
Chemoprevention of Urothelial Cell Carcinoma Tumorigenesis by Dietary Flavokawain A in UPII-Mutant Ha-ras Transgenic Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Breeding and Genotyping
2.2. FKA Diet, Experimental Groups, and Animal Care
2.3. FKA Measurement
2.4. Necropsy, Tissue Processing, and Histology Analysis
2.5. IHC and DeadEnd Colorimetric TUNEL Assay
2.6. Analysis of Urine
2.7. Statistical Analysis
3. Results
3.1. Homozygous UPII-Mutant Ha-ras Mice That Were Fed with a FKA Containing Diet Have a Better Survival than Those Fed with Control Diet
3.2. Dietary FKA Inhibits Mutant Ha-Ras Initiated Tumorigenesis in the Bladder of Mice
3.3. Dietary FKA Reduces the Incidence of Low-Grade Papillary Carcinomas in Male UPII-Mutant Ha-Ras Transgenic Mice
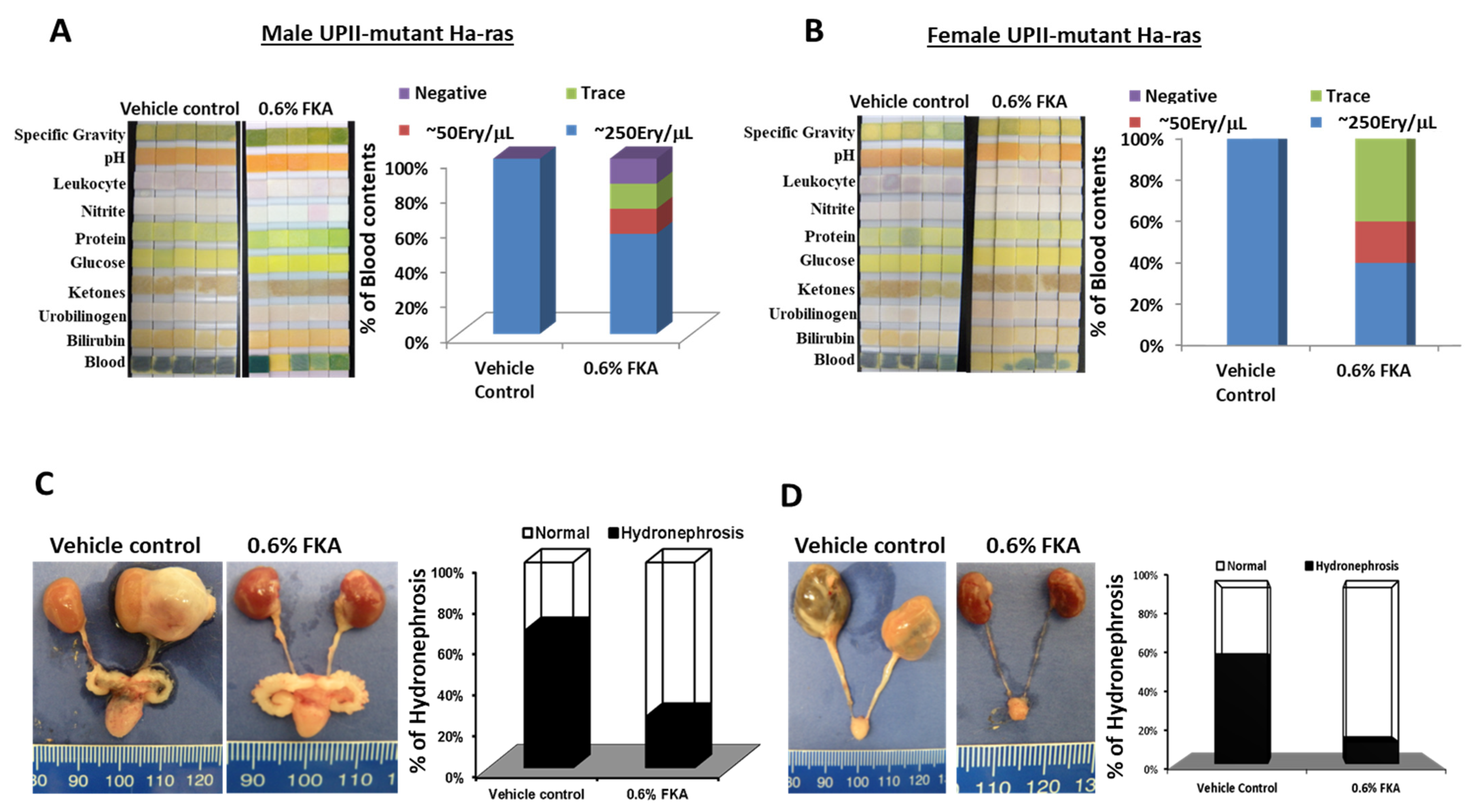
3.4. Dietary FKA Decreases Hematuria and Hydronephrosis
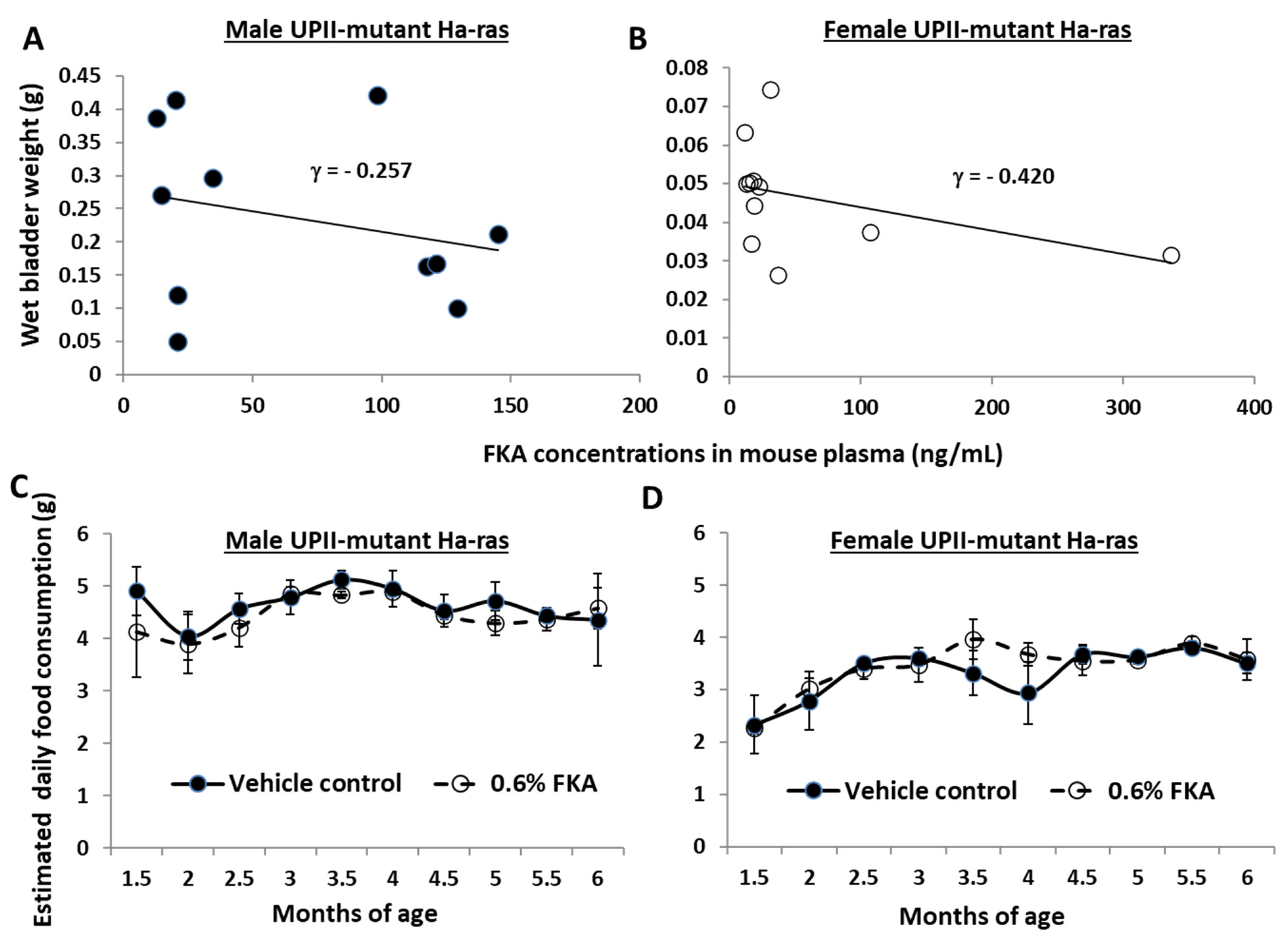
3.5. The Bladder Weights (a Surrogate of Tumor Burdens) of FKA-Diet-Fed Mice Are Inversely Related to Plasma FKA Concentrations
3.6. The In Vivo Anti-Proliferative and Apoptotic Effects of Dietary FKA
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bladder Cancer Key Statistics. Available online: https://www.cancer.net/cancer-types/bladder-cancer/statistics (accessed on 21 December 2021).
- Tran, L.; Xiao, J.F.; Agarwal, N.; Duex, J.E.; Theodorescu, D. Advances in bladder cancer biology and therapy. Nat. Rev. Cancer 2021, 21, 104–121. [Google Scholar] [CrossRef] [PubMed]
- Sievert, K.D.; Amend, B.; Nagele, U.; Schilling, D.; Bedke, J.; Horstmann, M.; Hennenlotter, J.; Kruck, S.; Stenzl, A. Economic aspects of bladder cancer: What are the benefits and costs? World J. Urol. 2009, 27, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.B.; Howard, L.E.; Foster, M.L.; Klaassen, Z.; Sieluk, J.; De Hoedt, A.M.; Freedland, S.J. Estimated Costs and Long-term Outcomes of Patients With High-Risk Non-Muscle-Invasive Bladder Cancer Treated With Bacillus Calmette-Guerin in the Veterans Affairs Health System. JAMA Netw. Open 2021, 4, e213800. [Google Scholar] [CrossRef]
- Wu, X.R. Urothelial tumorigenesis: A tale of divergent pathways. Nat. Rev. Cancer 2005, 5, 713–725. [Google Scholar] [CrossRef]
- Mo, L.; Zheng, X.; Huang, H.Y.; Shapiro, E.; Lepor, H.; Cordon-Cardo, C.; Sun, T.T.; Wu, X.R. Hyperactivation of Ha-ras oncogene, but not Ink4a/Arf deficiency, triggers bladder tumorigenesis. J. Clin. Investig. 2007, 117, 314–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zi, X.; Simoneau, A.R. Flavokawain A, a novel chalcone from kava extract, induces apoptosis in bladder cancer cells by involvement of Bax protein-dependent and mitochondria-dependent apoptotic pathway and suppresses tumor growth in mice. Cancer Res. 2005, 65, 3479–3486. [Google Scholar] [CrossRef] [Green Version]
- Steiner, G.G. The correlation between cancer incidence and kava consumption. Hawaii Med. J. 2000, 59, 420–422. [Google Scholar]
- Thomsen, M.; Schmidt, M. Health policy versus kava (Piper methysticum): Anxiolytic efficacy may be instrumental in restoring the reputation of a major South Pacific crop. J. Ethnopharmacol. 2021, 268, 113582. [Google Scholar] [CrossRef]
- Tang, Y.; Simoneau, A.R.; Xie, J.; Shahandeh, B.; Zi, X. Effects of the kava chalcone flavokawain A differ in bladder cancer cells with wild-type versus mutant p53. Cancer Prev. Res. 2008, 1, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Xu, X.; Li, X.; Liu, S.; Simoneau, A.R.; He, F.; Wu, X.R.; Zi, X. Kava chalcone, flavokawain A, inhibits urothelial tumorigenesis in the UPII-SV40T transgenic mouse model. Cancer Prev. Res. 2013, 6, 1365–1375. [Google Scholar] [CrossRef] [Green Version]
- Minoli, M.; Kiener, M.; Thalmann, G.N.; Kruithof-de Julio, M.; Seiler, R. Evolution of Urothelial Bladder Cancer in the Context of Molecular Classifications. Int. J. Mol. Sci. 2020, 21, 5670. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yokoyama, N.N.; Blair, C.A.; Li, X.; Avizonis, D.; Wu, X.R.; Uchio, E.; Youssef, R.; McClelland, M.; Pollak, M.; et al. High Sensitivity of an Ha-RAS Transgenic Model of Superficial Bladder Cancer to Metformin Is Associated with 240-Fold Higher Drug Concentration in Urine than Serum. Mol. Cancer Ther. 2016, 15, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matulewicz, R.S.; Rademaker, A.; Meeks, J.J. A simplified nomogram to assess risk of bladder cancer in patients with a new diagnosis of microscopic hematuria. Urol. Oncol. 2020, 38, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Chromecki, T.F.; Bensalah, K.; Remzi, M.; Verhoest, G.; Cha, E.K.; Scherr, D.S.; Novara, G.; Karakiewicz, P.I.; Shariat, S.F. Prognostic factors for upper urinary tract urothelial carcinoma. Nat. Rev. Urol. 2011, 8, 440–447. [Google Scholar] [CrossRef]
- Woldu, S.L.; Bagrodia, A.; Lotan, Y. Guideline of guidelines: Non-muscle-invasive bladder cancer. BJU Int. 2017, 119, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Zenger, K.; Agnolet, S.; Schneider, B.; Kraus, B. Biotransformation of Flavokawains A, B, and C, Chalcones from Kava (Piper methysticum), by Human Liver Microsomes. J. Agric. Food Chem. 2015, 63, 6376–6385. [Google Scholar] [CrossRef]
- Hartmann, A.; Schlake, G.; Zaak, D.; Hungerhuber, E.; Hofstetter, A.; Hofstaedter, F.; Knuechel, R. Occurrence of chromosome 9 and p53 alterations in multifocal dysplasia and carcinoma in situ of human urinary bladder. Cancer Res. 2002, 62, 809–818. [Google Scholar]
- Al-Ahmadie, H.A.; Iyer, G.; Janakiraman, M.; Lin, O.; Heguy, A.; Tickoo, S.K.; Fine, S.W.; Gopalan, A.; Chen, Y.B.; Balar, A.; et al. Somatic mutation of fibroblast growth factor receptor-3 (FGFR3) defines a distinct morphological subtype of high-grade urothelial carcinoma. J. Pathol. 2011, 224, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Abu, N.; Akhtar, M.N.; Yeap, S.K.; Lim, K.L.; Ho, W.Y.; Zulfadli, A.J.; Omar, A.R.; Sulaiman, M.R.; Abdullah, M.P.; Alitheen, N.B. Flavokawain A induces apoptosis in MCF-7 and MDA-MB231 and inhibits the metastatic process in vitro. PLoS ONE 2014, 9, e105244. [Google Scholar] [CrossRef]
- Zhang, Z.T.; Pak, J.; Shapiro, E.; Sun, T.T.; Wu, X.R. Urothelium-specific expression of an oncogene in transgenic mice induced the formation of carcinoma in situ and invasive transitional cell carcinoma. Cancer Res. 1999, 59, 3512–3517. [Google Scholar]
- Jandial, D.D.; Krill, L.S.; Chen, L.; Wu, C.; Ke, Y.; Xie, J.; Hoang, B.H.; Zi, X. Induction of G2M Arrest by Flavokawain A, a Kava Chalcone, Increases the Responsiveness of HER2-Overexpressing Breast Cancer Cells to Herceptin. Molecules 2017, 22, 462. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, E.C.; Drezner, N.; Li, X.; Mishra-Kalyani, P.S.; Liu, Y.; Zhao, H.; Bi, Y.; Liu, J.; Rahman, A.; Wearne, E.; et al. FDA Approval Summary: Sotorasib for KRAS G12C-Mutated Metastatic NSCLC. Clin. Cancer Res. 2021; Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Enokida, H.; Yoshino, H.; Miyamoto, K.; Yonemori, M.; Sakaguchi, T.; Osako, Y.; Nakagawa, M. HRAS as a potential therapeutic target of salirasib RAS inhibitor in bladder cancer. Int. J. Oncol. 2018, 53, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Maas, M.; Stühler, V.; Walz, S.; Stenzl, A.; Bedke, J. Enfortumab vedotin—Next game-changer in urothelial cancer. Expert Opin. Biol. Ther. 2021, 21, 801–809. [Google Scholar] [CrossRef]
- Mohammed, A.; Miller, M.S.; Lubet, R.A.; Suen, C.S.; Sei, S.; Shoemaker, R.H.; Juliana, M.M.; Moeinpour, F.L.; Grubbs, C.J. Combination of Erlotinib and Naproxen Employing Pulsatile or Intermittent Dosing Profoundly Inhibits Urinary Bladder Cancers. Cancer Prev. Res. 2020, 13, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.; Maurer, A.; Wirtz, J.; Bleilevens, A.; Waldmann, T.; Wenz, M.; Eyll, M.; Geelvink, M.; Gereitzig, M.; Rüchel, N.; et al. EGFR activity addiction facilitates anti-ERBB based combination treatment of squamous bladder cancer. Oncogene 2020, 39, 6856–6870. [Google Scholar] [CrossRef]
- Li, X.; Xu, X.; Ji, T.; Liu, Z.; Gu, M.; Hoang, B.H.; Zi, X. Dietary feeding of Flavokawain A, a Kava chalcone, exhibits a satisfactory safety profile and its association with enhancement of phase II enzymes in mice. Toxicol. Rep. 2014, 1, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yokoyama, N.N.; Zhang, S.; Ding, L.; Liu, H.M.; Lilly, M.B.; Mercola, D.; Zi, X. Flavokawain A induces deNEDDylation and Skp2 degradation leading to inhibition of tumorigenesis and cancer progression in the TRAMP transgenic mouse model. Oncotarget 2015, 6, 41809–41824. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Song, L.; Xie, J.; Simoneau, A.R.; Uchio, E.; Zi, X. Chemoprevention of Urothelial Cell Carcinoma Tumorigenesis by Dietary Flavokawain A in UPII-Mutant Ha-ras Transgenic Mice. Pharmaceutics 2022, 14, 496. https://doi.org/10.3390/pharmaceutics14030496
Liu Z, Song L, Xie J, Simoneau AR, Uchio E, Zi X. Chemoprevention of Urothelial Cell Carcinoma Tumorigenesis by Dietary Flavokawain A in UPII-Mutant Ha-ras Transgenic Mice. Pharmaceutics. 2022; 14(3):496. https://doi.org/10.3390/pharmaceutics14030496
Chicago/Turabian StyleLiu, Zhongbo, Liankun Song, Jun Xie, Anne R. Simoneau, Edward Uchio, and Xiaolin Zi. 2022. "Chemoprevention of Urothelial Cell Carcinoma Tumorigenesis by Dietary Flavokawain A in UPII-Mutant Ha-ras Transgenic Mice" Pharmaceutics 14, no. 3: 496. https://doi.org/10.3390/pharmaceutics14030496
APA StyleLiu, Z., Song, L., Xie, J., Simoneau, A. R., Uchio, E., & Zi, X. (2022). Chemoprevention of Urothelial Cell Carcinoma Tumorigenesis by Dietary Flavokawain A in UPII-Mutant Ha-ras Transgenic Mice. Pharmaceutics, 14(3), 496. https://doi.org/10.3390/pharmaceutics14030496