
l-Arginine Induces White Adipose Tissue Browning—A New Pharmaceutical Alternative to Cold


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Study
2.2. Biochemistry/ELISA Assays
2.3. Isolation of Mature Adipocytes
2.4. NO Production
2.5. H&E Staining
2.6. Immunohistochemistry of UCP1
2.7. RT-PCR/Gene Expression Analysis
2.8. Mitochondrial DNA Copy Number (MCN)
2.9. Western Blot/Protein Expression Analysis
2.10. Statistical Analyses
3. Results
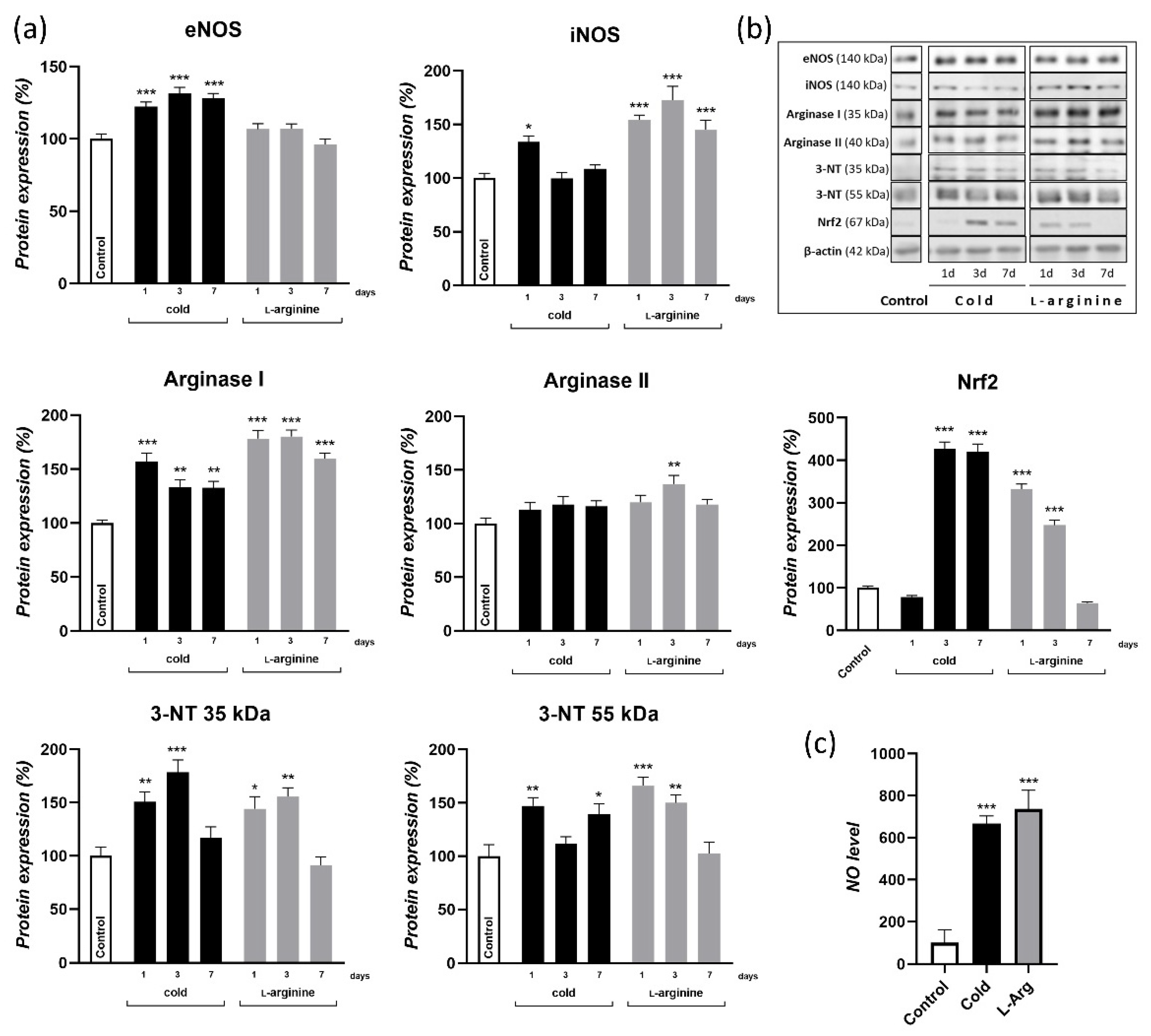
3.1. Cold Exposure-Induced Browning Is Associated with Endogenous NO Synthesis; l-Arginine Increases NO Synthesis in rpWAT to a Similar Extent
3.2. Effects of l-Arginine Treatment on Circulatory Parameters and Adipokines Levels
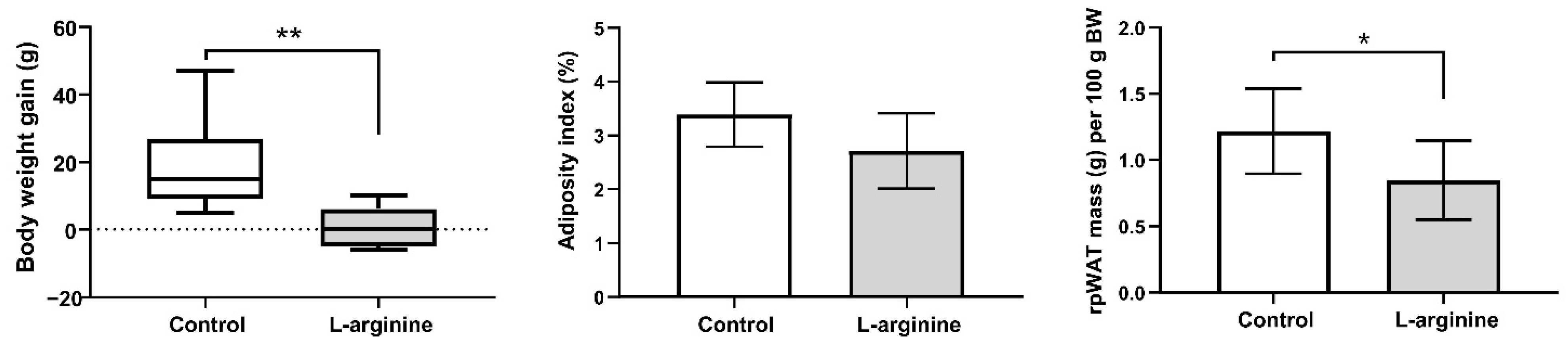
3.3. Slimming Effects of l-Arginine Are Associated with Diminution of rpWAT Depot Mass
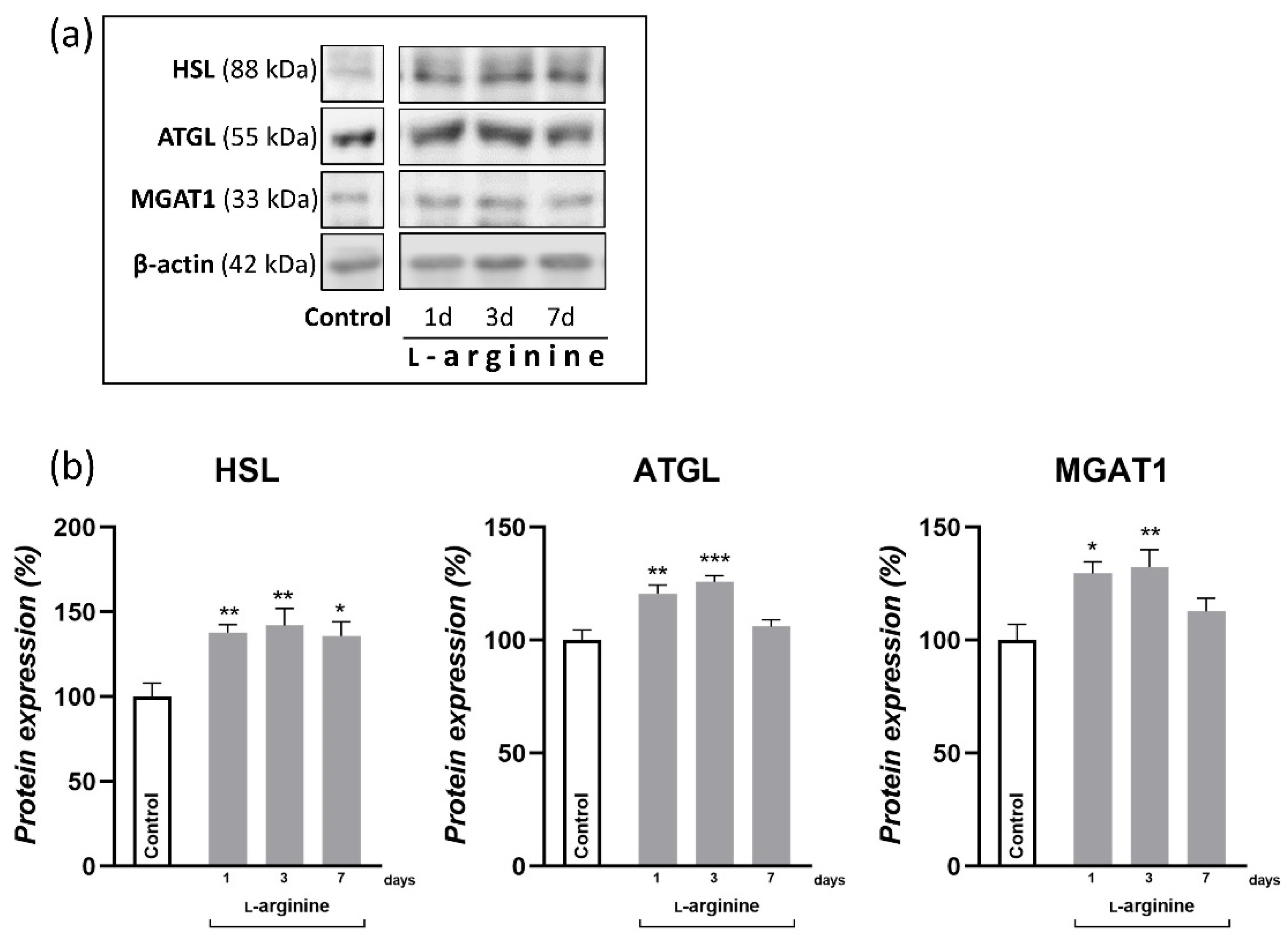
3.4. Net Lipid Mobilization Effect of l-Arginine Is Associated with Increased Lipolysis
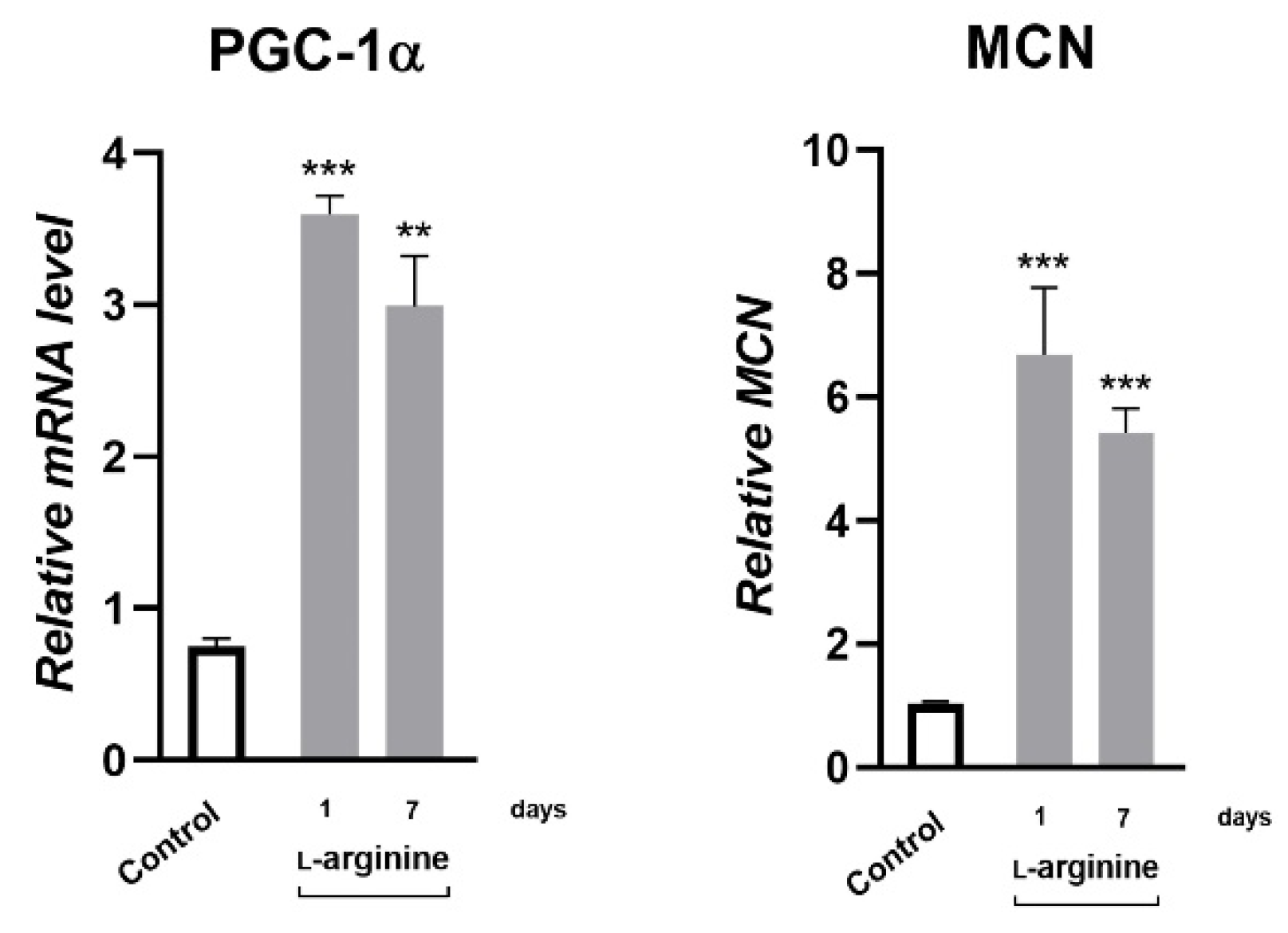
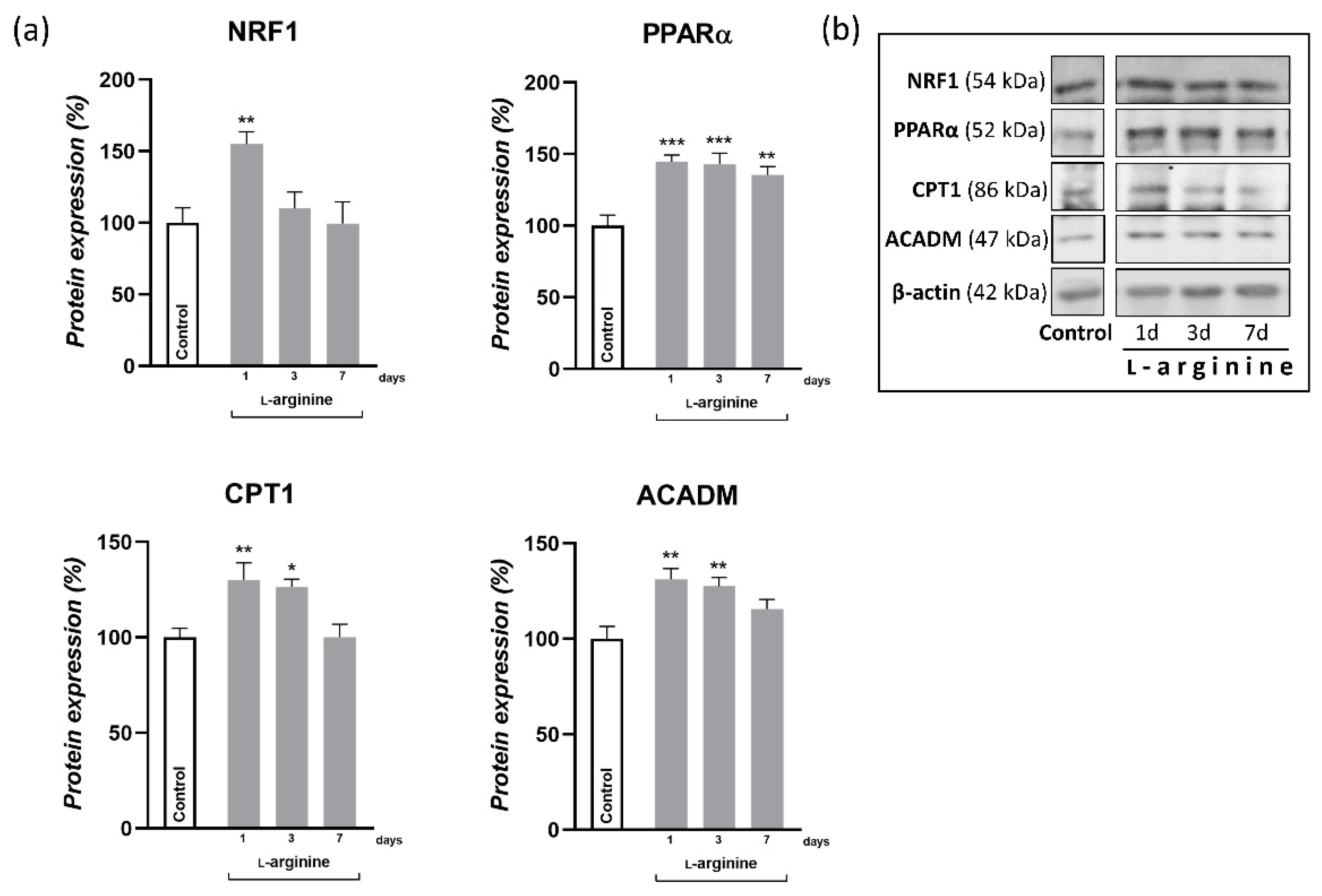
3.5. l-Arginine Elicits an Increase in Expression of PGC-1α, NRF1, PPARα, and Their Downstream Mitochondrial Targets in rpWAT
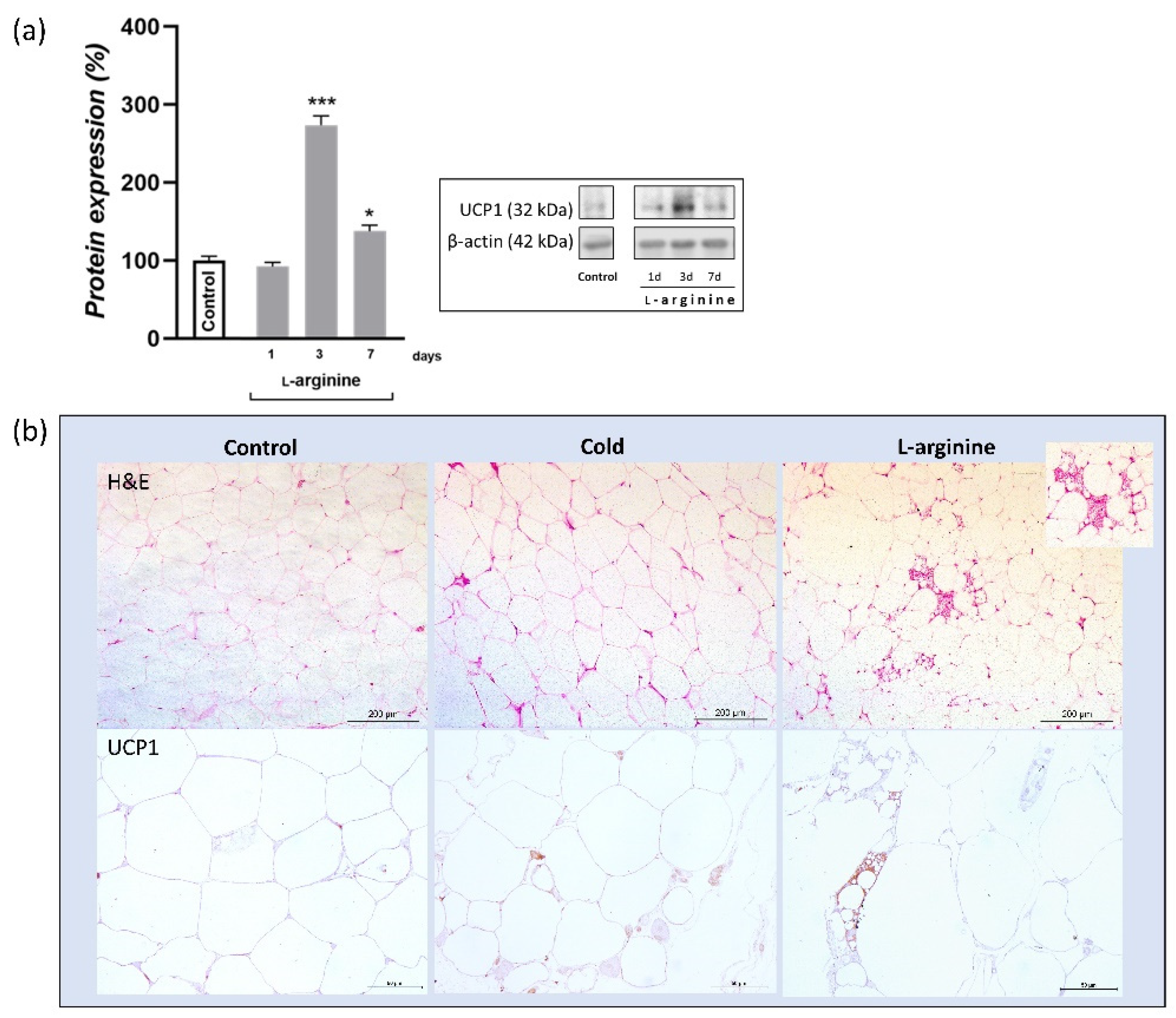
3.6. UCP1 Protein Expression Level Increases upon l-Arginine Treatment and Over-Exceeds Cold-Induced Effects
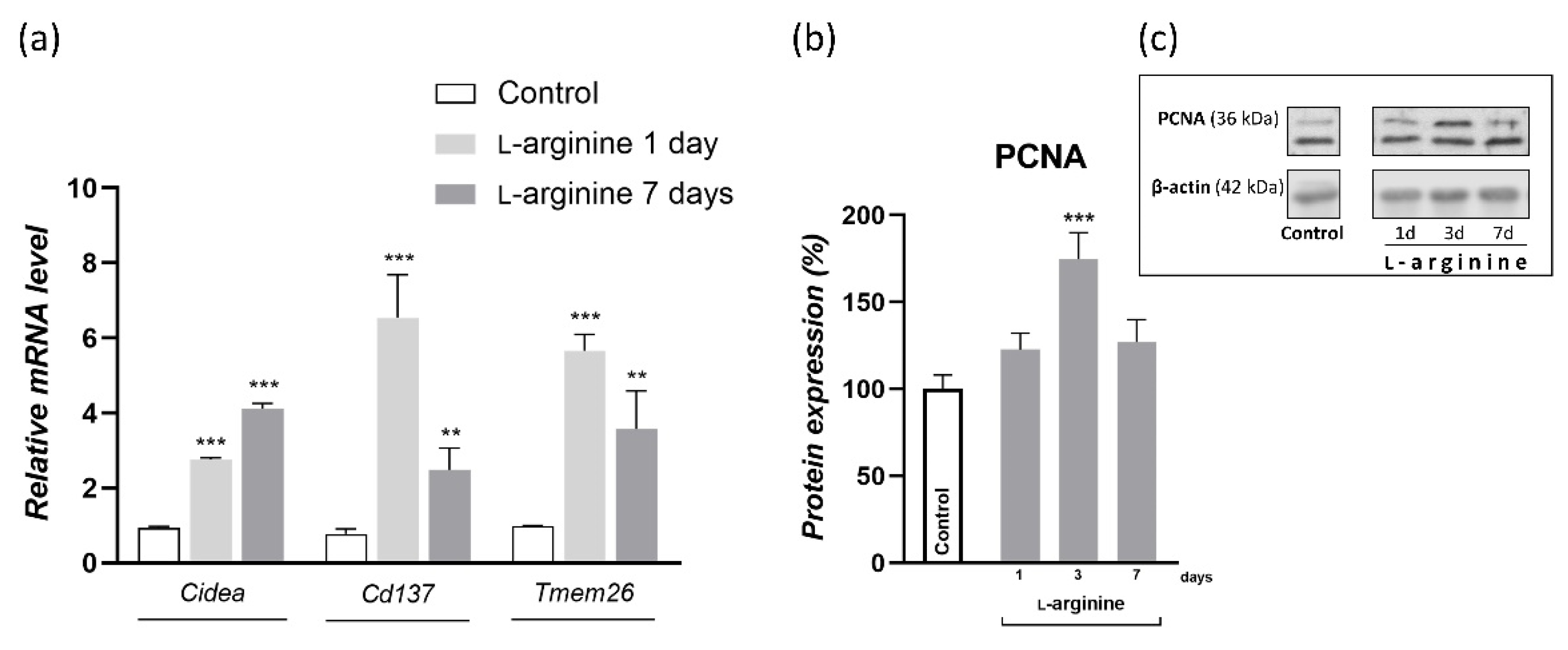
3.7. l-Arginine Elicits the Expression of Genetic Markers of Beige Adipocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sakers, A.; De Siqueira, M.K.; Seale, P.; Villanueva, C.J. Adipose-tissue plasticity in health and disease. Cell 2022, 185, 419–446. [Google Scholar] [CrossRef]
- Jankovic, A.; Golic, I.; Markelic, M.; Stancic, A.; Otasevic, V.; Buzadzic, B.; Korac, A.; Korac, B. Two key temporally distinguishable molecular and cellular components of white adipose tissue browning during cold acclimation. J. Physiol. 2015, 593, 3267–3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedergaard, J.; Golozoubova, V.; Matthias, A.; Asadi, A.; Jacobsson, A.; Cannon, B. UCP1: The only protein able to mediate adaptive non-shivering thermogenesis and metabolic inefficiency. Biochim. Biophys. Acta Bioenerg. 2001, 1504, 82–106. [Google Scholar] [CrossRef] [Green Version]
- Ricquier, D. Uncoupling protein 1 of brown adipocytes, the only uncoupler: A historical perspective. Front. Endocrinol. 2011, 2, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flachs, P.; Rossmeisl, M.; Kuda, O.; Kopecky, J. Stimulation of mitochondrial oxidative capacity in white fat independent of UCP1: A key to lean phenotype. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 986–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–206. [Google Scholar] [CrossRef]
- Yoneshiro, T.; Wang, Q.; Tajima, K.; Matsushita, M.; Maki, H.; Igarashi, K.; Dai, Z.; White, P.J.; McGarrah, R.W.; Ilkayeva, O.R.; et al. BCAA catabolism in brown fat controls energy homeostasis through SLC25A44. Nature 2019, 572, 614–619. [Google Scholar] [CrossRef]
- Jankovic, A.; Otasevic, V.; Stancic, A.; Buzadzic, B.; Korac, A.; Korac, B. Physiological regulation and metabolic role of browning in white adipose tissue. Horm. Mol. Biol. Clin. Investig. 2017, 31, 20170034. [Google Scholar] [CrossRef]
- Petrović, V.; Buzadžić, B.; Korać, A.; Vasilijević, A.; Janković, A.; Korać, B. NO modulates the molecular basis of rat interscapular brown adipose tissue thermogenesis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 147–159. [Google Scholar] [CrossRef]
- Kapur, S.; Picard, F.; Perreault, M.; Deshaies, Y.; Marette, A. Nitric oxide: A new player in the modulation of energy metabolism. Int. J. Obes. 2000, 24, S36–S40. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.K.; Kuroshima, A. Nitric oxide and thermogenic function of brown adipose tissue in rats. Jpn. J. Physiol. 2000, 50, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi-Utsumi, K.; Gao, B.; Ohinata, H.; Hashimoto, M.; Yamamoto, N.; Kuroshima, A. Enhanced gene expression of endothelial nitric oxide synthase in brown adipose tissue during cold exposure. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R623–R626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Satterfield, M.C.; Bazer, F.W.; Wu, G. Regulation of brown adipose tissue development and white fat reduction by L-arginine. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Ashmore, T.; Kotwica, A.O.; Murfitt, S.A.; Fernandez, B.O.; Feelisch, M.; Murray, A.J.; Griffin, J.L. Inorganic nitrate promotes the browning of white adipose tissue through the nitrate-nitrite-nitric oxide pathway. Diabetes 2015, 64, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Peleli, M.; Ferreira, D.M.S.; Tarnawski, L.; McCann Haworth, S.; Xuechen, L.; Zhuge, Z.; Newton, P.T.; Massart, J.; Chagin, A.S.; Olofsson, P.S.; et al. Dietary nitrate attenuates high-fat diet-induced obesity via mechanisms involving higher adipocyte respiration and alterations in inflammatory status. Redox Biol. 2020, 28, 101387. [Google Scholar] [CrossRef]
- Van Faassen, E.E.; Bahrami, S.; Feelisch, M.; Hogg, N.; Kelm, M.; Kim-Shapiro, D.B.; Kozlov, A.V.; Li, H.; Lundberg, J.O.; Mason, R.; et al. Nitrite as regulator of hypoxic signaling in mammalian physiology. Med. Res. Rev. 2009, 29, 683–741. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, A.J.; Fukuto, J.M.; Ignarro, L.J. Formation of free nitric oxide from L-arginine by nitric oxide synthase: Direct enhancement of generation by superoxide dismutase. Proc. Natl. Acad. Sci. USA 1994, 91, 10992–10996. [Google Scholar] [CrossRef] [Green Version]
- Jobgen, W.; Meininger, C.J.; Jobgen, S.C.; Li, P.; Lee, M.J.; Smith, S.B.; Spencer, T.E.; Fried, S.K.; Wu, G. Dietary L-arginine supplementation reduces white fat gain and enhances skeletal muscle and brown fat masses in diet-induced obese rats. J. Nutr. 2009, 139, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Jankovic, A.; Korac, A.; Buzadzic, B.; Stancic, A.; Otasevic, V.; Ferdinandy, P.; Daiber, A.; Korac, B. Targeting the NO/superoxide ratio in adipose tissue: Relevance to obesity and diabetes management. Br. J. Pharmacol. 2017, 174, 1570–1590. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.J.; Haynes, T.E.; Kohli, R.; Hu, J.; Shi, W.; Spencer, T.E.; Carroll, R.J.; Meininger, C.J.; Wu, G. Dietary L-arginine supplementation reduces fat mass in Zucker diabetic fatty rats. J. Nutr. 2005, 135, 714–721. [Google Scholar] [CrossRef] [Green Version]
- Monti, L.D.; Setola, E.; Lucotti, P.C.G.; Marrocco-Trischitta, M.M.; Comola, M.; Galluccio, E.; Poggi, A.; Mammì, S.; Catapano, A.L.; Comi, G.; et al. Effect of a long-term oral l-arginine supplementation on glucose metabolism: A randomized, double-blind, placebo-controlled trial. Diabetes Obes. Metab. 2012, 14, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Lucotti, P.; Setola, E.; Monti, L.D.; Galluccio, E.; Costa, S.; Sandoli, E.P.; Fermo, I.; Rabaiotti, G.; Gatti, R.; Piatti, P.M. Beneficial effects of a long-term oral L-arginine treatment added to a hypocaloric diet and exercise training program in obese, insulin-resistant type 2 diabetic patients. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E906–E912. [Google Scholar] [CrossRef] [PubMed]
- McKnight, J.R.; Satterfield, M.C.; Jobgen, W.S.; Smith, S.B.; Spencer, T.E.; Meininger, C.J.; McNeal, C.J.; Wu, G. Beneficial effects of L-arginine on reducing obesity: Potential mechanisms and important implications for human health. Amino Acids 2010, 39, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Nakamura, Y.; Yamaguchi, M.; Mori, T.; Uchida, M.; Fujita, S. Amino acid mixture enriched with arginine, alanine, and phenylalanine stimulates fat metabolism during exercise. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 46–54. [Google Scholar] [CrossRef]
- Sasai, H.; Ueda, K.; Tsujimoto, T.; Kobayashi, H.; Sanbongi, C.; Ikegami, S.; Nakata, Y. Dose-ranging pilot randomized trial of amino acid mixture combined with physical activity promotion for reducing abdominal fat in overweight adults. Diabetes Metab. Syndr. Obes. Targets Ther. 2017, 10, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Ueda, K.; Sanbongi, C.; Takai, S.; Ikegami, S.; Fujita, S. Combination of aerobic exercise and an arginine, alanine, and phenylalanine mixture increases fat mobilization and ketone body synthesis. Biosci. Biotechnol. Biochem. 2017, 81, 1417–1424. [Google Scholar] [CrossRef] [Green Version]
- Ueda, K.; Sasai, H.; Tsujimoto, T.; Sanbongi, C.; Ikegami, S.; Kobayashi, H.; Shioya, N.; Suzuki, S.; Nakata, Y. Randomized trial of amino acid mixture combined with physical activity promotion for abdominal fat reduction in overweight adults. Diabetes Metab. Syndr. Obes. Targets Ther. 2018, 11, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Jobgen, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.J.; Wu, G. Regulatory role for the arginine-nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem. 2006, 17, 571–588. [Google Scholar] [CrossRef]
- Hurt, R.T.; Ebbert, J.O.; Schroeder, D.R.; Croghan, I.T.; Bauer, B.A.; McClave, S.A.; Miles, J.M.; McClain, C.J. L-arginine for the treatment of centrally obese subjects: A pilot study. J. Diet. Suppl. 2014, 11, 40–52. [Google Scholar] [CrossRef]
- Lončar, D. Convertible adipose tissue in mice. Cell Tissue Res. 1991, 266, 149–161. [Google Scholar] [CrossRef]
- Giordano, A.; Morroni, M.; Carle, F.; Gesuita, R.; Marchesi, G.F.; Cinti, S. Sensory nerves affect the recruitment and differentiation of rat periovarian brown adipocytes during cold acclimation. J. Cell Sci. 1998, 111, 2587–2594. [Google Scholar] [CrossRef] [PubMed]
- Himms-Hagen, J.; Melnyk, A.; Zingaretti, M.C.; Ceresi, E.; Barbatelli, G.; Cinti, S. Multilocular fat cells in WAT of CL-316243-treated rats derive directly from white adipocytes. Am. J. Physiol. Cell Physiol. 2000, 279, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Frontini, A.; Cinti, S. Distribution and Development of Brown Adipocytes in the Murine and Human Adipose Organ. Cell Metab. 2010, 11, 253–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldén, T.B.; Hansen, I.R.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Recruited vs. nonrecruited molecular signatures of brown, “brite,” and white adipose tissues. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E19–E31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Lans, A.A.J.J.; Hoeks, J.; Brans, B.; Vijgen, G.H.E.J.; Visser, M.G.W.; Vosselman, M.J.; Hansen, J.; Jörgensen, J.A.; Wu, J.; Mottaghy, F.M.; et al. Cold acclimation recruits human brown fat and increases nonshivering thermogenesis. J. Clin. Invest. 2013, 123, 3395–3403. [Google Scholar] [CrossRef]
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.A.F.L.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J.J. Cold-Activated Brown Adipose Tissue in Healthy Men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.W.; Fernstrom, J.D.; Thompson, J.; Morris, S.M.; Kuller, L.H. Biochemical responses of healthy subjects during dietary supplementation with L-arginine. J. Nutr. Biochem. 2004, 15, 534–539. [Google Scholar] [CrossRef]
- Petrović, V.; Buzadžić, B.; Korać, A.; Korać, B. Antioxidative defense and mitochondrial thermogenic response in brown adipose tissue. Genes Nutr. 2010, 5, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Stancic, A.; Buzadzic, B.; Korac, A.; Otasevic, V.; Jankovic, A.; Vucetic, M.; Markelic, M.; Velickovic, K.; Golic, I.; Korac, B. Regulatory role of PGC-1α/PPAR signaling in skeletal muscle metabolic recruitment during cold acclimation. J. Exp. Biol. 2013, 216, 4233–4241. [Google Scholar] [CrossRef] [Green Version]
- Zeinali Khosroshahi, M.; Asbaghi, O.; Moradi, S.; Rezaei kelishadi, M.; Kaviani, M.; Mardani, M.; Jalili, C. The effects of supplementation with L-arginine on anthropometric indices and body composition in overweight or obese subjects: A systematic review and meta-analysis. J. Funct. Foods 2020, 71, 104022. [Google Scholar] [CrossRef]
- Mousavi, S.M.; Milajerdi, A.; Fatahi, S.; Rahmani, J.; Zarezadeh, M.; Ghaedi, E.; Varkaneh, H.K. The effect of L-arginine supplementation on obesity-related indices: A systematic review and meta-analysis of randomized clinical trials. Int. J. Vitam. Nutr. Res. 2021, 91, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Daneghian, S.; Ghaffari, A.; Ostadrahimi, A.; Safaeiyan, A.; Estakhri, R.; Gargari, B.P. The effect of hypocaloric diet enriched in legumes with or without L- Arginine and selenium on anthropometric measures in central obese women. J. Res. Med. Sci. 2010, 15, 331–343. [Google Scholar] [PubMed]
- Hadi, A.; Arab, A.; Moradi, S.; Pantovic, A.; Clark, C.C.T.; Ghaedi, E. The effect of l-arginine supplementation on lipid profile: A systematic review and meta-analysis of randomised controlled trials. Br. J. Nutr. 2019, 122, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Dashtabi, A.; Mazloom, Z.; Fararouei, M.; Hejazi, N. Oral L-Arginine Administration Improves Anthropometric and Biochemical Indices Associated With Cardiovascular Diseases in Obese Patients: A Randomized, Single Blind Placebo Controlled Clinical Trial. Res. Cardiovasc. Med. 2015, 5, e29419. [Google Scholar] [CrossRef] [Green Version]
- Míguez, I.; Mariño, G.; Rodríguez, B.; Taboada, C. Effects of dietary L-arginine supplementation on serum lipids and intestinal enzyme activities in diabetic rats. J. Physiol. Biochem. 2004, 60, 31–37. [Google Scholar] [CrossRef]
- Miczke, A.; Suliburska, J.; Pupek-Musialik, D.; Ostrowska, L.; Jabłecka, A.; Krejpcio, Z.; Skrypnik, D.; Bogdański, P. Effect of L-arginine supplementation on insulin resistance and serum adiponectin concentration in rats with fat diet. Int. J. Clin. Exp. Med. 2015, 8, 10358–10366. [Google Scholar]
- Tan, B.; Li, X.; Yin, Y.; Wu, Z.; Liu, C.; Tekwe, C.D.; Wu, G. Regulatory roles for L-arginine in reducing white adipose tissue. Front. Biosci. 2012, 17, 2237–2246. [Google Scholar] [CrossRef] [Green Version]
- Vucetic, M.; Otasevic, V.; Korac, A.; Stancic, A.; Jankovic, A.; Markelic, M.; Golic, I.; Velickovic, K.; Buzadzic, B.; Korac, B. Interscapular brown adipose tissue metabolic reprogramming during cold acclimation: Interplay of HIF-1α and AMPKα. Biochim. Biophys. Acta Gen. Subj. 2011, 1810, 1252–1261. [Google Scholar] [CrossRef]
- Stancic, A.; Filipovic, M.; Ivanovic-Burmazovic, I.; Masovic, S.; Jankovic, A.; Otasevic, V.; Korac, A.; Buzadzic, B.; Korac, B. Early energy metabolism-related molecular events in skeletal muscle of diabetic rats: The effects of L-arginine and SOD mimic. Chem. Biol. Interact. 2017, 272, 188–196. [Google Scholar] [CrossRef]
- Puigserver, P.; Wu, Z.; Park, C.W.; Graves, R.; Wright, M.; Spiegelman, B.M. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 1998, 92, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Puigserver, P.; Spiegelman, B.M. Peroxisome proliferator-activated receptor-γ coactivator 1α (PGC-1α): Transcriptional coactivator and metabolic regulator. Endocr. Rev. 2003, 24, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | l-Arginine | |
|---|---|---|
| Glucose (mmol L−1) | 5.8 ± 0.5 | 6.0 ± 0.4 |
| Total cholesterol (mmol L−1) | 1.63 ± 0.2 | 1.9 ± 0.2 * |
| LDL-cholesterol (mmol L−1) | 0.51 ± 0.04 | 0.61 ± 0.06 * |
| HDL-cholesterol (mmol L−1) | 0.7 ± 0.1 | 0.9 ± 0.1 * |
| Triglyceride (mmol L−1) | 0.7 ± 0.2 | 0.8 ± 0.1 |
| LDL-cholesterol/HDL-cholesterol | 0.7 ± 0.07 | 0.7 ± 0.02 |
| Total cholesterol/HDL-cholesterol | 2.4 ± 0.2 | 2.3 ± 0.1 |
| TG/HDL-cholesterol | 0.97 ± 0.2 | 0.98 ± 0.03 |
| Leptin (pg mL−1) | 985.7 ± 90.0 | 664.8 ± 56.4 *** |
| Adiponectin (ng mL−1) | 93.9 ± 1.9 | 88.9 ± 8.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalezic, A.; Korac, A.; Korac, B.; Jankovic, A. l-Arginine Induces White Adipose Tissue Browning—A New Pharmaceutical Alternative to Cold. Pharmaceutics 2022, 14, 1368. https://doi.org/10.3390/pharmaceutics14071368
Kalezic A, Korac A, Korac B, Jankovic A. l-Arginine Induces White Adipose Tissue Browning—A New Pharmaceutical Alternative to Cold. Pharmaceutics. 2022; 14(7):1368. https://doi.org/10.3390/pharmaceutics14071368
Chicago/Turabian StyleKalezic, Andjelika, Aleksandra Korac, Bato Korac, and Aleksandra Jankovic. 2022. "l-Arginine Induces White Adipose Tissue Browning—A New Pharmaceutical Alternative to Cold" Pharmaceutics 14, no. 7: 1368. https://doi.org/10.3390/pharmaceutics14071368
APA StyleKalezic, A., Korac, A., Korac, B., & Jankovic, A. (2022). l-Arginine Induces White Adipose Tissue Browning—A New Pharmaceutical Alternative to Cold. Pharmaceutics, 14(7), 1368. https://doi.org/10.3390/pharmaceutics14071368