.2. Assays of Antibacterial Activity of Ramosin Peptide from B. ramosi
3.2.1. Antibacterial Assay of Ramosin Peptide
3.2.2. Hemolytic Assay of Ramosin Peptide
3.2.3. Scanning Electron Microscopy Assay with Ramosin Peptide
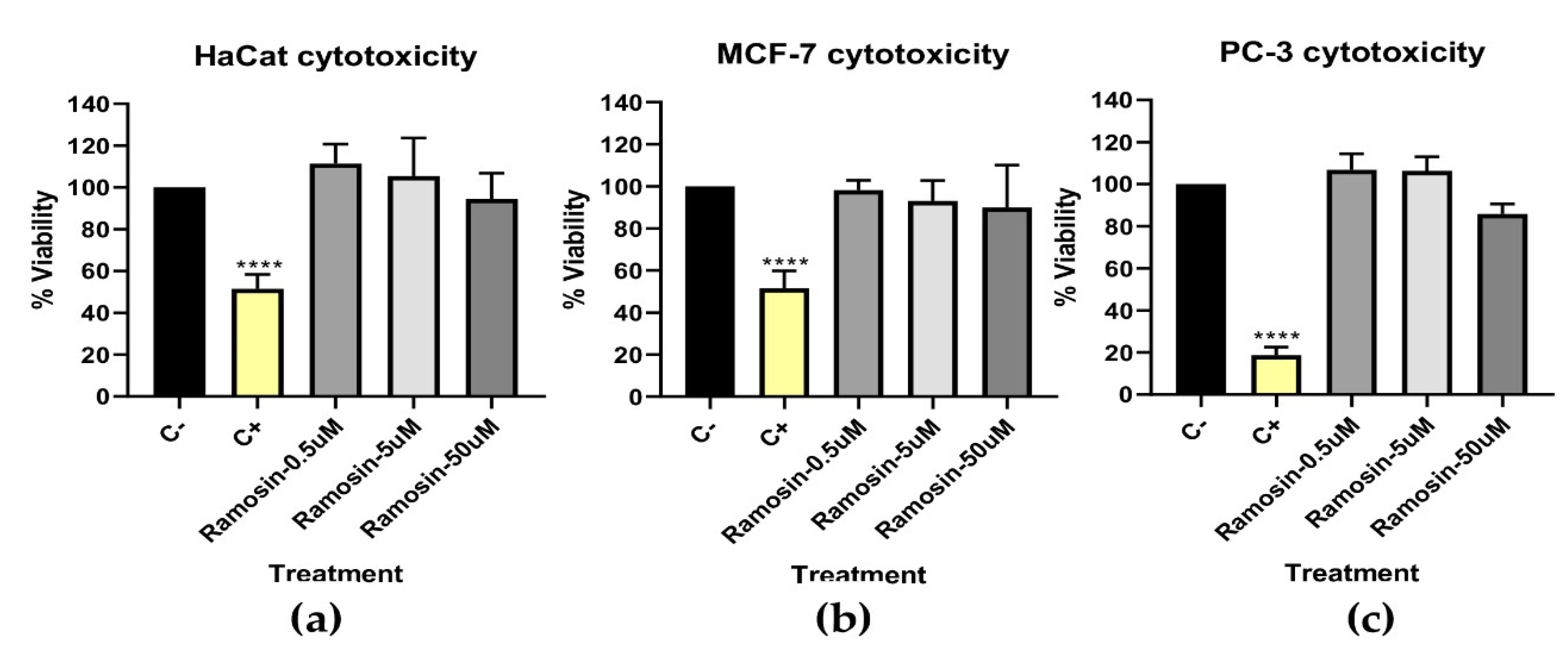
3.2.4. In Vitro Cytotoxicity Assay with MTT
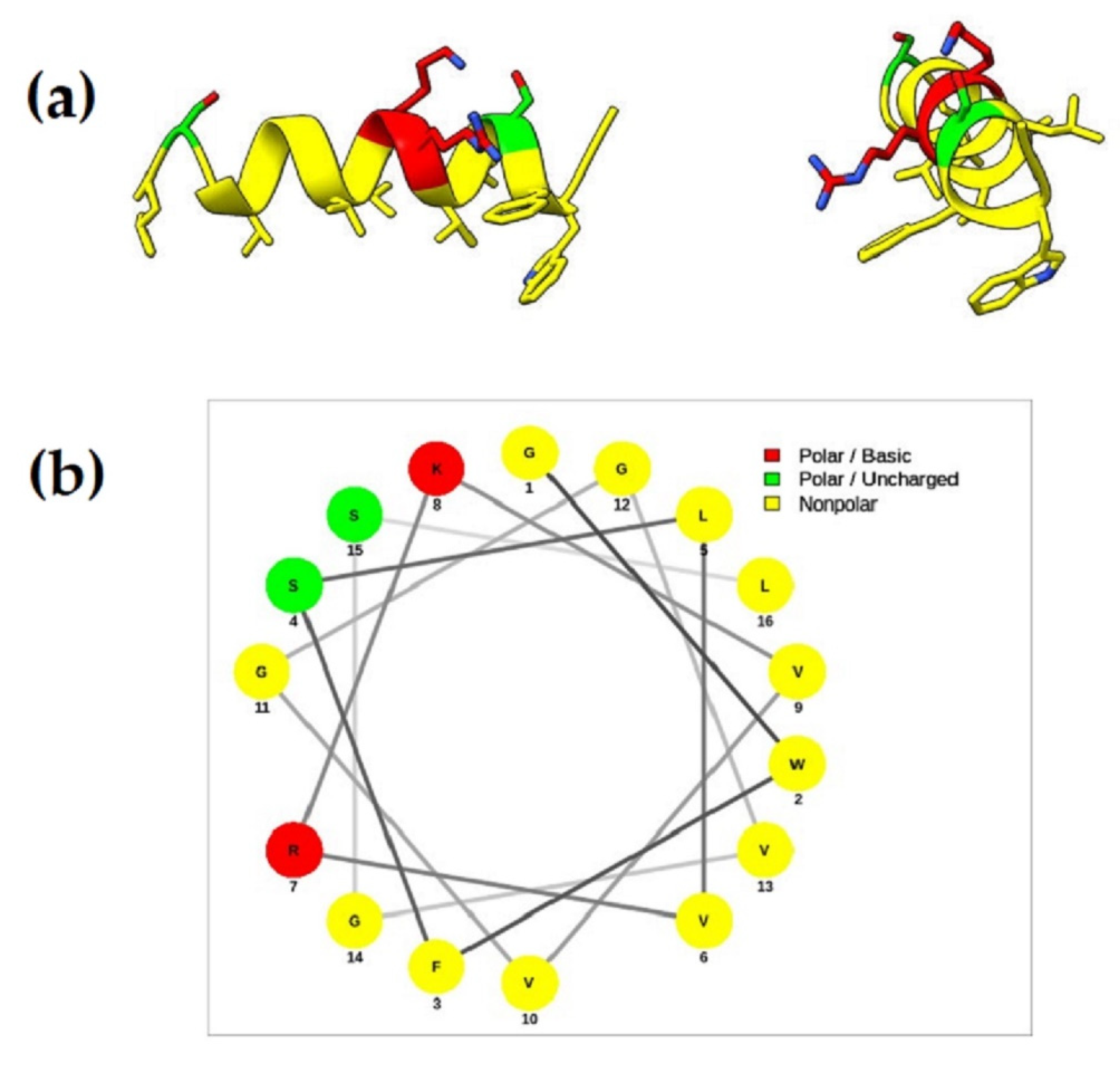
3.2.5. Molecular Characterization of Ramosin Peptide
3.2.6. Chemical Characterization of Ramosin Peptide of B. ramosi
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Protocol for the Screening Test
References
- Yeung, A.T.Y.; Gellatly, S.L.; Hancock, R.E.W. Multifunctional Cationic Host Defence Peptides and Their Clinical Applications. Cell. Mol. Life Sci. 2011, 68, 2161–2176. [Google Scholar] [CrossRef] [PubMed]
- Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Antimicrobial Peptides: Versatile Biological Properties. Int. J. Pept. 2013, 2013, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felício, M.R.; Silva, O.N.; Gonçalves, S.; Santos, N.C.; Franco, O.L. Peptides with Dual Antimicrobial and Anticancer Activities. Front. Chem. 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of Antimicrobial Peptide Action and Resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-K.; Kim, C.; Seo, C.H.; Park, Y. The Therapeutic Applications of Antimicrobial Peptides (AMPs): A Patent Review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef]
- Zanetti, M. Cathelicidins, Multifunctional Peptides of the Innate Immunity. J. Leukoc. Biol. 2004, 75, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Patrzykat, A. Clinical Development of Cationic Antimicrobial Peptides: From Natural to Novel Antibiotics. Curr. Drug Targets Infect. Disord. 2002, 2, 79–83. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K. The Roles of Antimicrobial Peptides in Innate Host Defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [Green Version]
- Ladram, A. Antimicrobial Peptides from Frog Skin Biodiversity and Therapeutic Promises. Front. Biosci. 2016, 21, 4461. [Google Scholar] [CrossRef]
- Toledo, R.C.; Jared, C. Cutaneous Granular Glands and Amphibian Venoms. Comp. Biochem. Physiol. Part A Physiol. 1995, 111, 1–29. [Google Scholar] [CrossRef]
- Conlon, J.M. Structural Diversity and Species Distribution of Host-Defense Peptides in Frog Skin Secretions. Cell. Mol. Life Sci. 2011, 68, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, R. The Chemistry and Biological Activities of Peptides from Amphibian Skin Secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, S.; Falanga, A.; Berisio, R.; Grieco, P.; Morelli, G.; Galdiero, M. Antimicrobial Peptides as an Opportunity Against Bacterial Diseases. Curr. Med. Chem. 2015, 22, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Chen, Y.; Yao, H.; Du, C.; Luan, N.; Yan, X. A Novel Defensin-like Antimicrobial Peptide from the Skin Secretions of the Tree Frog, Theloderma Kwangsiensis. Gene 2016, 576, 136–140. [Google Scholar] [CrossRef]
- Pei, J.; Jiang, L. Antimicrobial Peptide from Mucus of Andrias Davidianus: Screening and Purification by Magnetic Cell Membrane Separation Technique. Int. J. Antimicrob. Agents 2017, 50, 41–46. [Google Scholar] [CrossRef]
- Demori, I.; El Rashed, Z.; Corradino, V.; Catalano, A.; Rovegno, L.; Queirolo, L.; Salvidio, S.; Biggi, E.; Zanotti-Russo, M.; Canesi, L.; et al. Peptides for Skin Protection and Healing in Amphibians. Molecules 2019, 24, 347. [Google Scholar] [CrossRef] [Green Version]
- Lüddecke, T.; Schulz, S.; Steinfartz, S.; Vences, M. A Salamander’s Toxic Arsenal: Review of Skin Poison Diversity and Function in True Salamanders, Genus Salamandra. Sci. Nat. 2018, 105, 56. [Google Scholar] [CrossRef]
- Mu, L.; Tang, J.; Liu, H.; Shen, C.; Rong, M.; Zhang, Z.; Lai, R. A Potential Wound-healing-promoting Peptide from Salamander Skin. FASEB J. 2014, 28, 3919–3929. [Google Scholar] [CrossRef] [Green Version]
- Von Byern, J.; Grunwald, I.; Kosok, M.; Saporito, R.A.; DIcke, U.; Wetjen, O.; Thiel, K.; Borcherding, K.; Kowalik, T.; Marchetti-Deschmann, M. Chemical Characterization of the Adhesive Secretions of the Salamander Plethodon Shermani (Caudata, Plethodontidae). Sci. Rep. 2017, 7, 6647. [Google Scholar] [CrossRef]
- Rovito, S.M.; Parra-Olea, G.; Vásquez-Almazán, C.R.; Luna-Reyes, R.; Wake, D.B. Deep Divergences and Extensive Phylogeographic Structure in a Clade of Lowland Tropical Salamanders. BMC Evol. Biol. 2012, 12, 255. [Google Scholar] [CrossRef] [Green Version]
- Corantioquia: Centro de Información Ambiental Koha. Detalles Para: Anfibios y Reptiles Del Valle de Aburrá. Available online: http://cia.corantioquia.gov.co/cgi-bin/koha/opac-detail.pl?biblionumber=12415 (accessed on 27 December 2020).
- Arenas Gómez, C.M.; Gómez Molina, A.; Zapata, J.D.; Delgado, J.P. Limb Regeneration in a Direct-Developing Terrestrial Salamander, Bolitoglossa Ramosi (Caudata: Plethodontidae). Regeneration 2017, 4, 227–235. [Google Scholar] [CrossRef]
- IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 27 December 2020).
- Arenas Gómez, C.M.; Woodcock, R.M.; Smith, J.J.; Voss, R.S.; Delgado, J.P. Using Transcriptomics to Enable a Plethodontid Salamander (Bolitoglossa Ramosi) for Limb Regeneration Research 06 Biological Sciences 0604 Genetics. BMC Genom. 2018, 19, 704. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.L.; Mcdermott, A.M.; Zasloff, M. Antimicrobial Peptides and Wound Healing: Biological and Therapeutic Considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schikorski, D.; Cuvillier-Hot, V.; Leippe, M.; Boidin-Wichlacz, C.; Slomianny, C.; Macagno, E.; Salzet, M.; Tasiemski, A. Microbial Challenge Promotes the Regenerative Process of the Injured Central Nervous System of the Medicinal Leech by Inducing the Synthesis of Antimicrobial Peptides in Neurons and Microglia. J. Immunol. 2008, 181, 1083–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibardi, L.; Celeghin, A.; Dalla Valle, L. Wounding in Lizards Results in the Release of Beta-Defensins at the Wound Site and Formation of an Antimicrobial Barrier. Dev. Comp. Immunol. 2012, 36, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M. Host-Defense Peptides of the Skin with Therapeutic Potential: From Hagfish to Human. Peptides 2015, 67, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Wu, H.; Huang, J.; Wang, W.; Ge, K.; Li, G.; Zhong, J.; Huang, Q. LAMP2: A Major Update of the Database Linking Antimicrobial Peptides. Database 2020, 2020, baaa061. [Google Scholar] [CrossRef]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef]
- Liscano, Y.; Medina, L.; Oñate-Garzón, J.; Gúzman, F.; Pickholz, M.; Delgado, J.P. In Silico Selection and Evaluation of Pugnins with Antibacterial and Anticancer Activity Using Skin Transcriptome of Treefrog (Boana Pugnax). Pharmaceutics 2021, 13, 578. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The Antimicrobial Peptide Database as a Tool for Research and Education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3 Database. Available online: http://aps.unmc.edu/AP/statistic/statistic.php (accessed on 27 December 2020).
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Pearson, W.R. Finding Protein and Nucleotide Similarities with FASTA. Curr. Protoc. Bioinform. 2016, 2016, 3.9.1–3.9.25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TransDecoder. Available online: https://github.com/TransDecoder/TransDecoder/wiki (accessed on 28 December 2020).
- Liscano, Y.; Oñate-Garzón, J.; Delgado, J.P. Peptides with Dual Antimicrobial–Anticancer Activity: Strategies to Overcome Peptide Limitations and Rational Design of Anticancer Peptides. Molecules 2020, 25, 4245. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Karnik, S.; Nilawe, P.; Jayaraman, V.K.; Idicula-Thomas, S. ClassAMP: A Prediction Tool for Classification of Antimicrobial Peptides. IEEE/ACM Trans. Comput. Biol. Bioinform. 2012, 9, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- Houghten, R.A. General Method for the Rapid Solid-Phase Synthesis of Large Numbers of Peptides: Specificity of Antigen-Antibody Interaction at the Level of Individual Amino Acids. Proc. Natl. Acad. Sci. USA 1985, 82, 5131–5135. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, F.; Gauna, A.; Roman, T.; Luna, O.; Álvarez, C.; Pareja-Barrueto, C.; Mercado, L.; Albericio, F.; Cárdenas, C. Tea Bags for Fmoc Solid-Phase Peptide Synthesis: An Example of Circular Economy. Molecules 2021, 26, 5035. [Google Scholar] [CrossRef]
- Santana, P.A.; Álvarez, C.A.; Guzmán, F.; Mercado, L. Development of a Sandwich ELISA for Quantifying Hepcidin in Rainbow Trout. Fish Shellfish Immunol. 2013, 35, 748–755. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Barchiesi, F.; Del Prete, M.S.; Fortuna, M.; Caselli, F.; Scalise, G. In Vitro Susceptibility Tests for Cationic Peptides: Comparison of Broth Microdilution Methods for Bacteria That Grow Aerobically. Antimicrob. Agents Chemother. 2000, 44, 1694–1696. [Google Scholar] [CrossRef]
- Tavanti, A.; Maisetta, G.; Del Gaudio, G.; Petruzzelli, R.; Sanguinetti, M.; Batoni, G.; Senesi, S. Fungicidal Activity of the Human Peptide Hepcidin 20 Alone or in Combination with Other Antifungals against Candida Glabrata Isolates. Peptides 2011, 32, 2484–2487. [Google Scholar] [CrossRef] [PubMed]
- Santana, P.A.; Salinas, N.; Álvarez, C.A.; Mercado, L.A.; Guzmán, F. Alpha-Helical Domain from IL-8 of Salmonids: Mechanism of Action and Identification of a Novel Antimicrobial Function. Biochem. Biophys. Res. Commun. 2018, 498, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Segura, C.; Guzmán, F.; Salazar, L.M.; Patarroyo, M.E.; Orduz, S.; Lemeshko, V. BTM-P1 Polycationic Peptide Biological Activity and 3D-Dimensional Structure. Biochem. Biophys. Res. Commun. 2007, 353, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Rozek, T.; Bowie, J.H.; Wallace, J.C.; Tyler, M.J. The Antibiotic and Anticancer Active Aurein Peptides from the Australian Bell Frogs Litoria Aurea and Litoria Raniformis. Part 2. Sequence Determination Using Electrospray Mass Spectrometry. Rapid Commun. Mass Spectrom. 2000, 14, 2002–2011. [Google Scholar] [CrossRef]
- Rozek, T.; Wegener, K.L.; Bowie, J.H.; Olver, I.N.; Carver, J.A.; Wallace, J.C.; Tyler, M.J. The Antibiotic and Anticancer Active Aurein Peptides from the Australian Bell Frogs Litoria Aurea and Litoria Raniformis: The Solution Structure of Aurein 1.2. Eur. J. Biochem. 2000, 267, 5330–5341. [Google Scholar] [CrossRef]
- Chang, K.Y.; Yang, J.-R. Analysis and Prediction of Highly Effective Antiviral Peptides Based on Random Forests. PLoS ONE 2013, 8, e70166. [Google Scholar] [CrossRef] [Green Version]
- Povey, J.F.; Smales, C.M.; Hassard, S.J.; Howard, M.J. Comparison of the Effects of 2,2,2-Trifluoroethanol on Peptide and Protein Structure and Function. J. Struct. Biol. 2007, 157, 329–338. [Google Scholar] [CrossRef]
- Roccatano, D.; Colombo, G.; Fioroni, M.; Mark, A.E. Mechanism by Which 2,2,2-Trifluoroethanol/Water Mixtures Stabilize Secondary-Structure Formation in Peptides: A Molecular Dynamics Study. Proc. Natl. Acad. Sci. USA 2002, 99, 12179–12184. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, Y. I-TASSER Server: New Development for Protein Structure and Function Predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure Visualization for Researchers, Educators, and Developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Mól, A.R.; Castro, M.S.; Fontes, W. NetWheels: A Web Application to Create High Quality Peptide Helical Wheel and Net Projections. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Lyu, P.; Xi, X.; Ge, L.; Mahadevappa, R.; Shaw, C.; Kwok, H.F. Triggering of Cancer Cell Cycle Arrest by a Novel Scorpion Venom-derived Peptide—Gonearrestide. J. Cell. Mol. Med. 2018, 22, 4460–4473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Lu, B.; Zhou, D.; Zhao, L.; Song, W.; Wang, L. Identification of the First Cathelicidin Gene from Skin of Chinese Giant Salamanders Andrias Davidianus with Its Potent Antimicrobial Activity. Dev. Comp. Immunol. 2017, 77, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Fredericks, L.P.; Dankert, J.R. Antibacterial and Hemolytic Activity of the Skin of the Terrestrial Salamander, Plethodon Cinereus. J. Exp. Zool. 2000, 287, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Sheafor, B.; Davidson, E.W.; Parr, L.; Rollins-Smith, L. Antimicrobial Peptide Defenses in the Salamander, Ambystoma Tigrinum, against Emerging Amphibian Pathogens. J. Wildl. Dis. 2008, 44, 226–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, P.; Yang, S.; Shen, C.; Jiang, K.; Rong, M.; Lai, R. The First Salamander Defensin Antimicrobial Peptide. PLoS ONE 2013, 8, e83044. [Google Scholar] [CrossRef] [PubMed]
- Chernysh, S.; Cociancich, S.; Briand, J.-P.; Hetru, C.; Bulet, P. The Inducible Antibacterial Peptides of the Hemipteran Insect Palomena Prasina: Identification of a Unique Family of Prolinerich Peptides and of a Novel Insect Defensin. J. Insect Physiol. 1996, 42, 81–89. [Google Scholar] [CrossRef]
- Zeng, X.C.; Wang, S.X.; Zhu, Y.; Zhu, S.Y.; Li, W.X. Identification and Functional Characterization of Novel Scorpion Venom Peptides with No Disulfide Bridge from Buthus Martensii Karsch. Peptides 2004, 25, 143–150. [Google Scholar] [CrossRef]
- Scott, M.G.; Gold, M.R.; Hancock, R.E.W. Interaction of Cationic Peptides with Lipoteichoic Acid and Gram-Positive Bacteria. Infect. Immun. 1999, 67, 6445–6453. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.G.; Yan, H.; Hancock, R.E.W. Biological Properties of Structurally Related α-Helical Cationic Antimicrobial Peptides. Infect. Immun. 1999, 67, 2005–2009. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, M.; Wang, L.; McGrath, S.; Chen, T.; Chen, X.; Shaw, C. Phylloseptin-1 (PSN-1) from Phyllomedusa Sauvagei Skin Secretion: A Novel Broad-Spectrum Antimicrobial Peptide with Antibiofilm Activity. Mol. Immunol. 2010, 47, 2030–2037. [Google Scholar] [CrossRef] [PubMed]
- Ebbensgaard, A.; Mordhorst, H.; Aarestrup, F.M.; Hansen, E.B. The Role of Outer Membrane Proteins and Lipopolysaccharides for the Sensitivity of Escherichia Coli to Antimicrobial Peptides. Front. Microbiol. 2018, 9, 2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial Host Defence Peptides: Functions and Clinical Potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Leite, J.R.S.A.; Silva, L.P.; Rodrigues, M.I.S.; Prates, M.V.; Brand, G.D.; Lacava, B.M.; Azevedo, R.B.; Bocca, A.L.; Albuquerque, S.; Bloch, C. Phylloseptins: A Novel Class of Anti-Bacterial and Anti-Protozoan Peptides from the Phyllomedusa Genus. Peptides 2005, 26, 565–573. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| APD3 | Length (aa*) | Net Charge | % Hydrophobicity | Peptide Source | Activity of the Reported Peptides on APD3 Database [34] |
|---|---|---|---|---|---|
| AP00176 | 30 | +3 | 53 | Neutrophils; natural killer cells, monocytes; saliva; Homo sapiens | Anti-Gram+ & Gram-, antiviral, antifungal, antiparasitic, anti-HIV, chemotactic, anti-**MRSA, anti-toxin, enzyme inhibitor, anti-sepsis, wound-healing, anticancer |
| AP00177 | 29 | +3 | 51 | Neutrophils; natural killer cells, monocytes; saliva; Homo sapiens | Anti-Gram+ & Gram-, antiviral, antifungal, anti-HIV, chemotactic, anti-toxin, enzyme inhibitor, anticancer |
| AP00208 | 32 | +5 | 28 | Pig, Sus scrofa | anti-Gram+ |
| AP00308 | 21 | +6 | 33 | Bufo bufo gargarizans. | Anti-Gram+ & Gram-, antifungal, candidacidal, antiparasitic, anti-sepsis, anticancer |
| AP00336 | 12 | 0 | 50 | Rainbow trout, Oncorhynchus mykiss | Anti-Gram+, antifungal |
| AP00449 | 13 | +1 | 30 | Brain, Homo sapiens | Anti-Gram+, antiviral, antifungal, candidacidal, anti-HIV |
| AP00528 | 7 | −7 | 0 | Ovis aries | Anti-Gram+ & Gram- |
| AP02017 | 31 | +5 | 45 | Placental tissue, Homo sapiens | Antifungal, candidacidal |
| AP02030 | 74 | 0 | 33 | Gill, Pacific oyster, Crassostrea gigas | Anti-Gram+ & Gram- |
| AP02096 | 59 | +19 | 25 | Homo sapiens; cytosol, macrophage, RAW264.7, Mus musculus; Rattus norvegicus | Anti-Gram+ & Gram-, anti-MRSA |
| AP02231 | 19 | +2 | 26 | Corneas, eyes, Homo sapiens | Anti-Gram- |
| AP02257 | 130 | +8 | 40 | Secretions and tissues, tears, saliva, human milk, and mucus; Homo sapiens | Anti-Gram+ & Gram-, antifungal, synergistic AMPs |
| AP02343 | 99 | −2 | 31 | Human amniotic fluid, Homo sapiens | Anti-Gram+ & Gram- |
| AP02441 | 6 | 0 | 16 | Streptomyces amritsarensis | Anti-Gram+, anti-MRSA |
| AP02791 | 22 | +7 | 22 | Blood plasma, Varanus komodoensis | Anti-Gram+ & Gram- |
| AP02807 | 103 | +18 | 33 | American cupped oysters, Crassostrea virginica | Anti-Gram- |
| AP02813 | 24 | +3 | 41 | Leukocytes; the Russian Sturgeon, Acipenser gueldenstaedtii | Anti-Gram- |
| AP02884 | 7 | −6 | 0 | Ovis aries | Anti-Gram- |
| AP02885 | 7 | −5 | 14 | Ovis aries | Anti-Gram- |
| AP02984 | 6 | −1 | 33 | Bovine milk digestion by bacteria, Bos taurus | Anti-Gram+ |
| AP03140 | 13 | 0 | 53 | Excretions and Secretions, Sarconesiopsis magellanica | Anti-Gram+ & Gram- |
| AP03151 | 20 | +7 | 35 | Fragments of a human antimicrobial protein | Anti-Gram+ & Gram-, antifungal, candidacidal, anti-inflammatory, anti-sepsis |
| AP03159 | 20 | +1 | 50 | Endometrial fluid peptides, Homo sapiens | Anti-Gram+ & Gram-, antifungal, anti-MRSA |
| Candidate Peptide Code | Peptide Sequences Synthesized | Net Charge at pH 7 | MW (g/mol) | Family | APD3 Similar Peptide | (%) Similarity with APD3 Peptides * |
|---|---|---|---|---|---|---|
| 3412 | GWFSLVRKVVGGVGSL-NH2 | +3 | 1659.96 | Aurein | AP00016 | 93.8 |
| 3413 | KQYQLVERIIGSIGSL-NH2 | +2 | 1803.1 | Aurein | AP00016 | 87.5 |
| 3414 | GMLMMVRRPFGPFGSI-NH2 | +3 | 1795.24 | Aurein | AP00014 | 87.5 |
| 3415 | QLHDVMKRVAKSF-NH2 | +3 | 1557.85 | Aurein | AP00013 | 84.6 |
| 3416 | GAFDDVKKVATTI-NH2 | +1 | 1363.55 | Aurein | AP00012 | 80 |
| 3417 | SLLSAWGKILGSKLNEKLTQ-NH2 | +3 | 2185.55 | Dahlein | AP00703 | 80.1 |
| 3418 | GFLNYYRRFIGSFAEVVT-NH2 | +2 | 2138.42 | Maculatin | AP00262 | 83.3 |
| 3419 | VLSPSLGSLAGVLGGVLKLA-NH2 | +2 | 1850.24 | Maximin | AP00832 | 85 |
| 3420 | RVRLEACVRGICRRNCK-NH2 | +6 | 2031.48 | Tachyplesin | AP00214 | 80.3 |
| 3424 | GIFTLIHCSLEGKVKKIECS-NH2 | +2 | 2204.64 | Odorranain-J1 | AP01298 | 80 |
| 3428 | AVAGRSQGQ-NH2 | +2 | 871.94 | Cn-AMP1 | AP01342 | 100 |
| Sample | Code | Number of reads | % Discordance |
|---|---|---|---|
| Control | BRE001 | 6 | 1.32 |
| BRE002 | 4 | 0.248 | |
| BRE003 | 22 | 5.176 | |
| Limb | BR006 | 18 | 4.125 |
| BR007 | 18 | 0.495 | |
| Blastema | BR004 | 8 | 0.866 |
| BR005 | 2 | 0.495 | |
| BR008 | 10 | 0.693 | |
| BR009 | 0 | 0 | |
| BR010 | 4 | 0.99 | |
| Intestine | BRI001 | 20 | 1.139 |
| Skin | BRP001 | 2 | 1.98 |
| Physicochemical Properties of Ramosin Peptide | |
|---|---|
| Sequence of amino acids | GWFSLVRKVVGGVGSL-NH2 |
| Number of amino acids | 16 |
| Molecular weight | 1659.96 |
| Chemical formula | C78H126N22O18 |
| Isoelectric point | 11.48 |
| Net charge | +3 |
| GRAVY | 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina, L.; Guzmán, F.; Álvarez, C.; Delgado, J.P.; Carbonell-M, B. Ramosin: The First Antibacterial Peptide Identified on Bolitoglossa ramosi Colombian Salamander. Pharmaceutics 2022, 14, 2579. https://doi.org/10.3390/pharmaceutics14122579
Medina L, Guzmán F, Álvarez C, Delgado JP, Carbonell-M B. Ramosin: The First Antibacterial Peptide Identified on Bolitoglossa ramosi Colombian Salamander. Pharmaceutics. 2022; 14(12):2579. https://doi.org/10.3390/pharmaceutics14122579
Chicago/Turabian StyleMedina, Laura, Fanny Guzmán, Claudio Álvarez, Jean Paul Delgado, and Belfran Carbonell-M. 2022. "Ramosin: The First Antibacterial Peptide Identified on Bolitoglossa ramosi Colombian Salamander" Pharmaceutics 14, no. 12: 2579. https://doi.org/10.3390/pharmaceutics14122579
APA StyleMedina, L., Guzmán, F., Álvarez, C., Delgado, J. P., & Carbonell-M, B. (2022). Ramosin: The First Antibacterial Peptide Identified on Bolitoglossa ramosi Colombian Salamander. Pharmaceutics, 14(12), 2579. https://doi.org/10.3390/pharmaceutics14122579