
Trans-Cinnamaldehyde Eluting Porous Silicon Microparticles Mitigate Cariogenic Biofilms

 , ,
, ,  , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
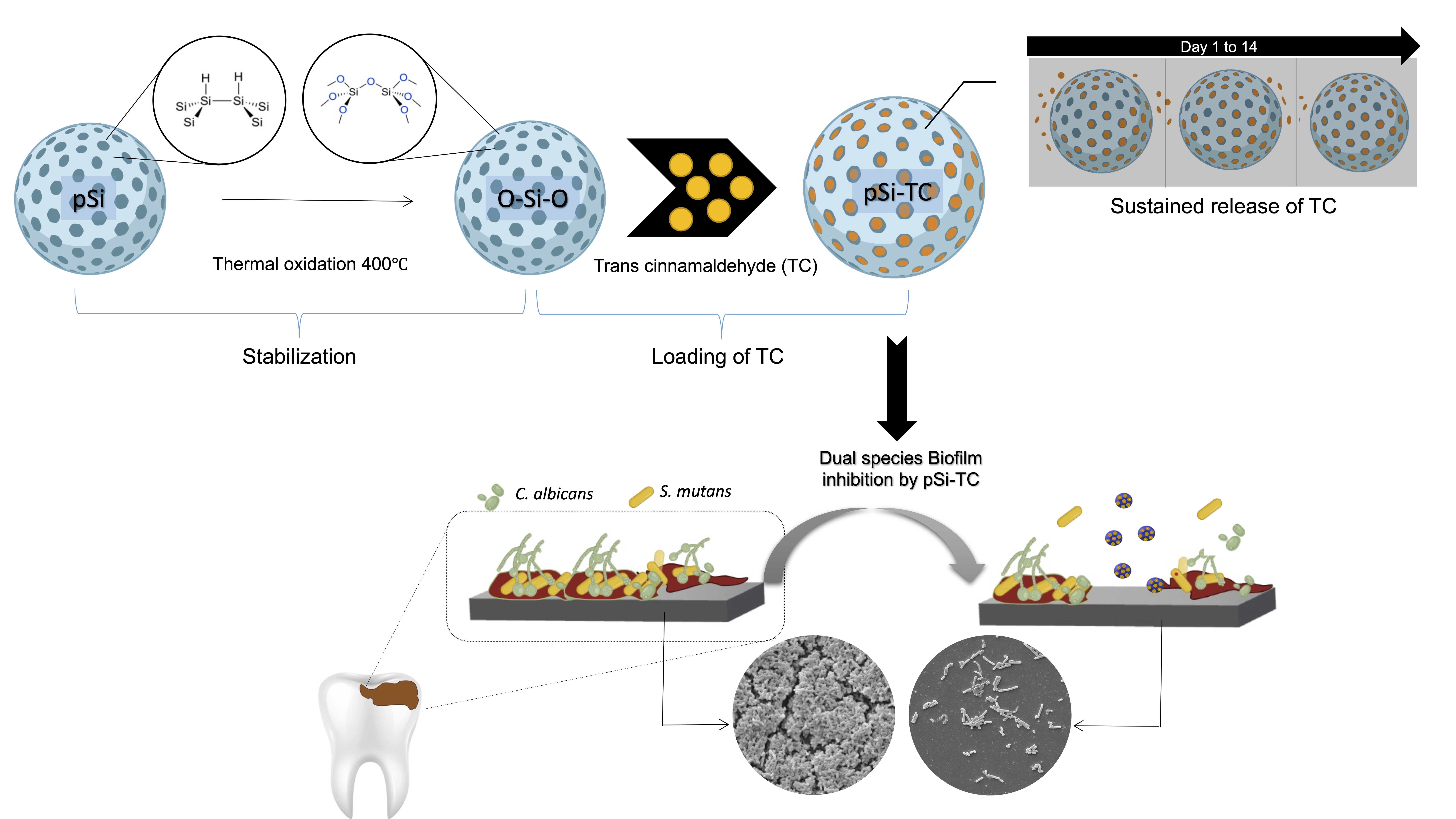
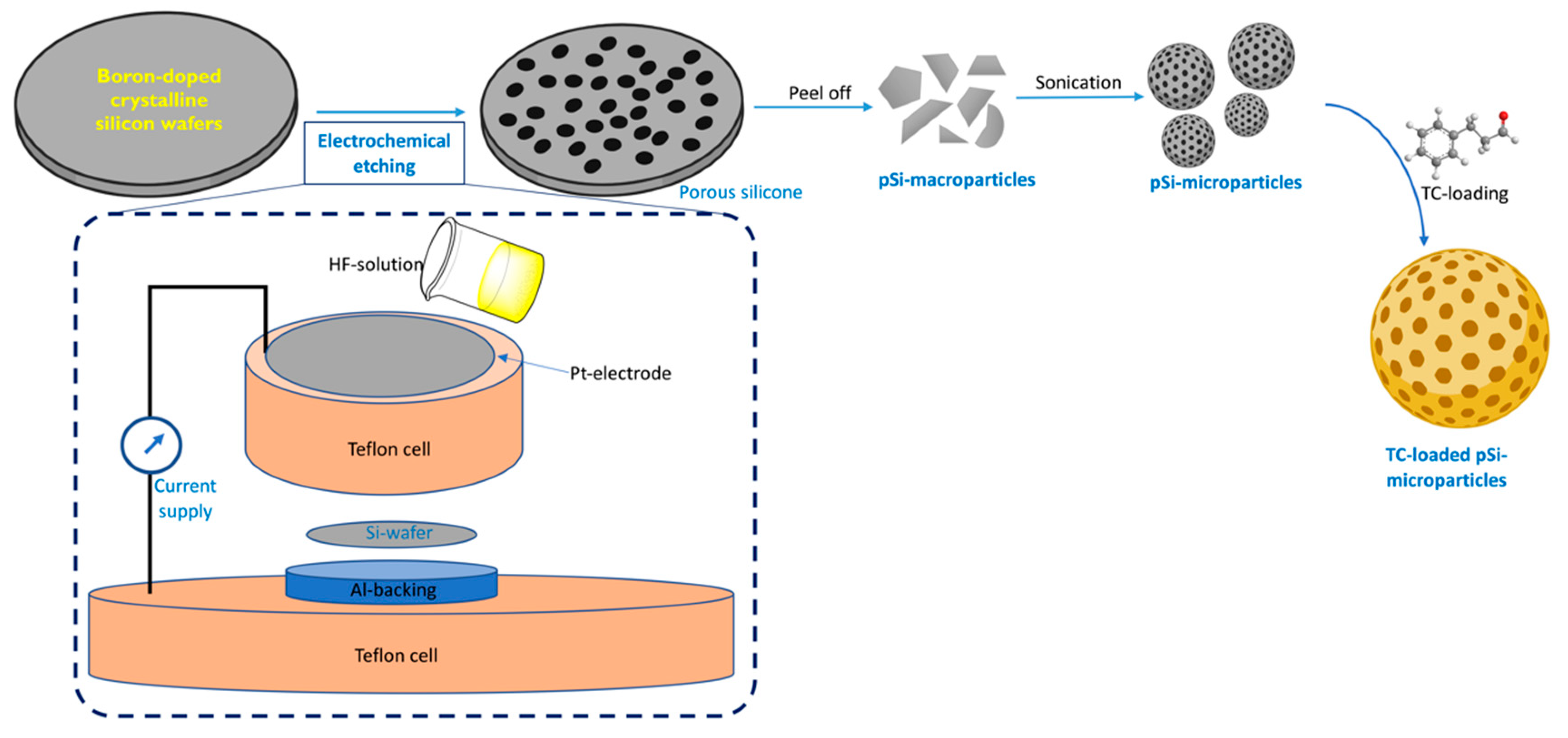
2.1. Synthesis and Characterization of pSi Particles
2.2. Characterization of Trans-Cinnamaldehyde Loaded pSi Particles
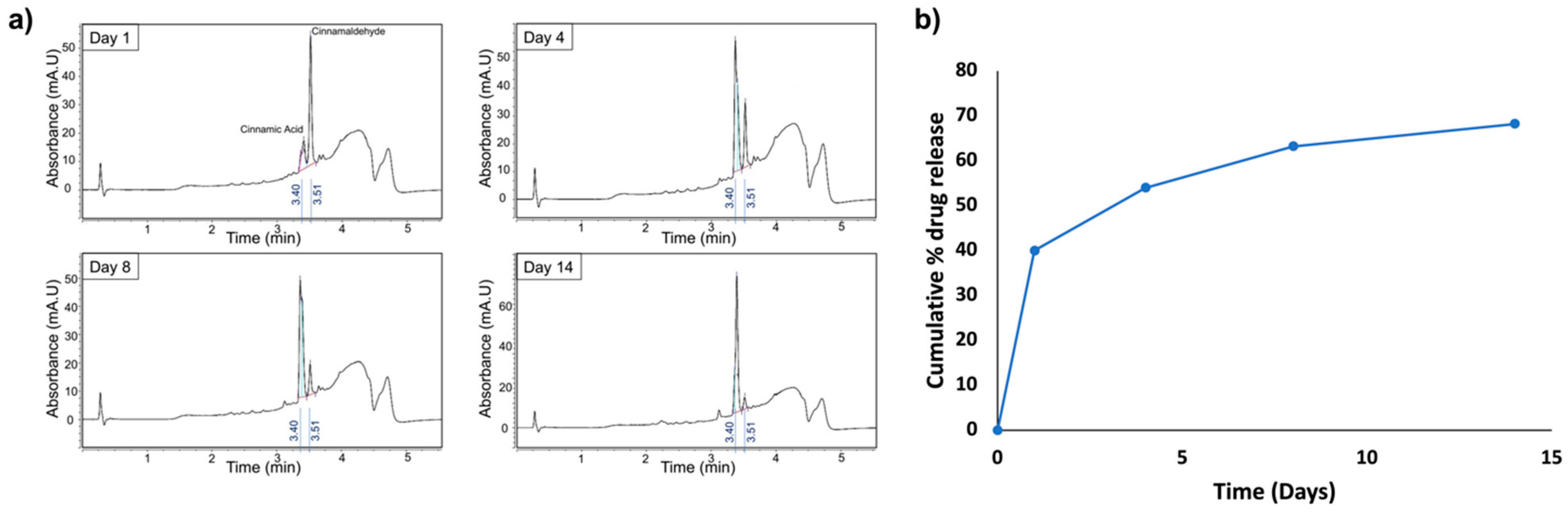
2.3. Trans-Cinnamaldehyde Loading and Release Kinetics
2.4. Microbial Strains and Culture Conditions
2.5. Microbial Growth
2.6. Biofilm Formation
2.7. Biofilm Microbial Composition
2.8. Acid Production
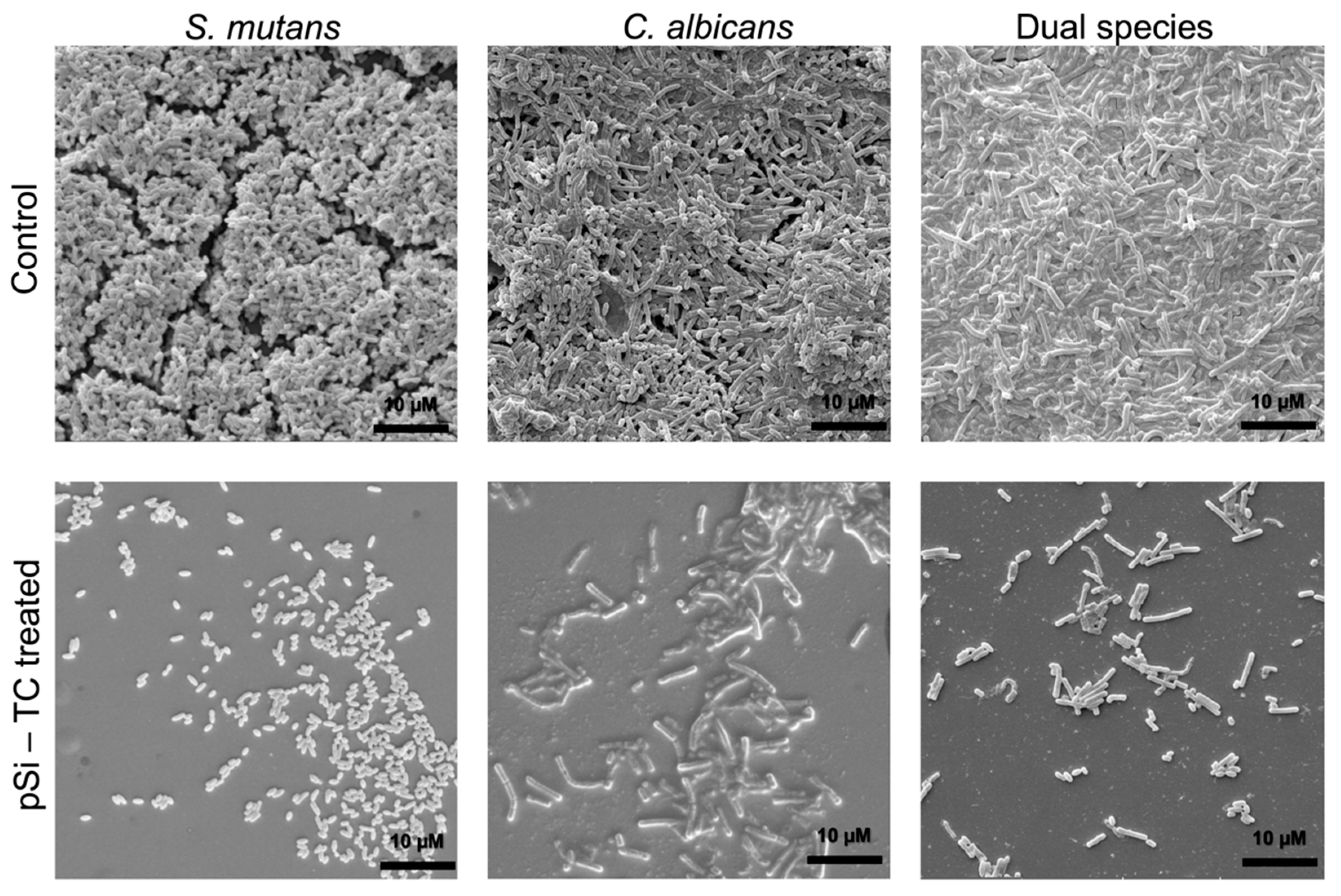
2.9. Scanning Electron Microscopic (SEM) Imaging
2.10. Gene Regulation Studies
2.11. Statistical Analysis
3. Results and Discussion
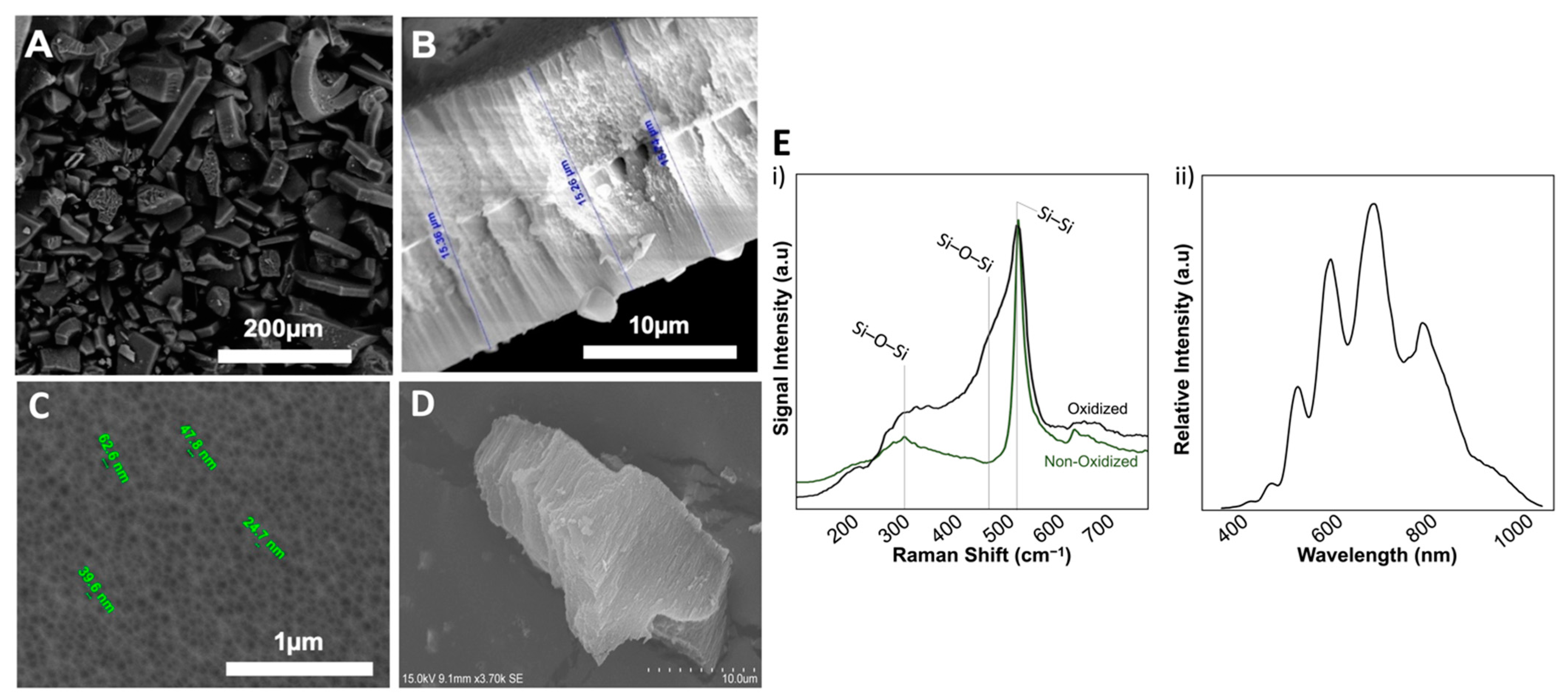
3.1. Characterization of pSi Particles
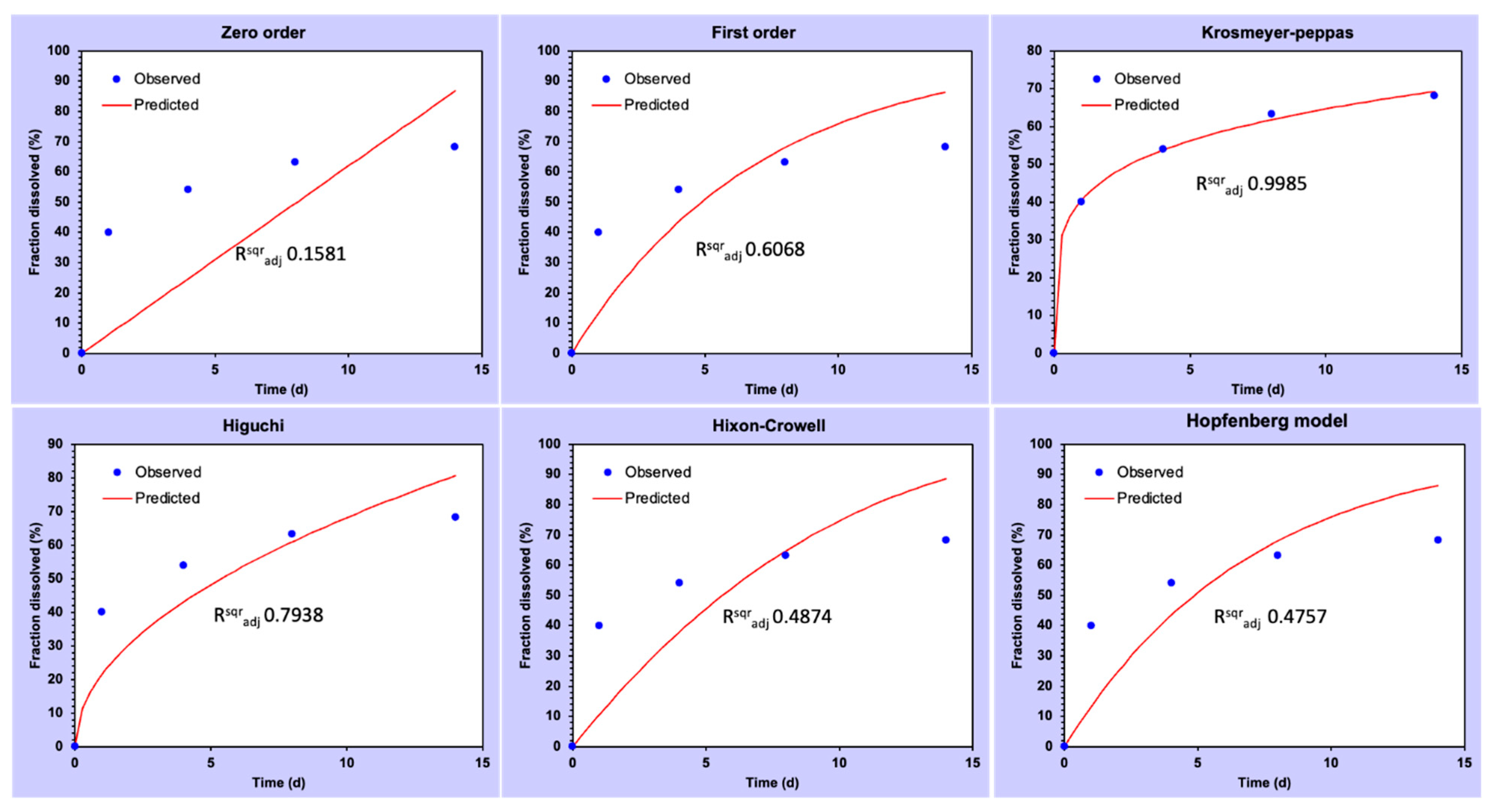
3.2. Drug Loading and Release Kinetics
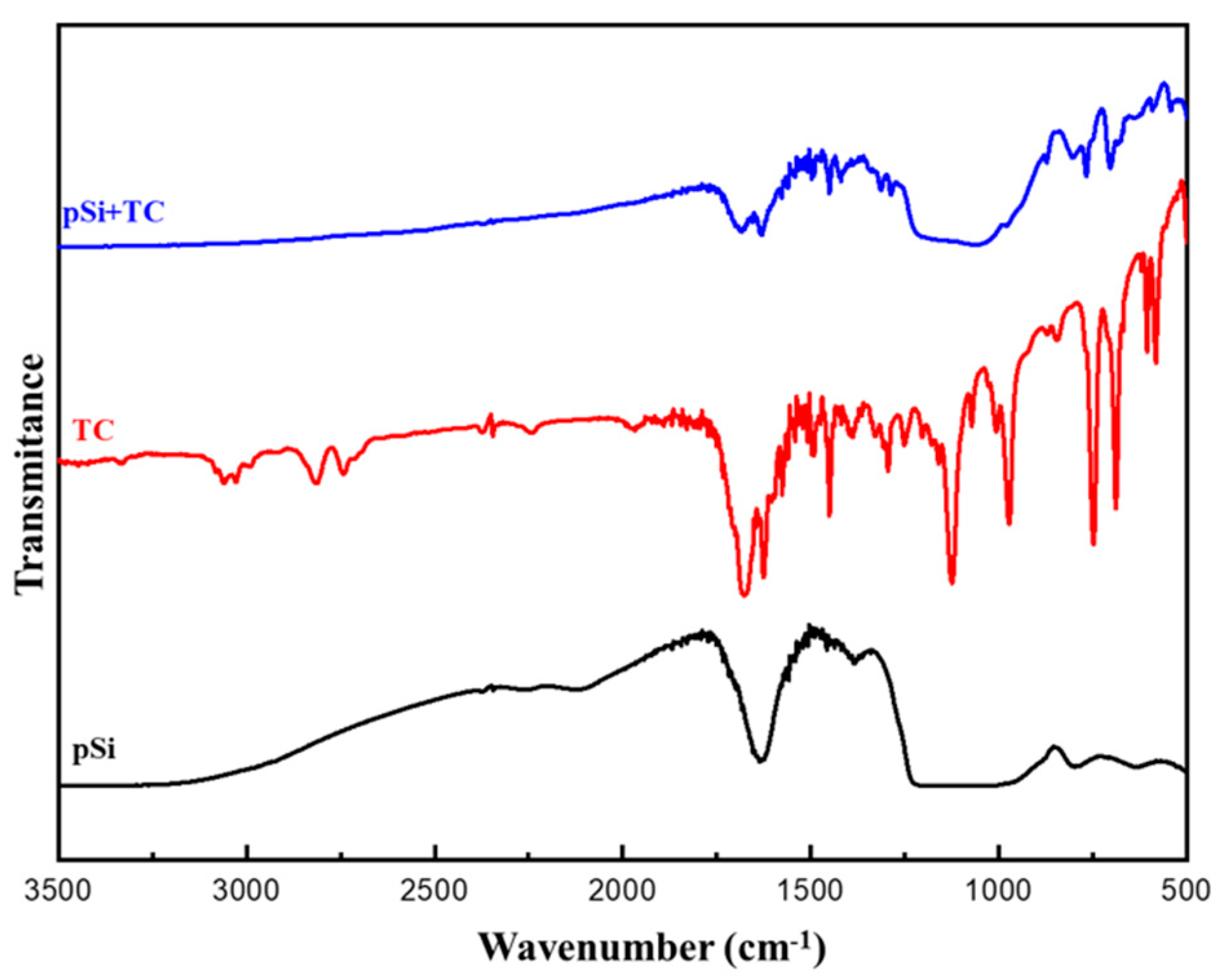
3.3. FTIR-Spectra of Trans-Cinnamaldehyde Loaded pSi Particles
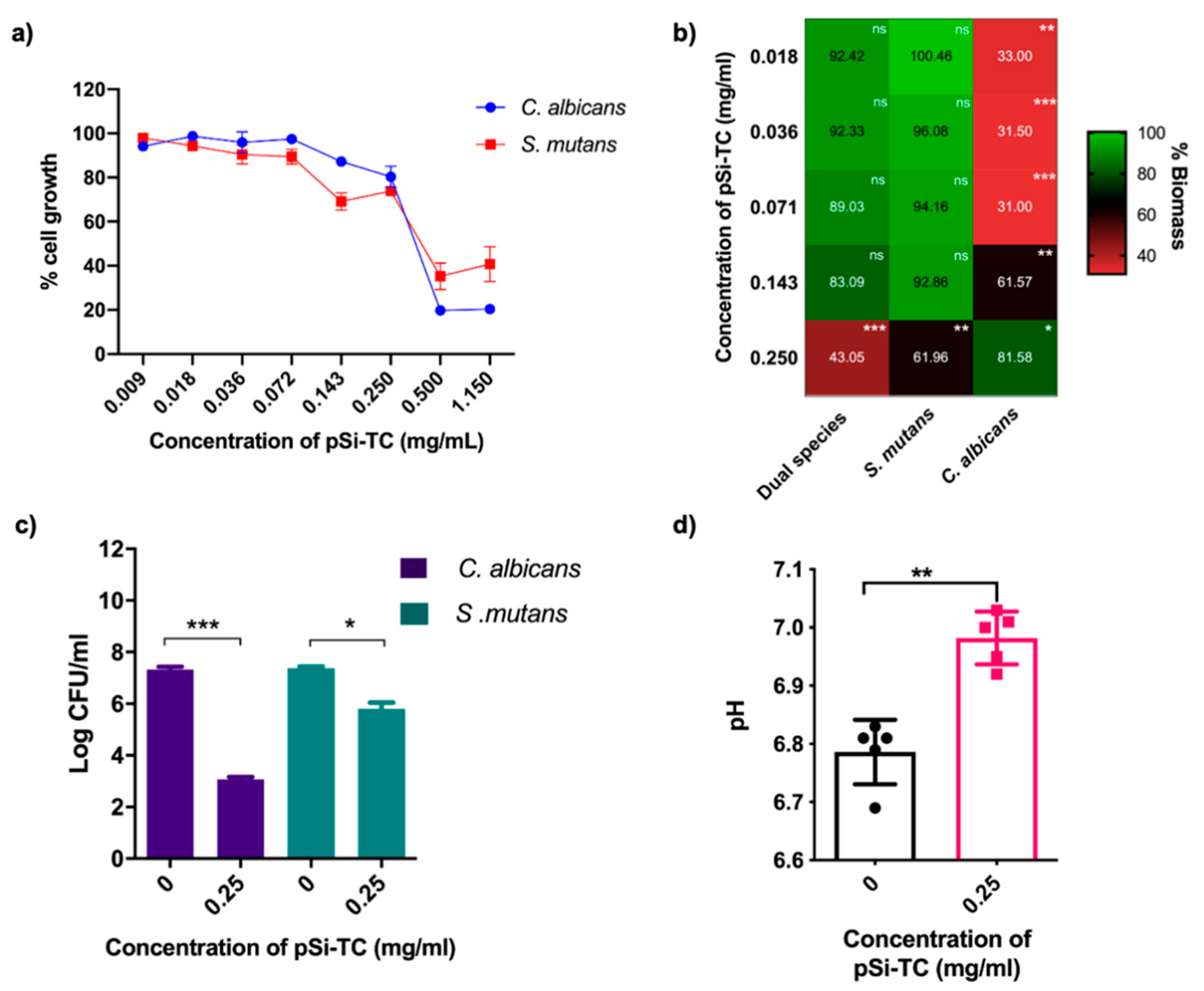
3.4. Sub-Inhibitory Concentration of pSi-TC Potently Inhibits Biofilms and Acid Production
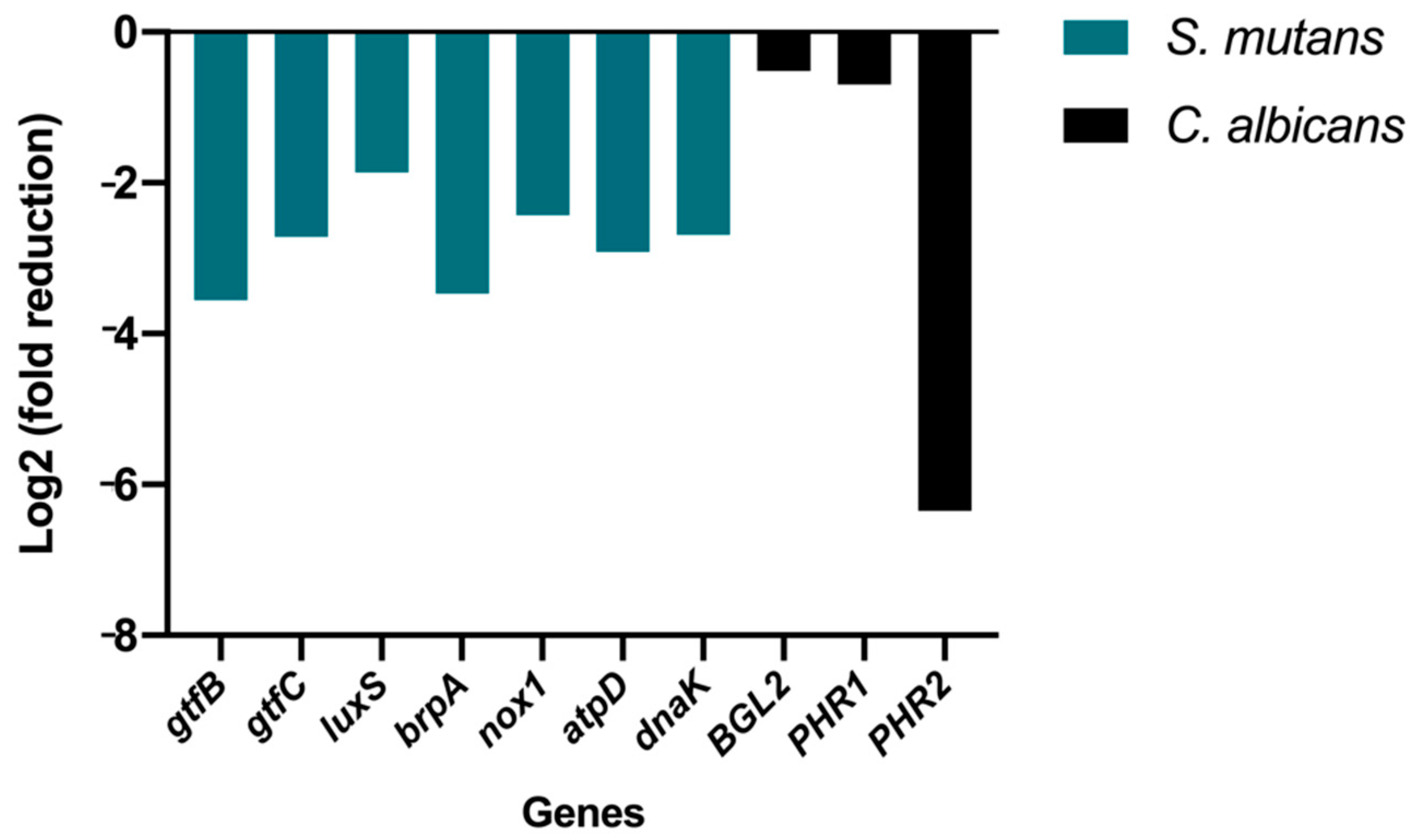
3.5. Gene Expression Analysis
4. Conclusions
- Porous silicon particles are an innovative and effective approach to carrying and releasing TC.
- pSi-TC successfully inhibited the development of cross-kingdom biofilms of S. mutans and C. albicans and its acid production.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vachirarojpisan, T.; Shinada, K.; Kawaguchi, Y.; Laungwechakan, P.; Somkote, T.; Detsomboonrat, P. Early childhood caries in children aged 6–19 months. Community Dent. Oral Epidemiol. 2004, 32, 133–142. [Google Scholar] [CrossRef]
- Bowen, W.H.; Burne, R.A.; Wu, H.; Koo, H. Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Bijle, M.N.; Neelakantan, P.; Ekambaram, M.; Lo, E.; Yiu, C. Effect of a novel synbiotic on Streptococcus mutans. Sci. Rep. 2020, 10, 7951. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Liao, Y.; Brandt, B.W.; Wei, X.; Liu, H.; Crielaard, W.; van Loveren, C.; Deng, D.M. The Fitness Cost of Fluoride Resistance for Different Streptococcus mutans Strains in Biofilms. Front. Microbiol. 2017, 8, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatzigiannidou, I.; Teughels, W.; van de Wiele, T.; Boon, N. Oral biofilms exposure to chlorhexidine results in altered microbial composition and metabolic profile. NPJ Biofilms Microbiomes 2020, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Bonez, P.C.; Dos Santos Alves, C.F.; Dalmolin, T.V.; Agertt, V.A.; Mizdal, C.R.; Flores, V.; Marques, J.B.; Santos, R.C.; Anraku de Campos, M.M. Chlorhexidine activity against bacterial biofilms. Am. J. Infect. Control. 2013, 41, e119–e122. [Google Scholar] [CrossRef]
- Ten Cate, J.M. Novel anticaries and remineralizing agents: Prospects for the future. J. Dent. Res. 2012, 91, 813–815. [Google Scholar] [CrossRef]
- Kot, B.; Sytykiewicz, H.; Sprawka, I.; Witeska, M. Effect of trans-Cinnamaldehyde on Methicillin-Resistant Staphylococcus aureus Biofilm Formation: Metabolic Activity Assessment and Analysis of the Biofilm-Associated Genes Expression. Int. J. Mol. Sci. 2019, 21, 102. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.A.K.S.; Rudden, M.; Smyth, T.J.; Dooley, J.; Marchant, R.; Banat, I.M. Natural quorum sensing inhibitors effectively downregulate gene expression of Pseudomonas aeruginosa virulence factors. Appl. Microbiol. Biotechnol. 2019, 103, 3521–3535. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M.; Malheiro, J.; Grenho, L.; Fernandes, M.H.; Simões, M. Cytotoxicity and antimicrobial action of selected phytochemicals against planktonic and sessile Streptococcus mutans. PeerJ 2018, 6, e4872. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, A.R.; Vasudevan, S.; Shanmugam, K.; Lévesque, C.M.; Solomon, A.P.; Neelakantan, P. Combinatorial effects of trans-cinnamaldehyde with fluoride and chlorhexidine on Streptococcus mutans. J. Appl. Microbiol. 2021, 130, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Matinlinna, J.P.; Lévesque, C.M.; Neelakantan, P. Trans-Cinnamaldehyde Attenuates Enterococcus faecalis Virulence and Inhibits Biofilm Formation. J. Antibiot. 2021, 10, 702. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, Y.; Hasumi, Y.; Abe, S.; Nishiyama, Y. The effect of cinnamaldehyde on the growth and the morphology of Candida albicans. Med. Mol. Morphol. 2013, 46, 8–13. [Google Scholar] [CrossRef]
- Kohane, D.S. Microparticles and nanoparticles for drug delivery. Biotechnol. Bioeng. 2007, 96, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Bimbo, L.M.; Mäkilä, E.; Laaksonen, T.; Lehto, V.P.; Salonen, J.; Hirvonen, J.; Santos, H.A. Drug permeation across intestinal epithelial cells using porous silicon nanoparticles. Biomaterials 2011, 32, 2625–2633. [Google Scholar] [CrossRef] [PubMed]
- Salonen, J.; Kaukonen, A.M.; Hirvonen, J.; Lehto, V.P. Mesoporous Silicon in Drug Delivery Applications. J. Pharm. Sci. 2008, 97, 632–653. [Google Scholar] [CrossRef]
- Trewyn, B.G.; Giri, S.; Slowing, I.I.; Lin, V.S. Mesoporous silica nanoparticle based controlled release, drug delivery, and biosensor systems. Chem. Commun. 2007, 3236–3245. [Google Scholar] [CrossRef]
- Dutilleul, P.C.; Périère, D.D.; Cuisinier, F.J.; Cunin, F.; Gergely, C. Porous silicon scaffolds for stem cells growth and osteodifferentiation. In Porous Silicon for Biomedical Applications; Santos, H.A., Ed.; Woodhead Publishing: Cambridge, UK, 2014; pp. 486–506. [Google Scholar]
- Hamada, S.; Slade, H.D. Slade, Biology, immunology, and cariogenicity of Streptococcus mutans. Microbiol. Rev. 1980, 44, 331–384. [Google Scholar] [CrossRef]
- Du, Q.; Fu, M.; Zhou, Y.; Cao, Y.; Guo, T.; Zhou, Z.; Li, M.; Peng, X.; Zheng, X.; Li, Y.; et al. Sucrose promotes caries progression by disrupting the microecological balance in oral biofilms: An in vitro study. Sci. Rep. 2020, 10, 2961. [Google Scholar] [CrossRef] [Green Version]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar] [CrossRef]
- Ellepola, K.; Truong, T.; Liu, Y.; Lin, Q.; Lim, T.K.; Lee, Y.M.; Cao, T.; Koo, H.; Seneviratne, C.J. Multi-omics Analyses Reveal Synergistic Carbohydrate Metabolism in Streptococcus mutans-Candida albicans Mixed-Species Biofilms. Infect. Immun. 2019, 87, e00339-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belli, W.A.; Marquis, R.E. Adaptation of Streptococcus mutans and Enterococcus hirae to acid stress in continuous culture. Appl. Environ. Microbiol. 1991, 57, 1134–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huffines, J.T.; Scoffield, J.A. Disruption of Streptococcus mutans and Candida albicans synergy by a commensal streptococcus. Sci. Rep. 2020, 10, 19661. [Google Scholar] [CrossRef] [PubMed]
- Haidary, M.S.; Awaz, B.M.; Emma, P.C.; Nihad, K.A.; Ahmad, M.R. Effect of coatings and surface modification on porous silicon nanoparticles for delivery of the anticancer drug tamoxifen. Microelectron. Eng. 2016, 161, 1–6. [Google Scholar] [CrossRef]
- Kim, D.; Sengupta, A.; Niepa, T.H.R.; Lee, B.H.; Weljie, A.; Freitas-Blanco, V.S.; Murata, R.M.; Stebe, K.J.; Lee, D.; Koo, H. Candida albicans stimulates Streptococcus mutans microcolony development via cross-kingdom biofilm-derived metabolites. Sci. Rep. 2017, 7, 41332. [Google Scholar] [CrossRef]
- Rodríguez-Tudela, J.L.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Denning, D.; Donnelly, J.P.; Dupont, B.; Fegeler, W.; Moore, C.; et al. Method for the determination of minimum inhibitory concentration (MIC) by broth dilution of fermentative yeasts. Clin. Microbiol. Infect. 2003, 9, i–viii. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, D.R.; Gorup, L.F.; Silva, S.; Negri, M.; de Camargo, E.R.; Oliveira, R.; Barbosa, D.B.; Henriques, M.; Gorup, L.F.; Silva, S.; et al. Silver colloidal nanoparticles: Antifungal effect against adhered cells and biofilms of Candida albicans and Candida glabrata. J. Biofouling. 2011, 27, 711–719. [Google Scholar] [CrossRef]
- Rajasekar, V.; Darne, P.; Prabhune, A.; Kao, R.; Solomon, A.P.; Ramage, G.; Samaranayake, L.; Neelakantan, P. A curcumin-sophorolipid nanocomplex inhibits Candida albicans filamentation and biofilm development. Colloids Surf B Biointerfaces 2021, 200, 111617. [Google Scholar] [CrossRef]
- Eidt, G.; Andrade, C.G.; Negrini, T.C.; Arthur, R.A. Role of Candida albicans on enamel demineralization and on acidogenic potential of Streptococcus mutans in vitro biofilms. J. Appl. Oral Sci. 2019, 27, 1–8. [Google Scholar] [CrossRef] [Green Version]
- McInnes, S.J.P.; Voelcker, N.H. Porous silicon–polymer composites for cell culture and tissue engineering applications. In Porous Silicon for Biomedical Applications; Santos, H.A., Ed.; Woodhead Publishing: Cambridge, UK, 2014; pp. 420–469. [Google Scholar]
- Majeed, H.; Bian, Y.-Y.; Ali, B.; Jamil, A.; Majeed, U.; Khan, Q.F.; Iqbal, K.J.; Shoemaker, C.F.; Fang, Z. Essential oil encapsulations: Uses, procedures, and trends. RSC Adv. 2015, 5, 58449–58463. [Google Scholar] [CrossRef]
- Ruiz-Rico, M.; Pérez-Esteve, É.; Bernardos, A.; Sancenón, F.; Martínez-Máñez, R.; Marcos, M.D.; Barat, J.M. Enhanced antimicrobial activity of essential oil components immobilized on silica particles. Food Chem. 2017, 233, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Bernardos, A.; Marina, T.; Žáček, P.; Pérez-Esteve, É.; Martínez-Mañez, R.; Lhotka, M.; Kouřimská, L.; Pulkrábek, J.; Klouček, P. Antifungal effect of essential oil components against Aspergillus niger when loaded into silica mesoporous supports. J. Sci. Food Agric. 2015, 95, 2824–2831. [Google Scholar] [CrossRef] [PubMed]
- Dancil, K.-P.S.; Greiner, D.P.; Sailor, M.J. A Porous Silicon Optical Biosensor: Detection of Reversible Binding of IgG to a Protein A-Modified Surface. J. Am. Chem. Soc. 1999, 121, 7925–7930. [Google Scholar] [CrossRef]
- Sreeharsha, N.; Rajpoot, K.; Tekade, M.; Kalyane, D.; Nair, A.B.; Venugopala, K.N.; Tekade, R.K. Development of Metronidazole Loaded Chitosan Nanoparticles Using QbD Approach-A Novel and Potential Antibacterial Formulation. Pharmaceutics 2020, 12, 920. [Google Scholar] [CrossRef]
- He, Z.; Huang, Z.; Jiang, W.; Zhou, W. Antimicrobial Activity of Cinnamaldehyde on Streptococcus mutans Biofilms. Front. Microbiol. 2019, 10, 2241. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.N.; Khan, S.; Iqbal, J.; Khan, R.; Khan, A.U. Enhanced Killing and Antibiofilm Activity of Encapsulated Cinnamaldehyde against Candida albicans. Front. Microbiol. 2017, 8, 1641. [Google Scholar] [CrossRef]
- Ferhout, H.; Bohatier, J.; Guillot, J.; Chalchat, J.C. Antifungal Activity of Selected Essential Oils, Cinnamaldehyde and Carvacrol against Malassezia furfur and Candida albicans. J. Essent. Oil Res. 1999, 11, 119–129. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon, Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms in vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Lobo, C.I.V.; Rinaldi, T.B.; Christiano, C.M.S.; de Sales Leite, L.; Barbugli, P.A.; Klein, M.I. Dual-species biofilms of Streptococcus mutans and Candida albicans exhibit more biomass and are mutually beneficial compared with single-species biofilms. J. Oral Microbiol. 2019, 11, 1581520. [Google Scholar] [CrossRef] [Green Version]
- Sheng, J.; Marquis, R.E. Malolactic fermentation by Streptococcus mutans. FEMS Microbiol. Lett. 2007, 2721, 96–201. [Google Scholar]
- Poole, L.B.; Higuchi, M.; Shimada, M.; Calzi, M.L.; Kamio, Y. Streptococcus mutans H2O2-forming NADH oxidase is an alkyl hydroperoxide reductase protein. Free. Radic. Biol. Med. 2000, 28, 108–120. [Google Scholar] [CrossRef]
- Wen, Z.T.; Burne, R.A. LuxS-mediated signaling in Streptococcus mutans is involved in regulation of acid and oxidative stress tolerance and biofilm formation. J. Bacteriol. 2004, 186, 2682–2691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, K.; Sarveswari, H.B.; Udayashankar, A.; Swamy, S.S.; Pudipeddi, A.; Shanmugam, T.; Solomon, A.P.; Neelakantan, P. Guardian genes ensuring subsistence of oral Streptococcus mutans. Crit. Rev. Microbiol. 2020, 46, 475–491. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Biswas, I. Regulation of the glucosyltransferase (gtfBC) operon by CovR in Streptococcus mutans. J. Bacteriol. 2006, 188, 988–998. [Google Scholar] [CrossRef] [Green Version]
- Taff, H.T.; Nett, J.E.; Zarnowski, R.; Ross, K.M.; Sanchez, H.; Cain, M.T.; Hamaker, J.; Mitchell, A.P.; Andes, D.R. A Candida Biofilm-Induced Pathway for Matrix Glucan Delivery: Implications for Drug Resistance. PLoS Pathog. 2012, 8, e1002848. [Google Scholar] [CrossRef] [Green Version]
- Vediyappan, G.; Rossignol, T.; d’Enfert, C. Interaction of Candida albicans biofilms with antifungals: Transcriptional response and binding of antifungals to beta-glucans, Antimicrob. Agents Chemother. 2010, 54, 2096–2111. [Google Scholar] [CrossRef] [Green Version]
- Fonzi, W.A. PHR1 and PHR2 of Candida albicans encode putative glycosidases required for proper cross-linking of beta-1,3- and beta-1,6-glucans. J. Bacteriol. 1999, 181, 7070–7079. [Google Scholar] [CrossRef] [Green Version]
- Collart-Dutilleul, P.Y.; Secret, E.; Panayotov, I.; Deville de Périère, D.; Martín-Palma, R.J.; Torres-Costa, V.; Martin, M.; Gergely, C.; Durand, J.O.; Cunin, F.; et al. Adhesion and proliferation of human mesenchymal stem cells from dental pulp on porous silicon scaffolds. ACS Appl Mater Interfaces. 2014, 12, 1719–1728. [Google Scholar] [CrossRef]
- Soussi, I.; Mazouz, Z.; Collart-Dutilleul, P.Y.; Echabaane, M.; Martin, M.; Cloitre, T.; M’ghaieth, R.; Cuisinier, F.J.G.; Cunin, F.; Gergely, C.; et al. Electrochemical and optical investigation of dental pulp stem cell adhesion on modified porous silicon scaffolds. Colloids Surf B Biointerfaces 2019, 1, 489–497. [Google Scholar] [CrossRef]
- Collart-Dutilleul, P.Y.; Panayotov, I.; Secret, E.; Cunin, F.; Gergely, C.; Cuisinier, F.; Martin, M. Initial stem cell adhesion on porous silicon surface: Molecular architecture of actin cytoskeleton and filopodial growth. Nanoscale Res Lett. 2014, 10, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Gu, L.; von Maltzahn, G.; Ruoslahti, E.; Bhatia, S.N.; Sailor, M.J. Biodegradable luminescent porous silicon nanoparticles for in vivo applications. Nat. Mater. 2009, 8, 331–336. [Google Scholar] [CrossRef]
- Jurkić, L.M.; Cepanec, I.; Pavelić, S.K.; Pavelić, K. Biological and therapeutic effects of ortho-silicic acid and some ortho-silicic acid-releasing compounds: New perspectives for therapy. Nutr. Metab. 2013, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.E.; Na, J.Y.; Lee, J.S. Anti-inflammatory effects of trans-cinnamaldehyde on lipopolysaccharide-stimulated macrophage activation via MAPKs pathway regulation. Immunopharmacol. Immunotoxicol. 2018, 40, 219–224. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jailani, A.; Kalimuthu, S.; Rajasekar, V.; Ghosh, S.; Collart-Dutilleul, P.-Y.; Fatima, N.; Koo, H.; Solomon, A.P.; Cuisinier, F.; Neelakantan, P. Trans-Cinnamaldehyde Eluting Porous Silicon Microparticles Mitigate Cariogenic Biofilms. Pharmaceutics 2022, 14, 1428. https://doi.org/10.3390/pharmaceutics14071428
Jailani A, Kalimuthu S, Rajasekar V, Ghosh S, Collart-Dutilleul P-Y, Fatima N, Koo H, Solomon AP, Cuisinier F, Neelakantan P. Trans-Cinnamaldehyde Eluting Porous Silicon Microparticles Mitigate Cariogenic Biofilms. Pharmaceutics. 2022; 14(7):1428. https://doi.org/10.3390/pharmaceutics14071428
Chicago/Turabian StyleJailani, Afreen, Shanthini Kalimuthu, Vidhyashree Rajasekar, Sumanta Ghosh, Pierre-Yves Collart-Dutilleul, Naveen Fatima, Hyun Koo, Adline Princy Solomon, Frederic Cuisinier, and Prasanna Neelakantan. 2022. "Trans-Cinnamaldehyde Eluting Porous Silicon Microparticles Mitigate Cariogenic Biofilms" Pharmaceutics 14, no. 7: 1428. https://doi.org/10.3390/pharmaceutics14071428
APA StyleJailani, A., Kalimuthu, S., Rajasekar, V., Ghosh, S., Collart-Dutilleul, P. -Y., Fatima, N., Koo, H., Solomon, A. P., Cuisinier, F., & Neelakantan, P. (2022). Trans-Cinnamaldehyde Eluting Porous Silicon Microparticles Mitigate Cariogenic Biofilms. Pharmaceutics, 14(7), 1428. https://doi.org/10.3390/pharmaceutics14071428