
Trans-Anethole Alleviates Trimethyltin Chloride-Induced Impairments in Long-Term Potentiation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Preparation of Hippocampal Slices
2.3. Electrophysiological Recordings
2.4. Drugs
2.5. Data Analysis
3. Results
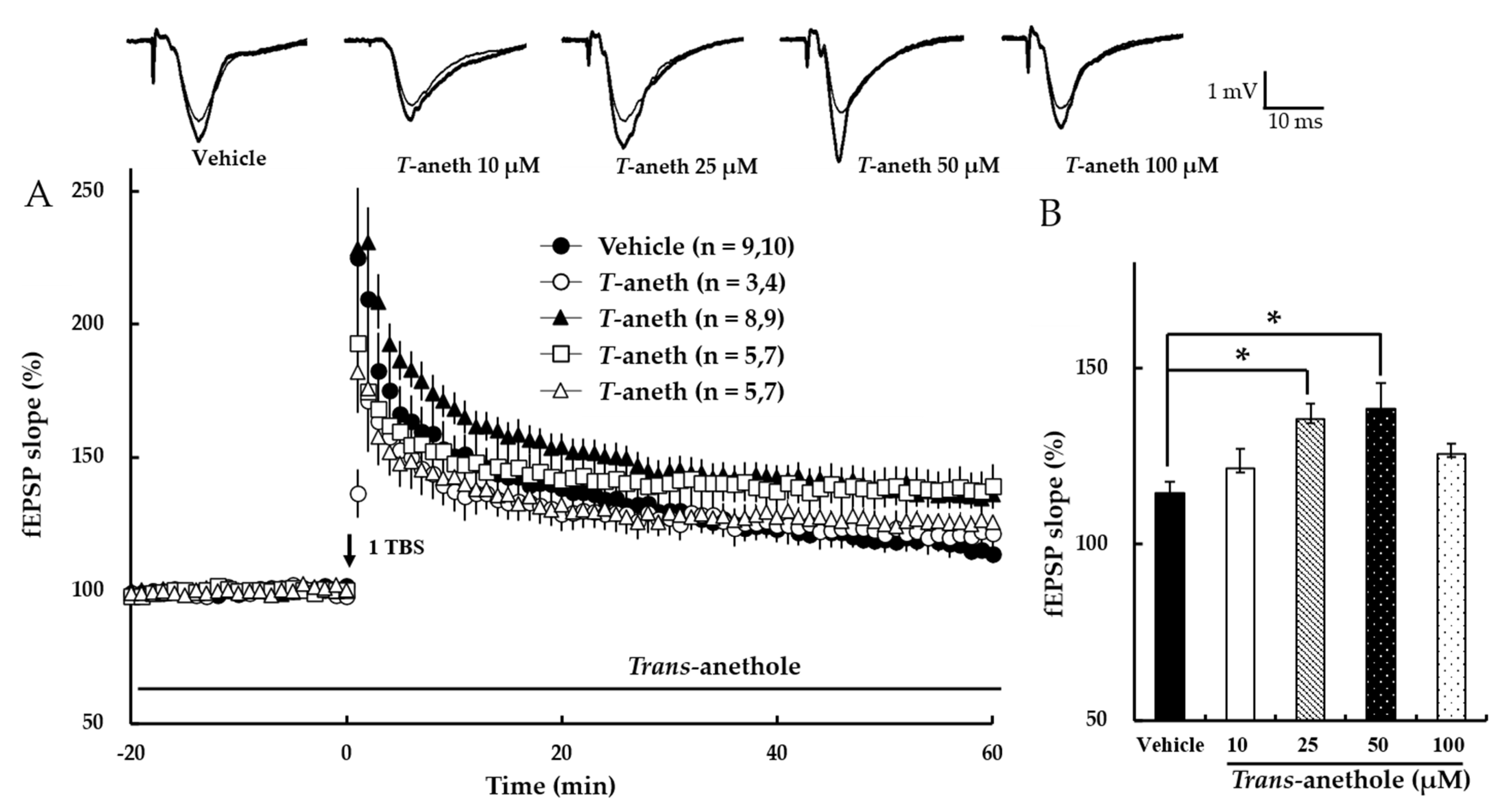
3.1. Trans-Anethole Enhances NMDA Receptor-Dependent LTP
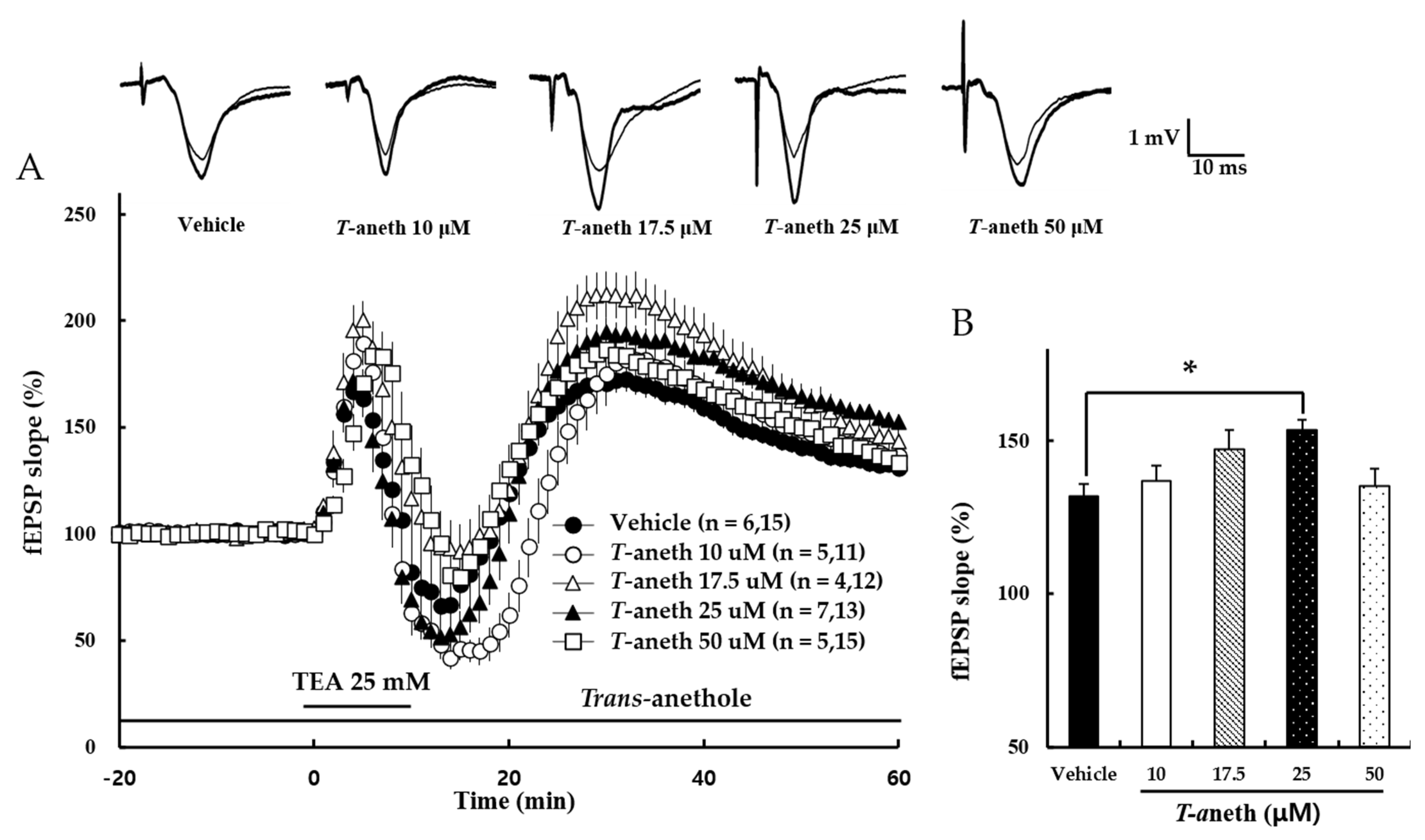
3.2. Trans-Anethole Enhances NMDA Receptor-Independent LTP
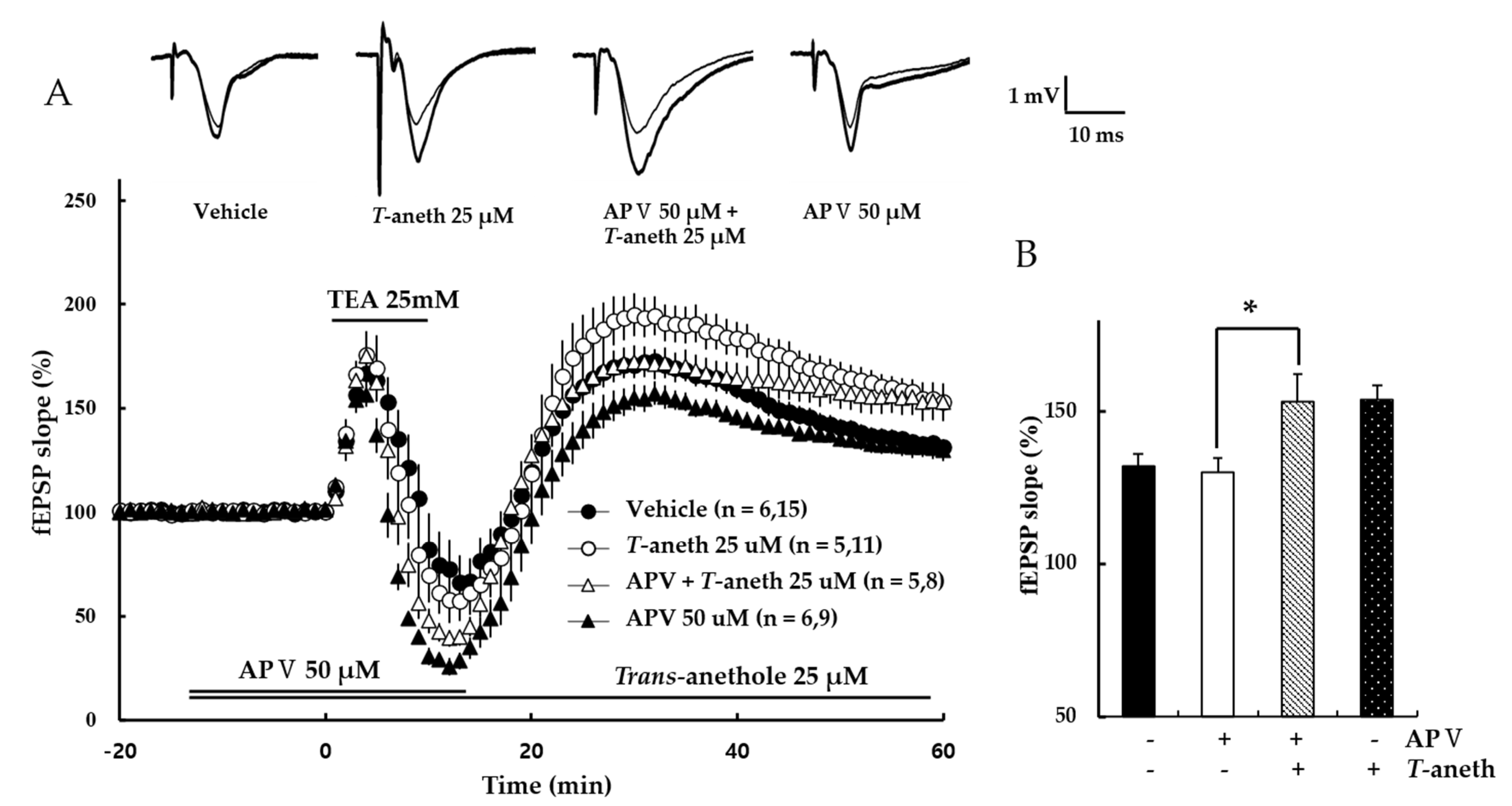
3.3. NMDA Receptor Antagonist Treatment Does Not Alter TEA-Induced LTP Induction
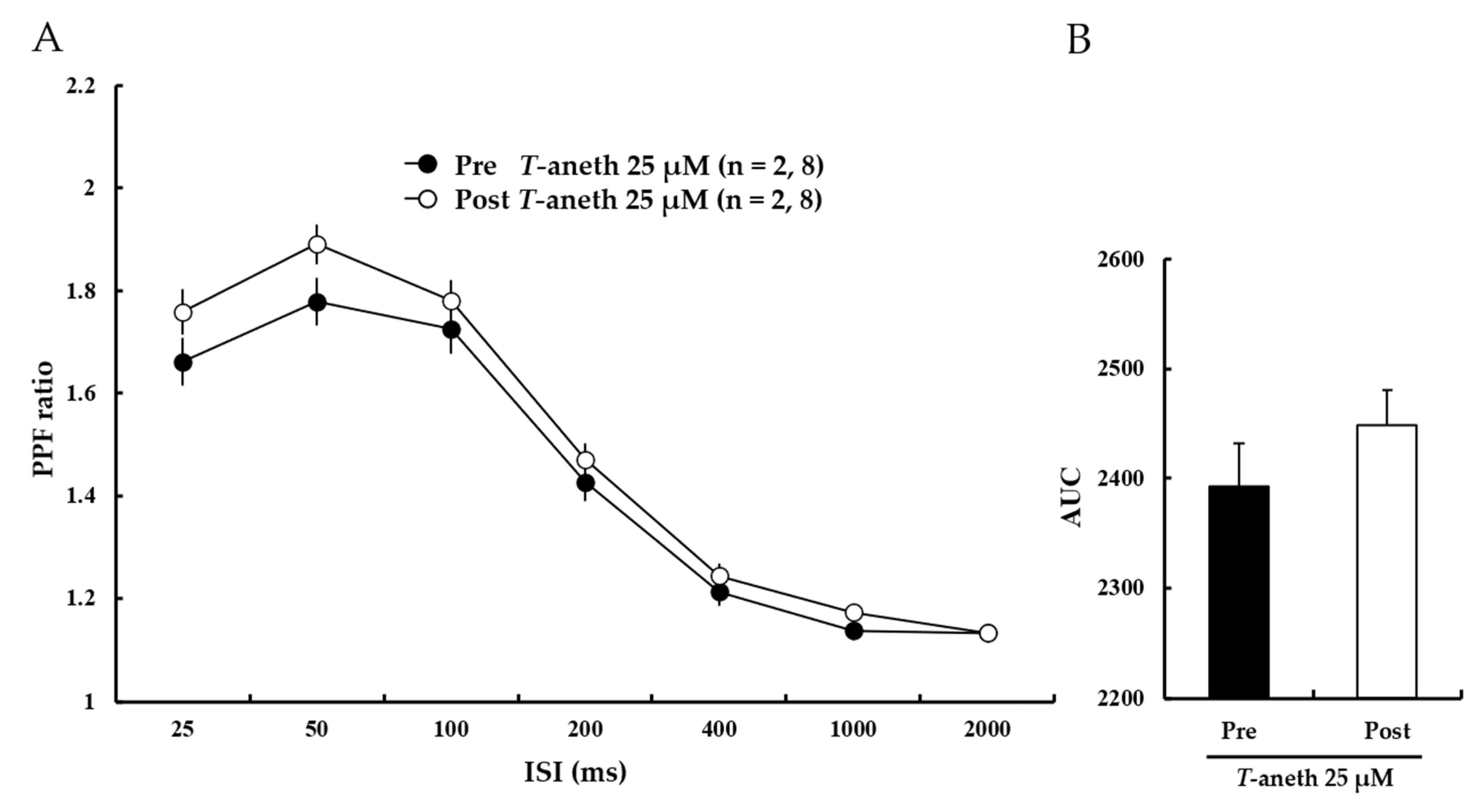
3.4. Trans-Anethole Does Not Affect the Paired-Pulse Facilitation Ratio
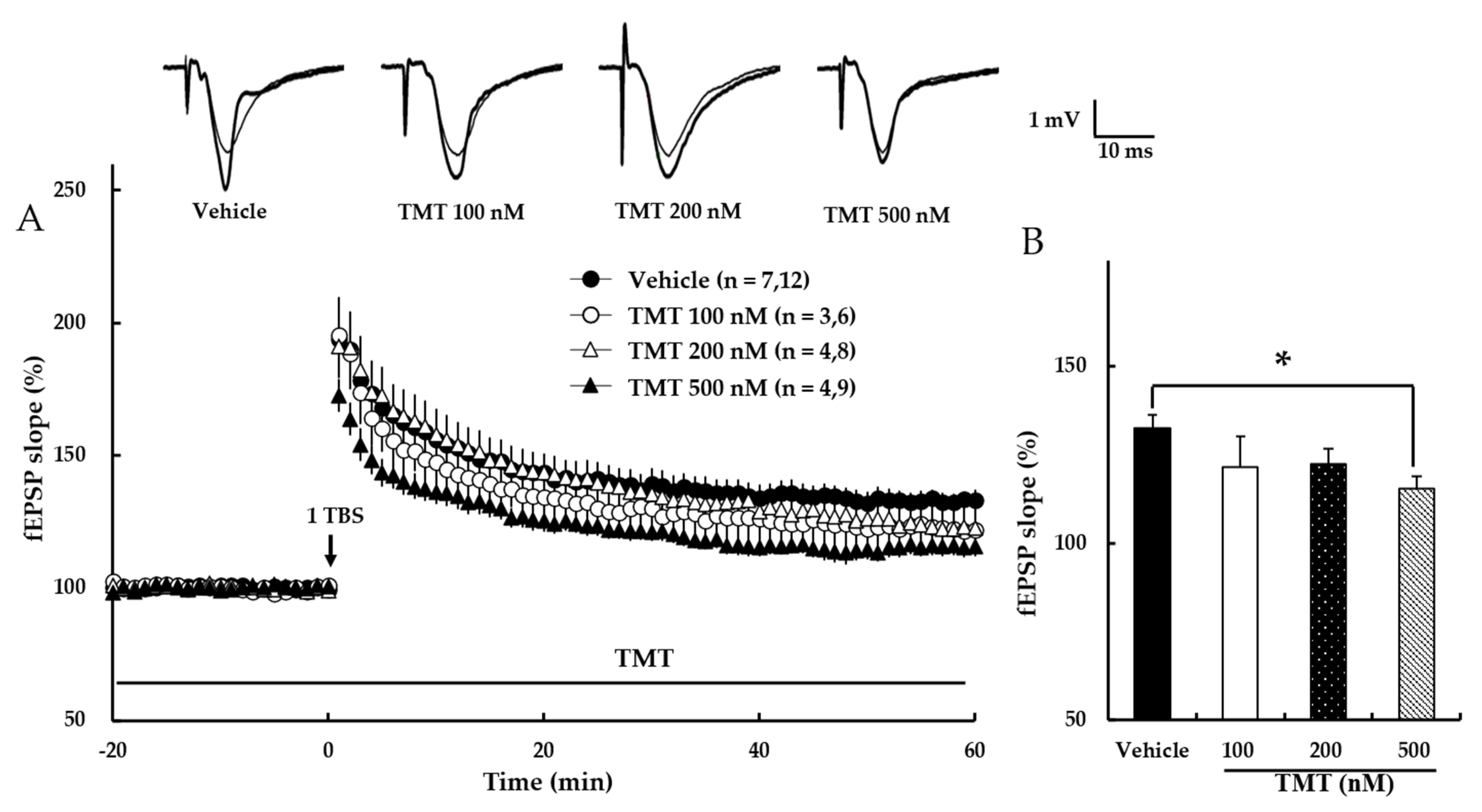
3.5. Trimethyltin Chloride (TMT) Dose-Dependently Impairs NMDA Receptor-Dependent LTP
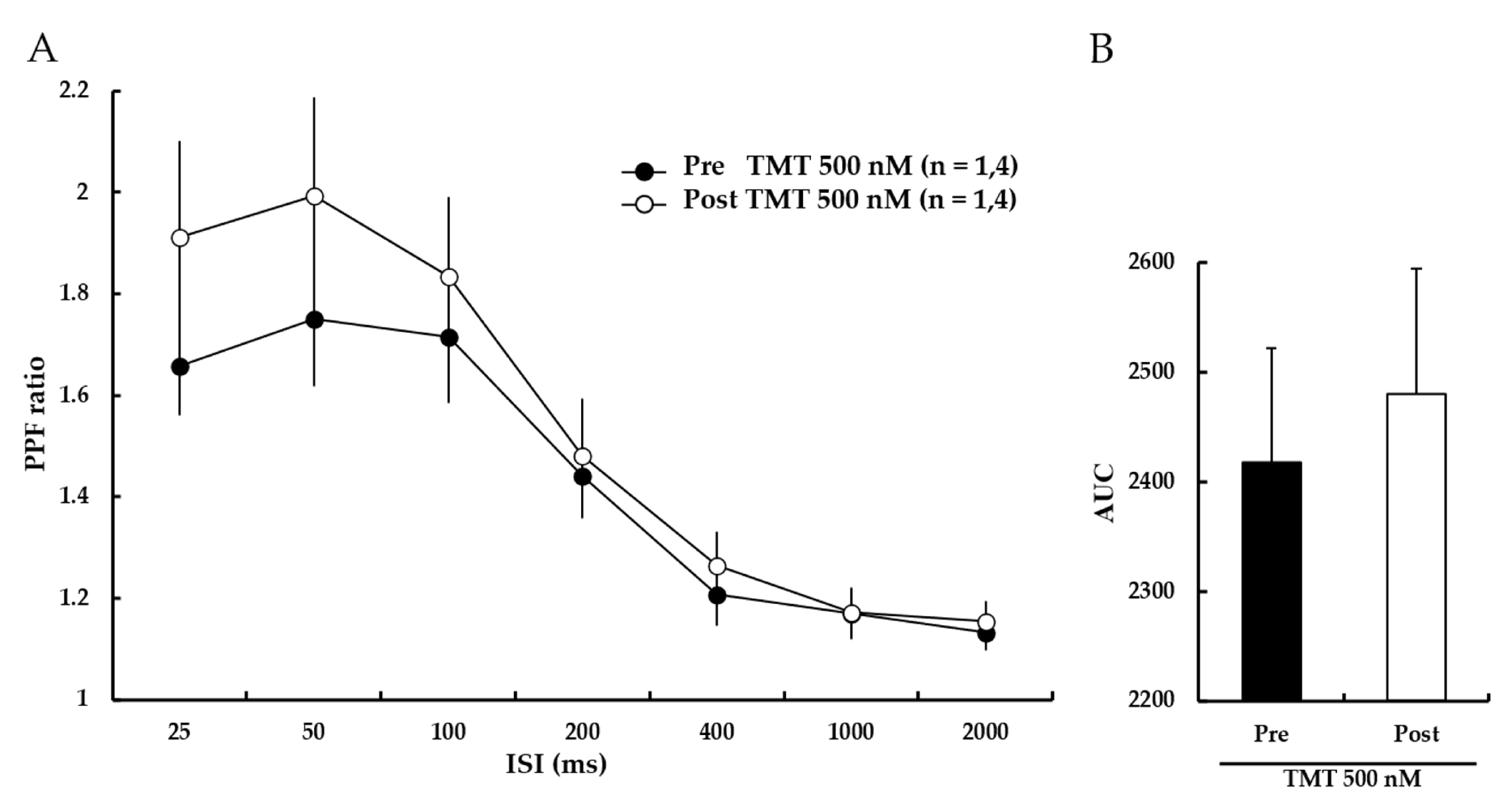
3.6. TMT Does Not Affect the PPF Ratio
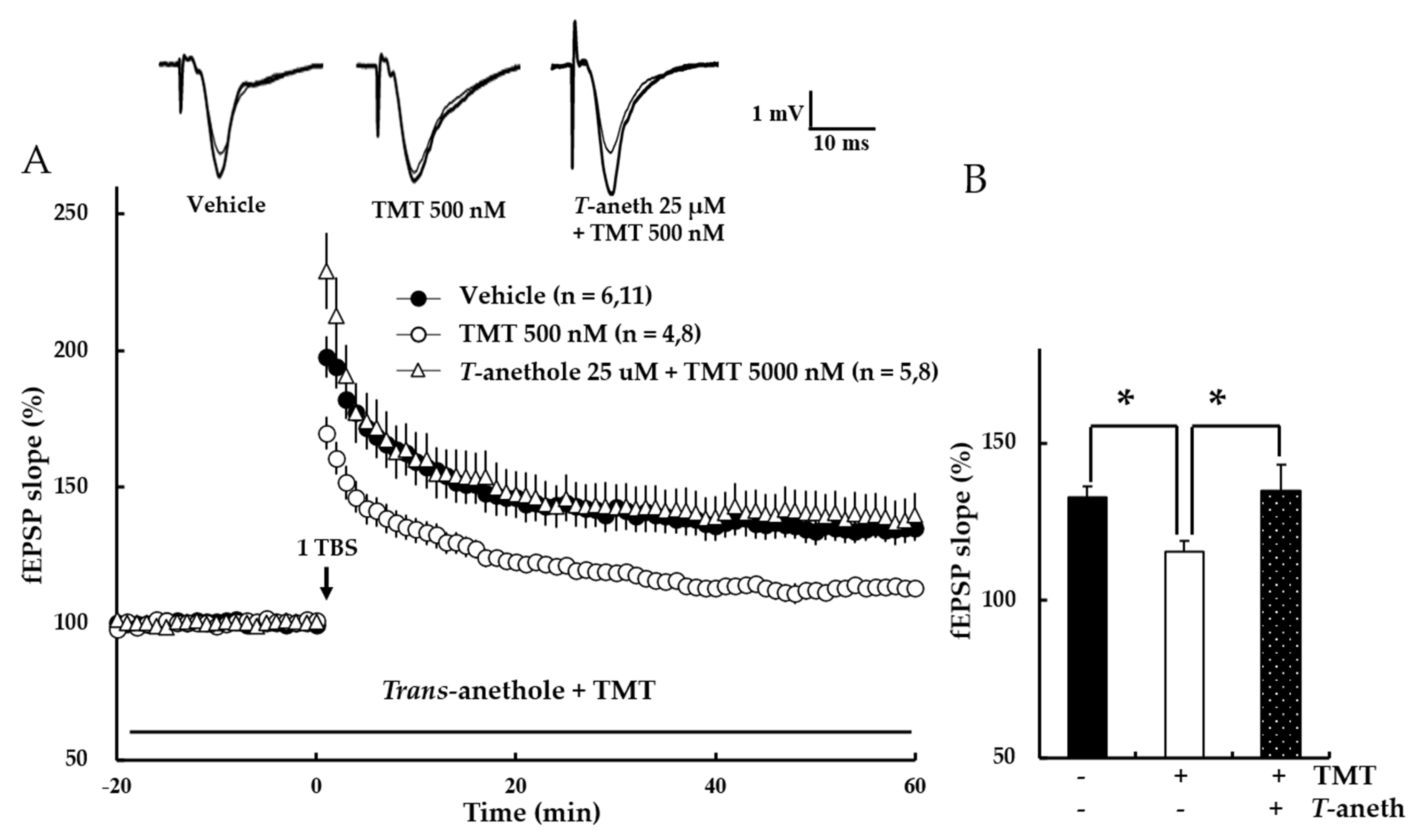
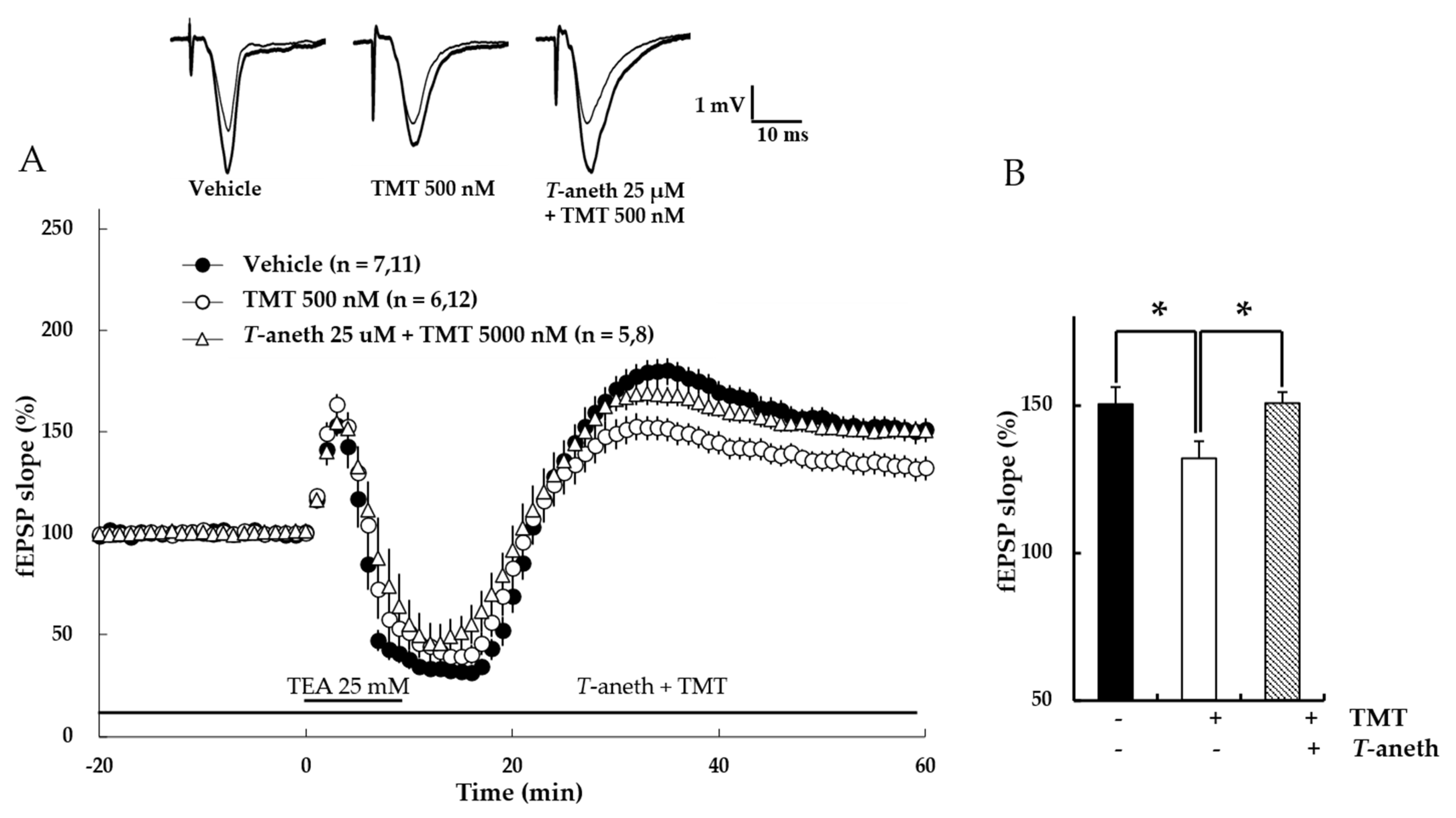
3.7. Trans-Anethole Blocks the TMT-Induced Impairment in NMDA Receptor-Dependent LTP
3.8. Trans-Anethole Blocks TMT-Induced NMDA Receptor-Independent LTP Impairment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.; Lim, C.L.; Zaki, M.; Jaenicke, S.; Chuah, G.K. A Dual-Functional Catalyst for Cascade Meerwein-Pondorf-Verley Reduction and Dehydration of 4’-Methoxypropiophenone to Anethole. ChemSusChem 2018, 11, 3007–3017. [Google Scholar] [CrossRef] [PubMed]
- Vendramin, V.; Pesce, A.; Vincenzi, S. Anethole Stability in Aniseed Spirits: Storage Condition Repercussions on Commercial Products. Beverages 2021, 7, 73. [Google Scholar] [CrossRef]
- Han, A.Y.; Lee, H.S.; Seol, G.H. Foeniculum vulgare Mill. increases cytosolic Ca(2+) concentration and inhibits store-operated Ca(2+) entry in vascular endothelial cells. Biomed. Pharm. 2016, 84, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Pourgholami, M.H.; Majzoob, S.; Javadi, M.; Kamalinejad, M.; Fanaee, G.H.; Sayyah, M. The fruit essential oil of Pimpinella anisum exerts anticonvulsant effects in mice. J. Ethnopharmacol. 1999, 66, 211–215. [Google Scholar] [CrossRef]
- Chen, C.H.; deGraffenried, L.A. Anethole suppressed cell survival and induced apoptosis in human breast cancer cells independent of estrogen receptor status. Phytomedicine 2012, 19, 763–767. [Google Scholar] [CrossRef]
- Domiciano, T.P.; Dalalio, M.M.; Silva, E.L.; Ritter, A.M.; Estevao-Silva, C.F.; Ramos, F.S.; Caparroz-Assef, S.M.; Cuman, R.K.; Bersani-Amado, C.A. Inhibitory effect of anethole in nonimmune acute inflammation. Naunyn. Schmiedebergs Arch. Pharmacol. 2013, 386, 331–338. [Google Scholar] [CrossRef]
- Ritter, A.M.; Domiciano, T.P.; Verri, W.A., Jr.; Zarpelon, A.C.; da Silva, L.G.; Barbosa, C.P.; Natali, M.R.; Cuman, R.K.; Bersani-Amado, C.A. Antihypernociceptive activity of anethole in experimental inflammatory pain. Inflammopharmacology 2013, 21, 187–197. [Google Scholar] [CrossRef]
- Shin, S.C.; Lee, D.U. Ameliorating effect of new constituents from the hooks of Uncaria rhynchophylla on scopolamine-induced memory impairment. Chin. J. Nat. Med. 2013, 11, 391–395. [Google Scholar] [CrossRef]
- Ryu, S.; Seol, G.H.; Park, H.; Choi, I.Y. Trans-anethole protects cortical neuronal cells against oxygen-glucose deprivation/reoxygenation. Neurol. Sci. 2014, 35, 1541–1547. [Google Scholar] [CrossRef]
- Casalbore, P.; Barone, I.; Felsani, A.; D’Agnano, I.; Michetti, F.; Maira, G.; Cenciarelli, C. Neural stem cells modified to express BDNF antagonize trimethyltin-induced neurotoxicity through PI3K/Akt and MAP kinase pathways. J. Cell Physiol. 2010, 224, 710–721. [Google Scholar] [CrossRef]
- Little, A.R.; Miller, D.B.; Li, S.; Kashon, M.L.; O’Callaghan, J.P. Trimethyltin-induced neurotoxicity: Gene expression pathway analysis, q-RT-PCR and immunoblotting reveal early effects associated with hippocampal damage and gliosis. Neurotoxicol. Teratol. 2012, 34, 72–82. [Google Scholar] [CrossRef]
- Mignini, F.; Nasuti, C.; Artico, M.; Giovannetti, F.; Fabrizi, C.; Fumagalli, L.; Iannetti, G.; Pompili, E. Effects of trimethyltin on hippocampal dopaminergic markers and cognitive behaviour. Int. J. Immunopathol. Pharmacol. 2012, 25, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Ishida, N.; Akaike, M.; Tsutsumi, S.; Kanai, H.; Masui, A.; Sadamatsu, M.; Kuroda, Y.; Watanabe, Y.; McEwen, B.S.; Kato, N. Trimethyltin syndrome as a hippocampal degeneration model: Temporal changes and neurochemical features of seizure susceptibility and learning impairment. Neuroscience 1997, 81, 1183–1191. [Google Scholar] [CrossRef]
- Earley, B.; Burke, M.; Leonard, B.E. Behavioural, biochemical and histological effects of trimethyltin (TMT) induced brain damage in the rat. Neurochem. Int. 1992, 21, 351–366. [Google Scholar] [CrossRef]
- Chang, L.W.; Wenger, G.R.; McMillan, D.E.; Dyer, R.S. Species and strain comparison of acute neurotoxic effects of trimethyltin in mice and rats. Neurobehav. Toxicol. Teratol. 1983, 5, 337–350. [Google Scholar]
- Larkman, A.U.; Jack, J.J. Synaptic plasticity: Hippocampal LTP. Curr. Opin. Neurobiol. 1995, 5, 324–334. [Google Scholar] [CrossRef]
- Kirkwood, A.; Dudek, S.M.; Gold, J.T.; Aizenman, C.D.; Bear, M.F. Common forms of synaptic plasticity in the hippocampus and neocortex in vitro. Science 1993, 260, 1518–1521. [Google Scholar] [CrossRef]
- Yun, S.H.; Huh, K.; Jung, M.W. Selective enhancement of non-NMDA receptor-mediated responses following induction of long-term potentiation in entorhinal cortex. Synapse 2000, 35, 1–7. [Google Scholar] [CrossRef]
- Morgan, S.L.; Teyler, T.J. Electrical stimuli patterned after the theta-rhythm induce multiple forms of LTP. J. Neurophysiol. 2001, 86, 1289–1296. [Google Scholar] [CrossRef]
- Citri, A.; Malenka, R.C. Synaptic plasticity: Multiple forms, functions, and mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef] [Green Version]
- Tewari, S.; Majumdar, K. A mathematical model for astrocytes mediated LTP at single hippocampal synapses. J. Comput. Neurosci. 2012, 33, 341–370. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, E.; Okada, T. TEA-induced long-term potentiation at hippocampal mossy fiber-CA3 synapses: Characteristics of its induction and expression. Brain Res. 2009, 1247, 21–27. [Google Scholar] [CrossRef]
- Suzuki, E.; Okada, T. Regional differences in GABAergic modulation for TEA-induced synaptic plasticity in rat hippocampal CA1, CA3 and dentate gyrus. Neurosci. Res. 2007, 59, 183–190. [Google Scholar] [CrossRef]
- Platt, B.; Behnisch, T.; Reymann, K.G. Metabotropic glutamate receptors are involved in TEA-induced long-term potentiation in area CA1 of the hippocampus. Neuropharmacology 1995, 34, 1339–1341. [Google Scholar] [CrossRef]
- Aniksztejn, L.; Ben-Ari, Y. Novel form of long-term potentiation produced by a K+ channel blocker in the hippocampus. Nature 1991, 349, 67–69. [Google Scholar] [CrossRef]
- Kim, H.; Han, S.H.; Quan, H.Y.; Jung, Y.J.; An, J.; Kang, P.; Park, J.B.; Yoon, B.J.; Seol, G.H.; Min, S.S. Bryostatin-1 promotes long-term potentiation via activation of PKCalpha and PKCepsilon in the hippocampus. Neuroscience 2012, 226, 348–355. [Google Scholar] [CrossRef]
- Kim, E.C.; Lee, M.J.; Shin, S.Y.; Seol, G.H.; Han, S.H.; Yee, J.; Kim, C.; Min, S.S. Phorbol 12-Myristate 13-Acetate Enhances Long-Term Potentiation in the Hippocampus through Activation of Protein Kinase Cdelta and epsilon. Korean J. Physiol. Pharmacol. 2013, 17, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Xie, X.; Wang, Z.; Berger, T.W. Differential effect of TEA on long-term synaptic modification in hippocampal CA1 and dentate gyrus in vitro. Neurobiol. Learn. Mem. 2001, 76, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Huguenard, J.R. Low-threshold calcium currents in central nervous system neurons. Annu. Rev. Physiol. 1996, 58, 329–348. [Google Scholar] [CrossRef]
- Onuma, H.; Lu, Y.F.; Tomizawa, K.; Moriwaki, A.; Tokuda, M.; Hatase, O.; Matsui, H. A calcineurin inhibitor, FK506, blocks voltage-gated calcium channel-dependent LTP in the hippocampus. Neurosci. Res. 1998, 30, 313–319. [Google Scholar] [CrossRef]
- Kruger, K.; Diepgrond, V.; Ahnefeld, M.; Wackerbeck, C.; Madeja, M.; Binding, N.; Musshoff, U. Blockade of glutamatergic and GABAergic receptor channels by trimethyltin chloride. Br. J. Pharmacol. 2005, 144, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.; An, J.; Seol, G.H.; Han, S.H.; Yee, J.; Min, S.S. Trans-Anethole Alleviates Trimethyltin Chloride-Induced Impairments in Long-Term Potentiation. Pharmaceutics 2022, 14, 1422. https://doi.org/10.3390/pharmaceutics14071422
Chang W, An J, Seol GH, Han SH, Yee J, Min SS. Trans-Anethole Alleviates Trimethyltin Chloride-Induced Impairments in Long-Term Potentiation. Pharmaceutics. 2022; 14(7):1422. https://doi.org/10.3390/pharmaceutics14071422
Chicago/Turabian StyleChang, Wonseok, Jihua An, Geun Hee Seol, Seung Ho Han, Jaeyong Yee, and Sun Seek Min. 2022. "Trans-Anethole Alleviates Trimethyltin Chloride-Induced Impairments in Long-Term Potentiation" Pharmaceutics 14, no. 7: 1422. https://doi.org/10.3390/pharmaceutics14071422
APA StyleChang, W., An, J., Seol, G. H., Han, S. H., Yee, J., & Min, S. S. (2022). Trans-Anethole Alleviates Trimethyltin Chloride-Induced Impairments in Long-Term Potentiation. Pharmaceutics, 14(7), 1422. https://doi.org/10.3390/pharmaceutics14071422