
In Vitro Evaluation of Five Newly Isolated Bacteriophages against E. faecalis Biofilm for Their Potential Use against Post-Treatment Apical Periodontitis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Antimicrobial Agents
2.2. Bacteriophage Isolation
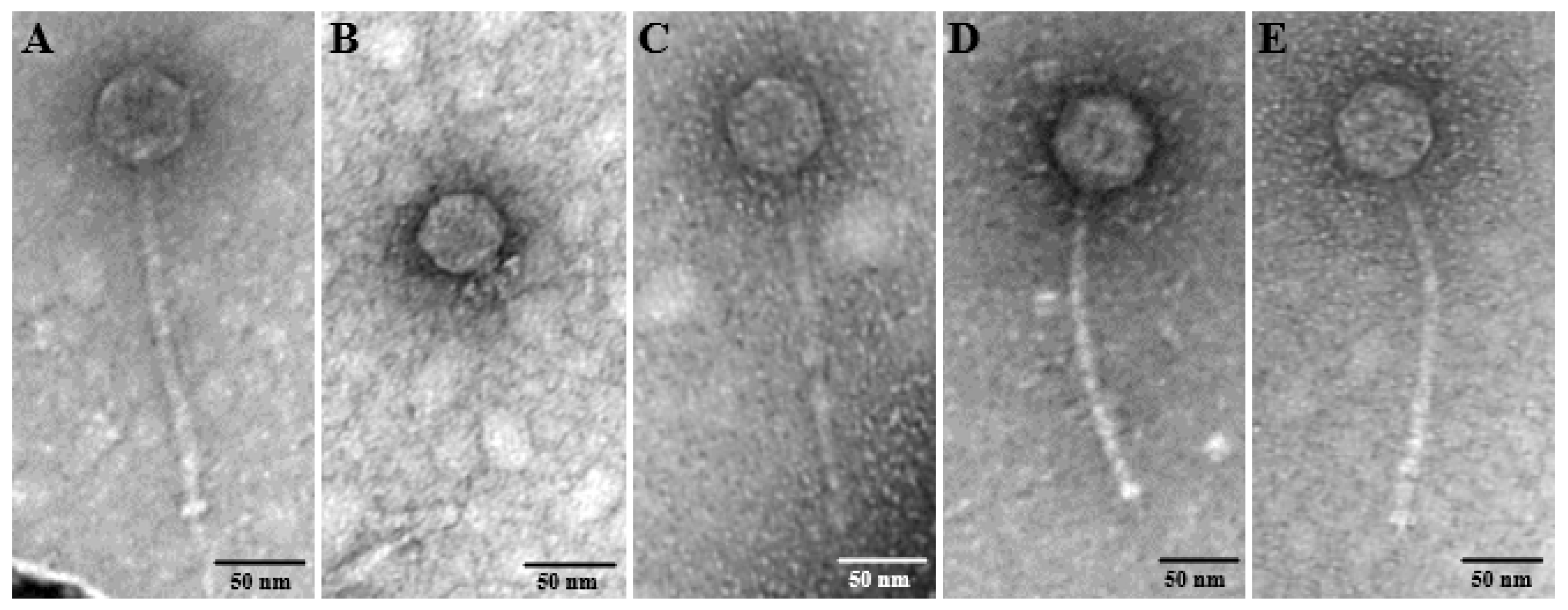
2.3. Morphological Analysis by Transmission Electron Microscopy
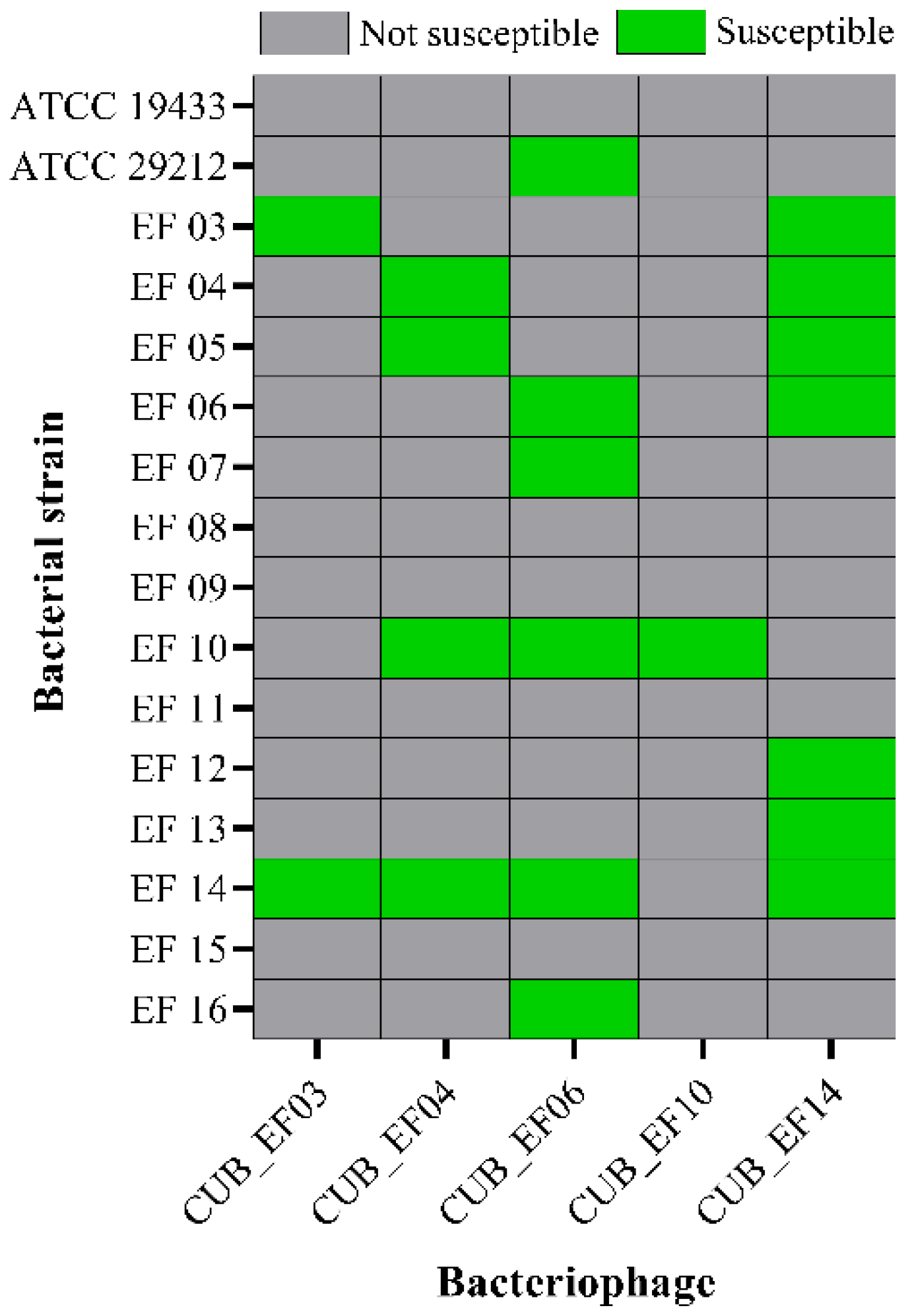
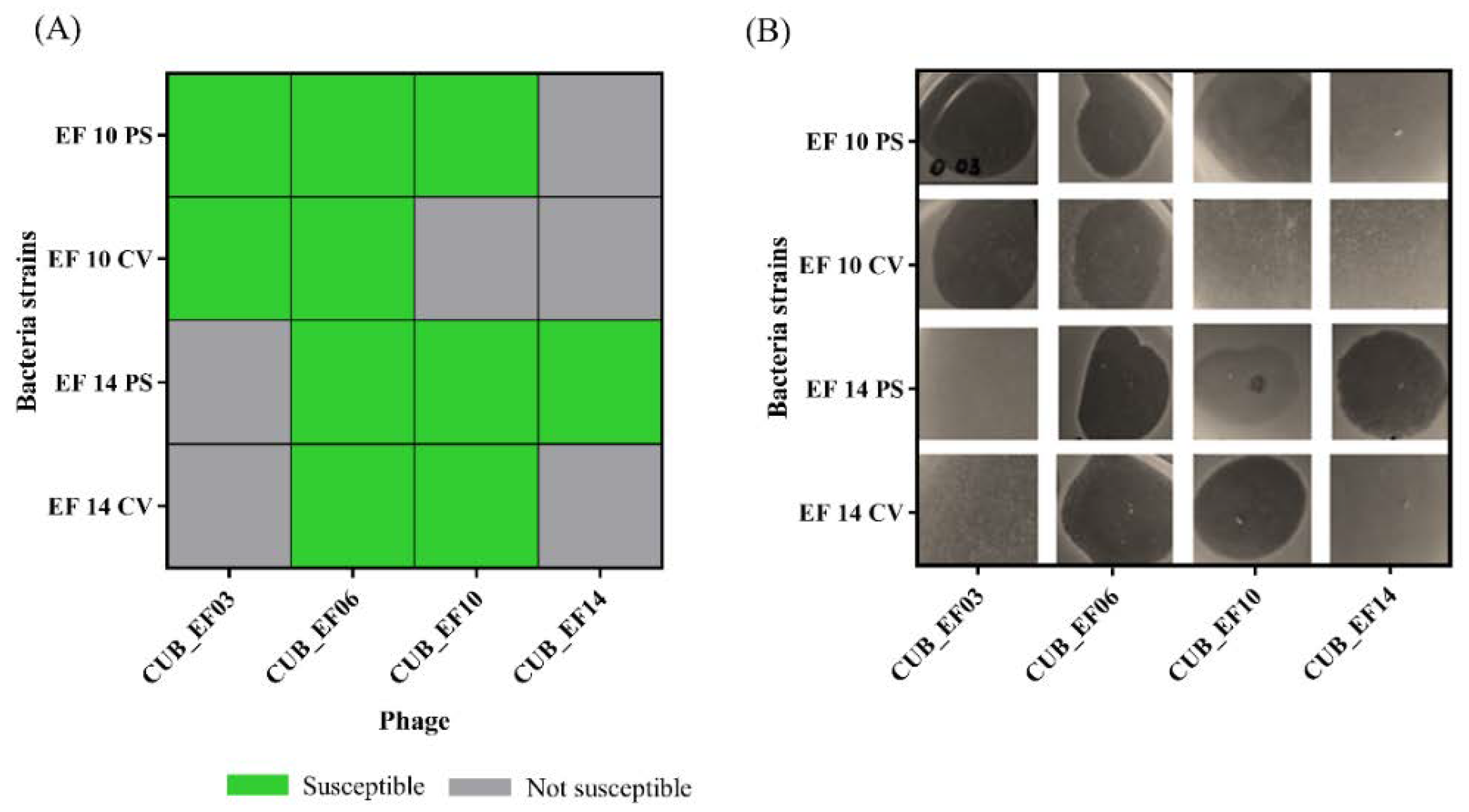
2.4. Host Range Analysis
2.5. Biofilm Time-Killing Assay
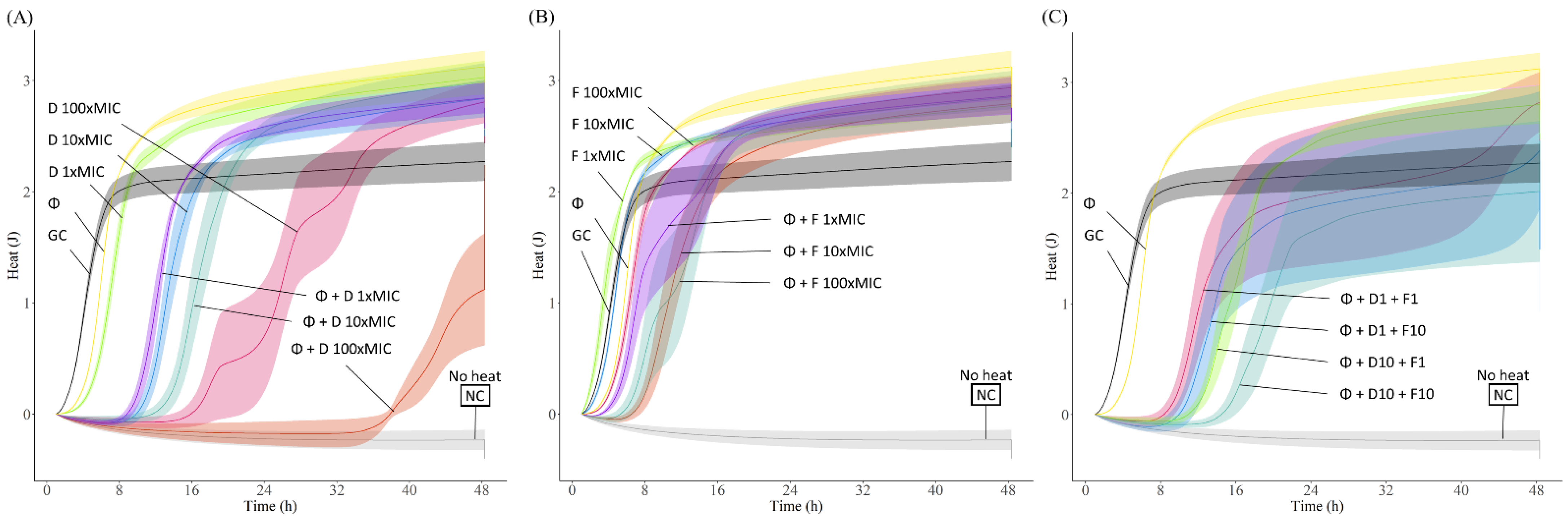
2.6. Phage Antimicrobial Activity Evaluated by Isothermal Microcalorimetry
2.7. Antibiotic Susceptibility Determination
2.8. Phage-Antibiotic Combinations against Biofilm
2.9. Assessment of Bacterial Resistance Development to Phage
3. Results
3.1. Phage Isolation and TEM Visualization
3.2. Host Range
3.3. Time-Killing Assay
3.4. Phage-Antibiotic Combinations against E. faecalis Biofilms
3.5. Phage-Antibiotic Combinations against Vancomycin and Gentamicin High Level Resistant E. faecalis Biofilm
3.6. Assessment of Bacterial Resistance Development to Phage
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Möller, A.J.; Fabricius, L.; Dahlén, G.; Ohman, A.E.; Heyden, G. Influence on periapical tissues of indigenous oral bacteria and necrotic pulp tissue in monkeys. Scand. J. Dent. Res. 1981, 89, 475–484. [Google Scholar] [CrossRef]
- Azuma, M.M.; Samuel, R.O.; Gomes-Filho, J.E.; Dezan-Junior, E.; Cintra, L.T. The role of IL-6 on apical periodontitis: A systematic review. Int. Endod. J. 2014, 47, 615–621. [Google Scholar] [CrossRef]
- Vire, D.E. Failure of endodontically treated teeth: Classification and evaluation. J. Endod. 1991, 17, 338–342. [Google Scholar] [CrossRef]
- Siqueira, J.F., Jr.; Rôças, I.N.; Ricucci, D.; Hülsmann, M. Causes and management of post-treatment apical periodontitis. Br. Dent. J. 2014, 216, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Hülsmann, M. Epidemiology of post-treatment disease. Endod. Top. 2016, 34, 42–63. [Google Scholar] [CrossRef]
- Sperr, W.R. A critical view of root canal therapy. Osterr. Z. Stomatol. 1970, 67, 249–254. [Google Scholar] [PubMed]
- Weiger, R.; Hitzler, S.; Hermle, G.; Löst, C. Periapical status, quality of root canal fillings and estimated endodontic treatment needs in an urban German population. Endod. Dent. Traumatol. 1997, 13, 69–74. [Google Scholar] [CrossRef]
- Bryce, G.; Ready, D.; Donnell, D.O.; Ng, P.; Pratten, J.; Gulabivala, K. Biofilm disruption by root canal irrigants and potential irrigants. Int. Endod. J. 2008, 41, 814–815. [Google Scholar] [CrossRef]
- De Moor, R.; Hülsmann, M.; Kirkevang, L.L.; Tanalp, J.; Whitworth, J. Undergraduate Curriculum Guidelines for Endodontology. Int. Endod. J. 2013, 46, 1105–1114. [Google Scholar] [CrossRef]
- Vieira, A.R.; Siqueira, J.F., Jr.; Ricucci, D.; Lopes, W.S. Dentinal tubule infection as the cause of recurrent disease and late endodontic treatment failure: A case report. J. Endod. 2012, 38, 250–254. [Google Scholar] [CrossRef]
- Ricucci, D.; Loghin, S.; Siqueira, J.F., Jr. Exuberant Biofilm infection in a lateral canal as the cause of short-term endodontic treatment failure: Report of a case. J. Endod. 2013, 39, 712–718. [Google Scholar] [CrossRef]
- Laplace, J.M.; Thuault, M.; Hartke, A.; Boutibonnes, P.; Auffray, Y. Sodium hypochlorite stress in Enterococcus faecalis: Influence of antecedent growth conditions and induced proteins. Curr. Microbiol. 1997, 34, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Peters, O.A. Current challenges and concepts in the preparation of root canal systems: A review. J. Endod. 2004, 30, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.K.; Wesselink, P.R. A primary observation on the preparation and obturation of oval canals. Int. Endod. J. 2001, 34, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Rôças, I.N.; Siqueira, J.F., Jr.; Santos, K.R. Association of Enterococcus faecalis with different forms of periradicular diseases. J. Endod. 2004, 30, 315–320. [Google Scholar] [CrossRef]
- Stuart, C.H.; Schwartz, S.A.; Beeson, T.J.; Owatz, C.B. Enterococcus faecalis: Its role in root canal treatment failure and current concepts in retreatment. J. Endod. 2006, 32, 93–98. [Google Scholar] [CrossRef]
- Narayanan, L.L.; Vaishnavi, C. Endodontic microbiology. J. Conserv. Dent. 2010, 13, 233–239. [Google Scholar] [CrossRef]
- Ran, S.; Liu, B.; Jiang, W.; Sun, Z.; Liang, J. Transcriptome analysis of Enterococcus faecalis in response to alkaline stress. Front. Microbiol. 2015, 6, 795. [Google Scholar] [CrossRef]
- Nehme, F.; Gitau, J.; Liu, J. Purulent pericarditis as a complication of bacteraemic Enterococcus faecalis urinary tract infection. BMJ Case Rep. 2017, 2017, bcr2017219498. [Google Scholar] [CrossRef]
- Lin, D.P.; Wada, S.; Jimenez-Lucho, V.E. Enterococcus faecalis endocarditis presenting as meningitis. Infection 1998, 26, 304–305. [Google Scholar] [CrossRef]
- Ehrlich, G.D.; Hu, F.Z.; Sotereanos, N.; Sewicke, J.; Parvizi, J.; Nara, P.L.; Arciola, C.R. What role do periodontal pathogens play in osteoarthritis and periprosthetic joint infections of the knee? J. Appl. Biomater. Funct. Mater. 2014, 12, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Fujita, N. Vancomycin-resistant enterococci (VRE). Rinsho Byori 2000, 48, 1036–1043. [Google Scholar]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–433. [Google Scholar] [CrossRef]
- Khalifa, L.; Shlezinger, M.; Beyth, S.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Phage therapy against Enterococcus faecalis in dental root canals. J. Oral. Microbiol. 2016, 8, 32157. [Google Scholar] [CrossRef] [PubMed]
- Bolocan, A.S.; Upadrasta, A.; Bettio, P.H.A.; Clooney, A.G.; Draper, L.A.; Ross, R.P.; Hill, C. Evaluation of Phage Therapy in the Context of Enterococcus faecalis and Its Associated Diseases. Viruses 2019, 11, 366. [Google Scholar] [CrossRef] [PubMed]
- González-Moreno, M.P.; Morovic, P.; Tkhilaishvili, T.; Trampuz, A. Bacteriophages for Treatment of Biofilm Infections. In Bone and Joint Infections: From Microbiology to Diagnostics and Treatment; Wiley: Hoboken, NJ, USA, 2021. [Google Scholar]
- Rasband, W.S. ImageJ; U.S. National Institutes of Health: Bethesda, MD, USA, 1997–2018. Available online: https://imagej.nih.gov/ij/ (accessed on 6 June 2022).
- Wang, L.; Tkhilaishvili, T.; Bernal Andres, B.; Trampuz, A.; Gonzalez Moreno, M. Bacteriophage-antibiotic combinations against ciprofloxacin/ceftriaxone-resistant Escherichia coli in vitro and in an experimental Galleria mellonella model. Int. J. Antimicrob. Agents 2020, 56, 106200. [Google Scholar] [CrossRef]
- Tkhilaishvili, T.; Di Luca, M.; Abbandonato, G.; Maiolo, E.M.; Klatt, A.B.; Reuter, M.; Möncke-Buchner, E.; Trampuz, A. Real-time assessment of bacteriophage T3-derived antimicrobial activity against planktonic and biofilm-embedded Escherichia coli by isothermal microcalorimetry. Res. Microbiol. 2018, 169, 515–521. [Google Scholar] [CrossRef]
- RStudio-Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 5 April 2022).
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th Edition. CLSI Document M100, Table 2D. 2020. Available online: https://www.nih.org.pk/wp-content/uploads/2021/02/CLSI-2020.pdf (accessed on 29 March 2022).
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 29 March 2022).
- Sjogren, U.; Hagglund, B.; Sundqvist, G.; Wing, K. Factors affecting the long-term results of endodontic treatment. J. Endod. 1990, 16, 498–504. [Google Scholar] [CrossRef]
- Ricucci, D.; Russo, J.; Rutberg, M.; Burleson, J.A.; Spångberg, L.S. A prospective cohort study of endodontic treatments of 1369 root canals: Results after 5 years. Oral. Surg. Oral. Med. Oral. Pathol. Oral. Radiol. Endod. 2011, 112, 825–842. [Google Scholar] [CrossRef]
- Ricucci, D.; Siqueira, J.F., Jr. Biofilms and apical periodontitis: Study of prevalence and association with clinical and histopathologic findings. J. Endod. 2010, 36, 1277–1288. [Google Scholar] [CrossRef]
- El-Telbany, M.; El-Didamony, G.; Askora, A.; Ariny, E.; Abdallah, D.; Connerton, I.F.; El-Shibiny, A. Bacteriophages to Control Multi-Drug Resistant Enterococcus faecalis Infection of Dental Root Canals. Microorganisms 2021, 9, 517. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Ma, C.; Yin, S.; Song, F.; Qin, K.; Ding, Y.; Yang, X.; Song, P.; Ji, X.; Wei, Y. Phage therapy for refractory periapical periodontitis caused by Enterococcus faecalis in vitro and in vivo. Appl. Microbiol. Biotechnol. 2022, 106, 2121–2131. [Google Scholar] [CrossRef]
- Oliva, A.; Furustrand Tafin, U.; Maiolo, E.M.; Jeddari, S.; Bétrisey, B.; Trampuz, A. Activities of Fosfomycin and Rifampin on Planktonic and Adherent Enterococcus faecalis Strains in an Experimental Foreign-Body Infection Model. Antimicrob. Agents Chemother. 2014, 58, 1284–1293. [Google Scholar] [CrossRef]
- El Haddad, L.; Harb, C.P.; Gebara, M.A.; Stibich, M.A.; Chemaly, R.F. A Systematic and Critical Review of Bacteriophage Therapy Against Multidrug-resistant ESKAPE Organisms in Humans. Clin. Infect. Dis. 2018, 69, 167–178. [Google Scholar] [CrossRef]
- Duerkop, B.A.; Huo, W.; Bhardwaj, P.; Palmer, K.L.; Hooper, L.V. Molecular Basis for Lytic Bacteriophage Resistance in Enterococci. mBio 2016, 7, e01304-16. [Google Scholar] [CrossRef]
- Chatterjee, A.; Johnson, C.N.; Luong, P.; Hullahalli, K.; McBride, S.W.; Schubert, A.M.; Palmer, K.L.; Carlson, P.E., Jr.; Duerkop, B.A. Bacteriophage Resistance Alters Antibiotic-Mediated Intestinal Expansion of Enterococci. Infect. Immun. 2019, 87, e00085-00019. [Google Scholar] [CrossRef] [PubMed]
- Tkhilaishvili, T.; Wang, L.; Perka, C.; Trampuz, A.; Moreno, M.G. Using Bacteriophages as a Trojan Horse to the Killing of Dual-Species Biofilm Formed by Pseudomonas aeruginosa and Methicillin Resistant Staphylococcus aureus. Front. Microbiol. 2020, 11, 695. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, D.; Taha, M.; Yi, Q.; Ramirez-Arcos, S.; Diallo, J.S.; Carli, A.; Abdelbary, H. Does Treatment Order Matter? Investigating the Ability of Bacteriophage to Augment Antibiotic Activity against Staphylococcus aureus Biofilms. Front. Microbiol. 2018, 9, 127. [Google Scholar] [CrossRef]
- Chaudhry, W.N.; Concepción-Acevedo, J.; Park, T.; Andleeb, S.; Bull, J.J.; Levin, B.R. Synergy and Order Effects of Antibiotics and Phages in Killing Pseudomonas aeruginosa Biofilms. PLoS ONE 2017, 12, e0168615. [Google Scholar] [CrossRef]
- Lebeaux, D.; Leflon-Guibout, V.; Ghigo, J.-M.; Beloin, C. In vitro activity of gentamicin, vancomycin or amikacin combined with EDTA or l-arginine as lock therapy against a wide spectrum of biofilm-forming clinical strains isolated from catheter-related infections. J. Antimicrob. Chemother. 2015, 70, 1704–1712. [Google Scholar] [CrossRef]
- Tafin, U.F.; Majic, I.; Belkhodja, C.Z.; Betrisey, B.; Corvec, S.; Zimmerli, W.; Trampuz, A. Gentamicin Improves the Activities of Daptomycin and Vancomycin against Enterococcus faecalis In Vitro and in an Experimental Foreign-Body Infection Model. Antimicrob. Agents Chemother. 2011, 55, 4821–4827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cetinkaya, Y.; Falk, P.; Mayhall, C.G. Vancomycin-resistant enterococci. Clin. Microbiol. Rev. 2000, 13, 686–707. [Google Scholar] [CrossRef] [PubMed]
- Markwart, R.; Willrich, N.; Haller, S.; Noll, I.; Koppe, U.; Werner, G.; Eckmanns, T.; Reuss, A. The rise in vancomycin-resistant Enterococcus faecium in Germany: Data from the German Antimicrobial Resistance Surveillance (ARS). Antimicrob. Resist. Infect. Control. 2019, 8, 147. [Google Scholar] [CrossRef] [PubMed]
- Machado, F.P.; Khoury, R.D.; Toia, C.C.; Orozco, E.I.F.; de Oliveira, F.E.; de Oliveira, L.D.; da Rosa Cardoso, F.G.; Valera, M.C. Primary versus post-treatment apical periodontitis: Microbial composition, lipopolysaccharides and lipoteichoic acid levels, signs and symptoms. Clin. Oral Investig. 2020, 24, 3169–3179. [Google Scholar] [CrossRef] [PubMed]
- Endo, M.S.; Ferraz, C.C.R.; Zaia, A.A.; Almeida, J.F.A.; Gomes, B. Quantitative and qualitative analysis of microorganisms in root-filled teeth with persistent infection: Monitoring of the endodontic retreatment. Eur J. Dent. 2013, 7, 302–309. [Google Scholar] [CrossRef]
- Schirrmeister, J.F.; Liebenow, A.L.; Pelz, K.; Wittmer, A.; Serr, A.; Hellwig, E.; Al-Ahmad, A. New bacterial compositions in root-filled teeth with periradicular lesions. J. Endod. 2009, 35, 169–174. [Google Scholar] [CrossRef]
- Rôças, I.N.; Siqueira, J.F., Jr. Characterization of microbiota of root canal-treated teeth with posttreatment disease. J. Clin. Microbiol. 2012, 50, 1721–1724. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | MIC EF 03 | MIC EF 06 | MIC EF 10 | MIC EF 14 | MIC EF 04 |
|---|---|---|---|---|---|
| Vancomycin | 2 µg/mL | 2 µg/mL | 2 µg/mL | 2 µg/mL | - |
| Gentamicin | 64 µg/mL | 64 µg/mL | 128 µg/mL | 64 µg/mL | - |
| Fosfomycin | - | - | - | - | 32 µg/mL |
| Daptomycin | - | - | - | - | 8 µg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voit, M.; Trampuz, A.; Gonzalez Moreno, M. In Vitro Evaluation of Five Newly Isolated Bacteriophages against E. faecalis Biofilm for Their Potential Use against Post-Treatment Apical Periodontitis. Pharmaceutics 2022, 14, 1779. https://doi.org/10.3390/pharmaceutics14091779
Voit M, Trampuz A, Gonzalez Moreno M. In Vitro Evaluation of Five Newly Isolated Bacteriophages against E. faecalis Biofilm for Their Potential Use against Post-Treatment Apical Periodontitis. Pharmaceutics. 2022; 14(9):1779. https://doi.org/10.3390/pharmaceutics14091779
Chicago/Turabian StyleVoit, Marie, Andrej Trampuz, and Mercedes Gonzalez Moreno. 2022. "In Vitro Evaluation of Five Newly Isolated Bacteriophages against E. faecalis Biofilm for Their Potential Use against Post-Treatment Apical Periodontitis" Pharmaceutics 14, no. 9: 1779. https://doi.org/10.3390/pharmaceutics14091779
APA StyleVoit, M., Trampuz, A., & Gonzalez Moreno, M. (2022). In Vitro Evaluation of Five Newly Isolated Bacteriophages against E. faecalis Biofilm for Their Potential Use against Post-Treatment Apical Periodontitis. Pharmaceutics, 14(9), 1779. https://doi.org/10.3390/pharmaceutics14091779