

Inhibitory Effect of Periodontitis through C/EBP and 11β-Hydroxysteroid Dehydrogenase Type 1 Regulation of Betulin Isolated from the Bark of Betula platyphylla
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plants Materials
2.3. Extraction and Isolation
2.4. Protein Analysis Using Western Blot Analysis
2.5. Analysis of Gene Level Using RT-qPCR
2.6. Mineralization Assay
2.7. Periodontitis Induction Model Using PG-LPS
2.8. Analysis of Periodontal Tissue Using Micro-CT Images
2.9. Histological Staining
2.10. Goldner’s Masson Trichrome Staining
2.11. ELISA Assays
2.12. Statistical Analysis
3. Results
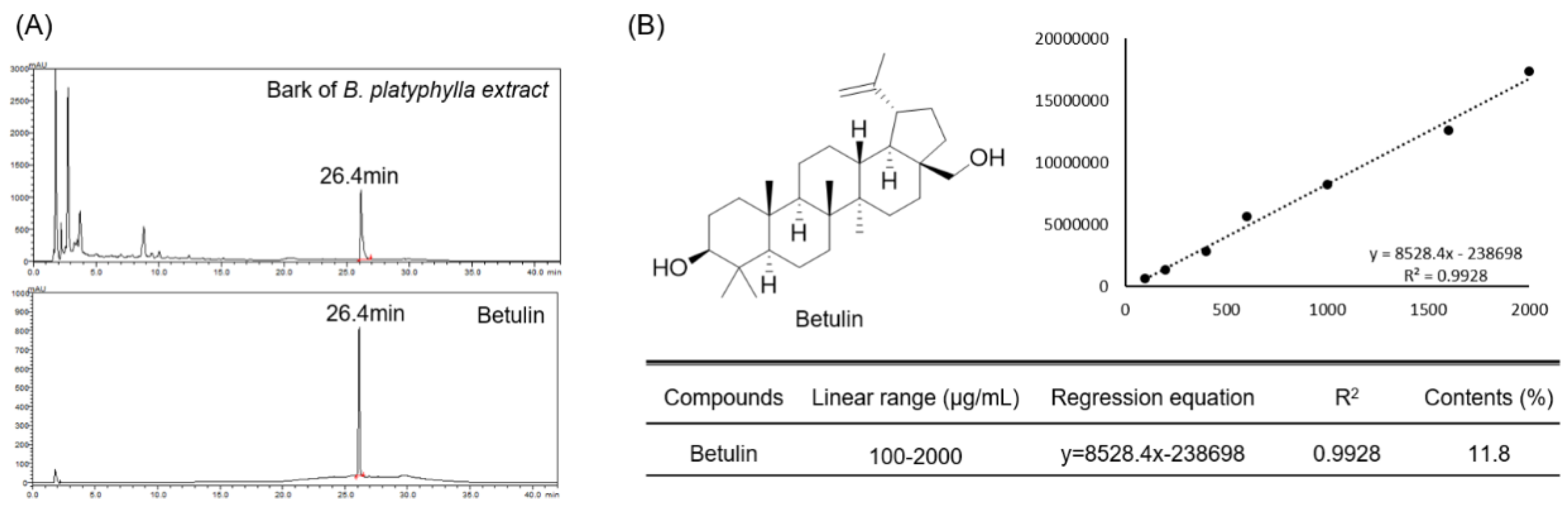
3.1. Quantitative Analysis of Betulin in Betula platyphylla Bark
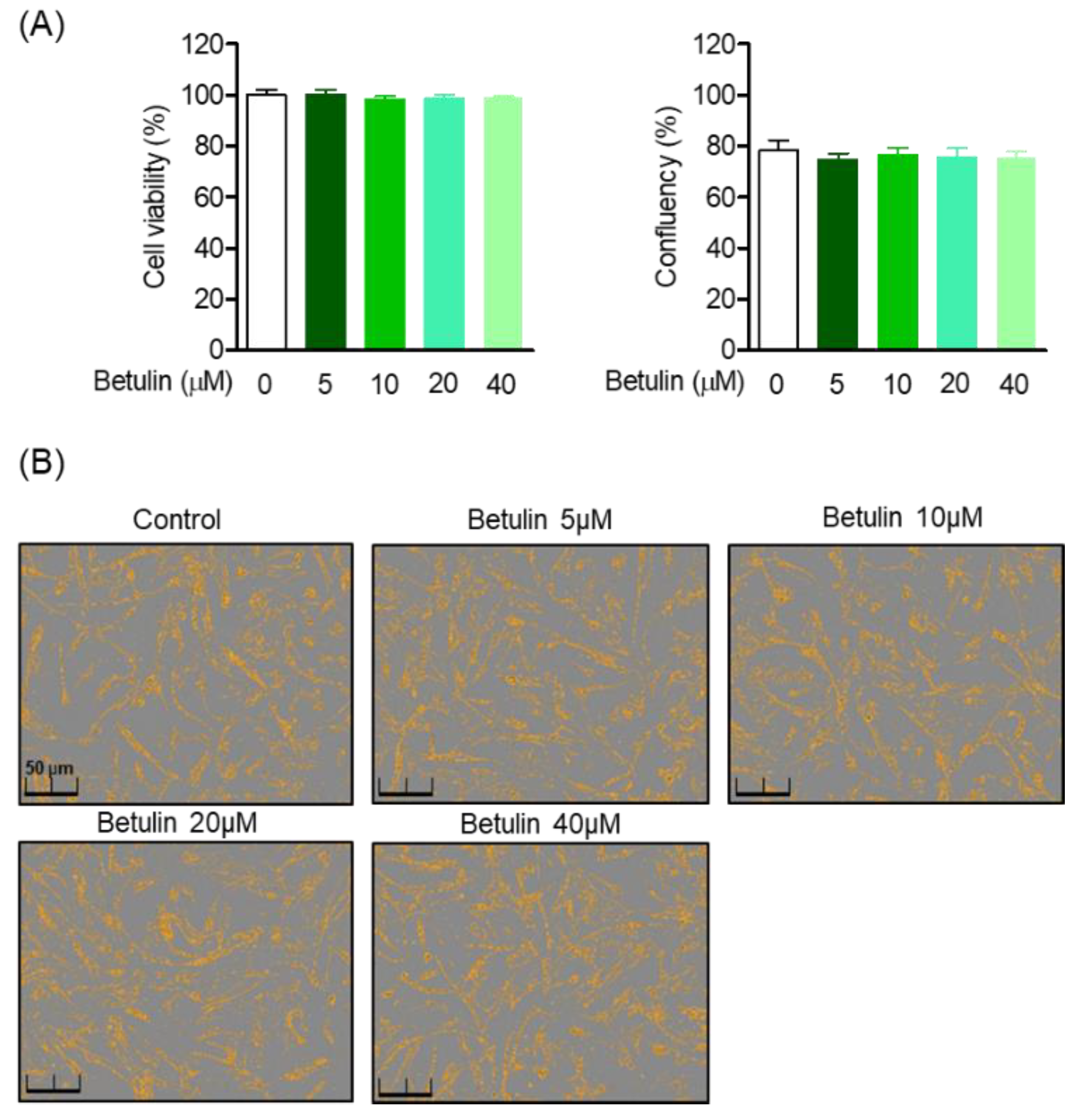
3.2. Effect of Betulin on Human Periodontal Ligament (HPDL) Cell Viability and Confluence
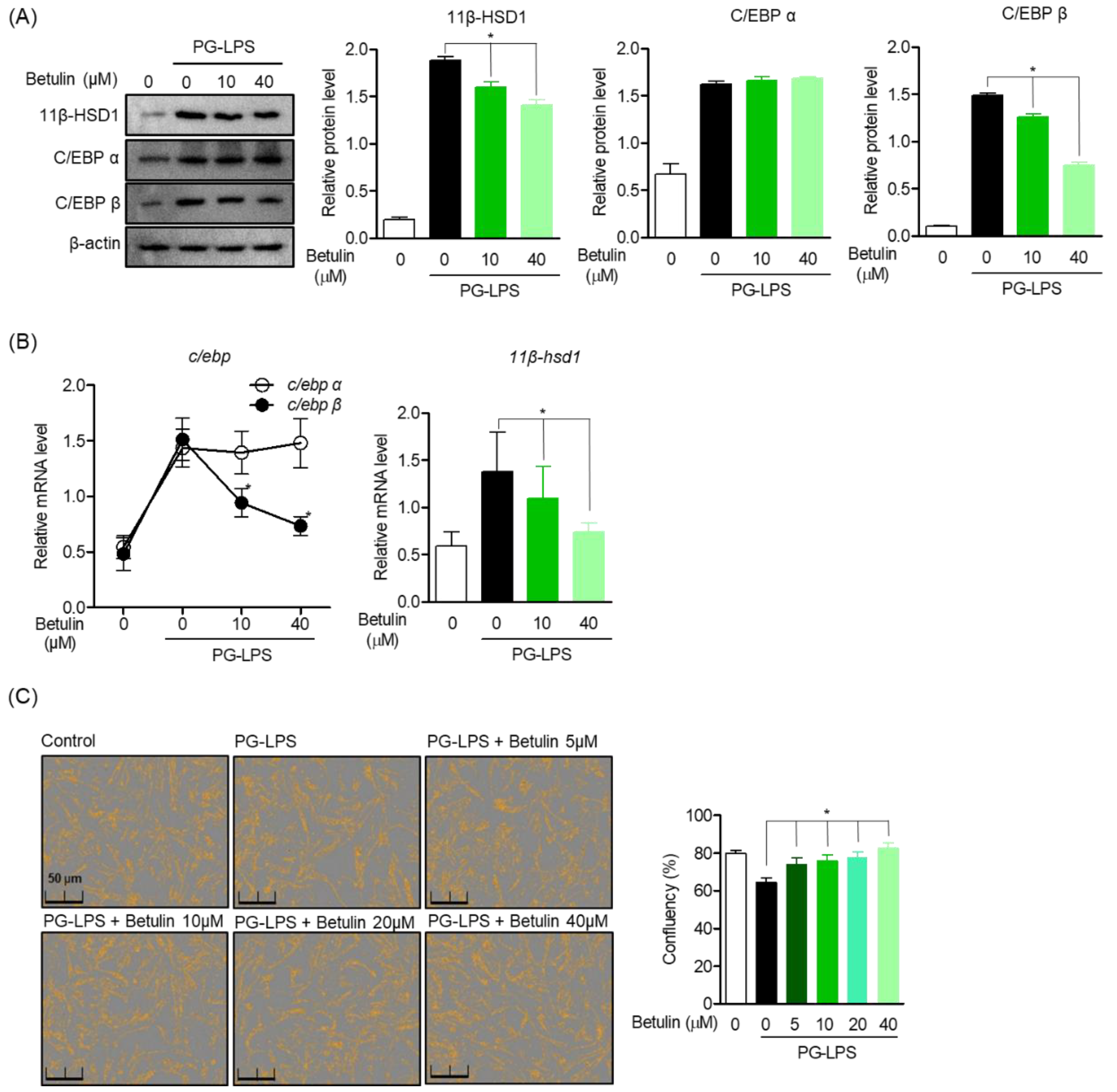
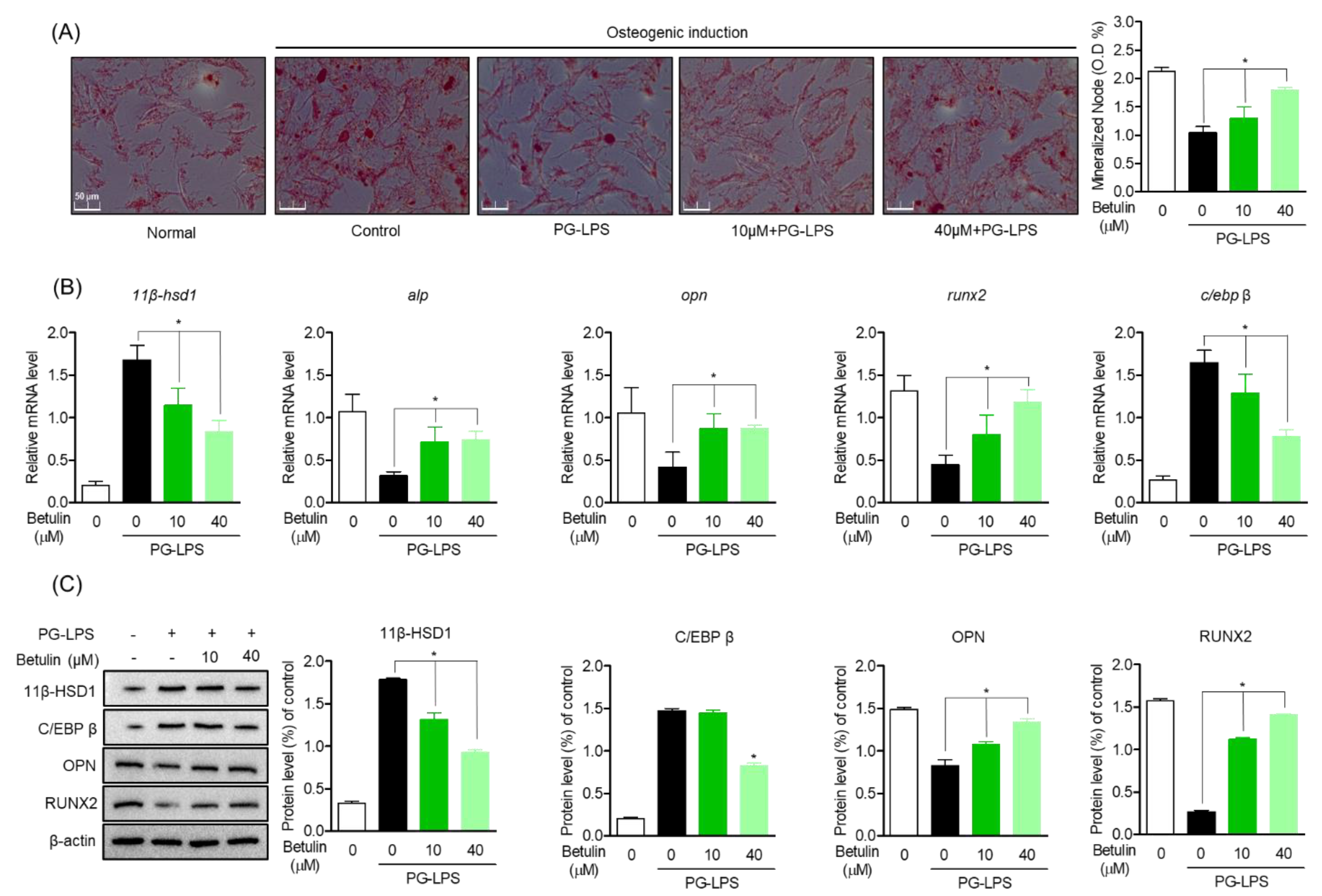
3.3. Effect of Betulin on C/EBP and 11β-HSD1 Activity Regulation in PG-LPS Stimulated HPDL Cells
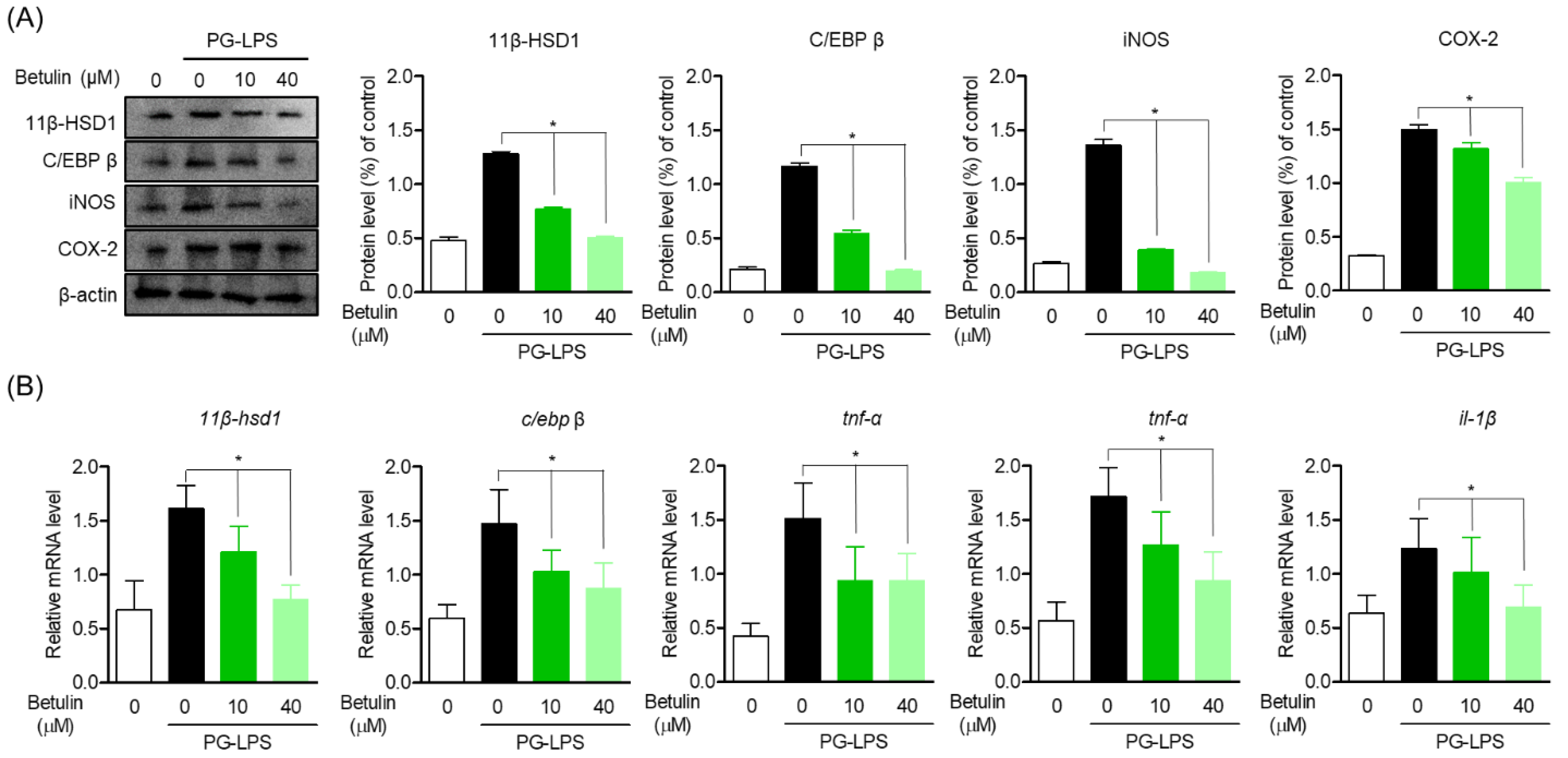
3.4. Inhibitory Effect of Betulin on Inflammatory Mediators by Regulating 11β-HSD1 and C/EBP Activities
3.5. Induction Effect of Betulin on Osteoblast Differentiation through HSD Activity Regulation
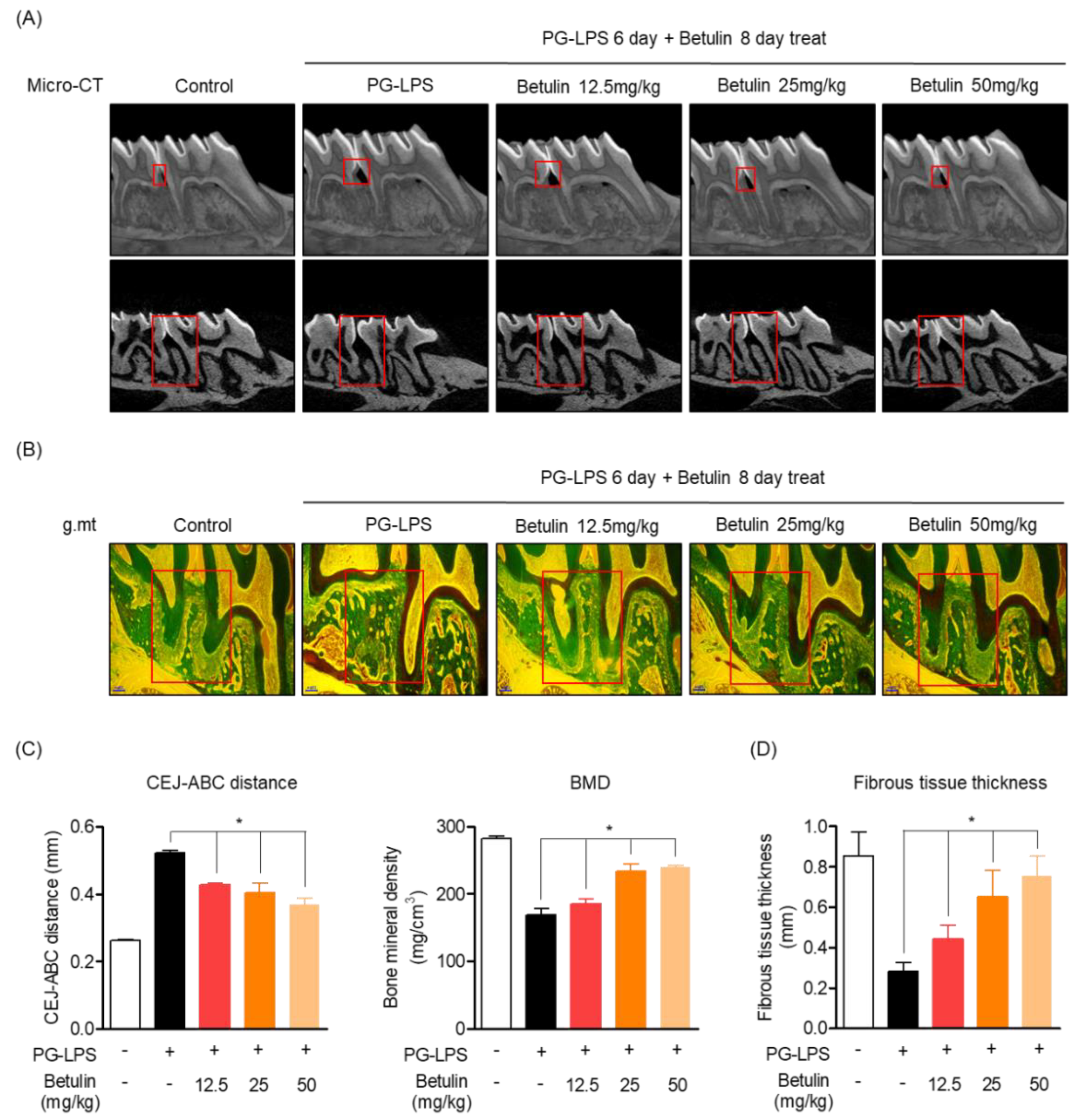
3.6. Protective Effect of Betulin on Alveolar Bone and Fibrous Tissue Lost with PG-LPS in a Periodontitis-Induced In Vivo Model
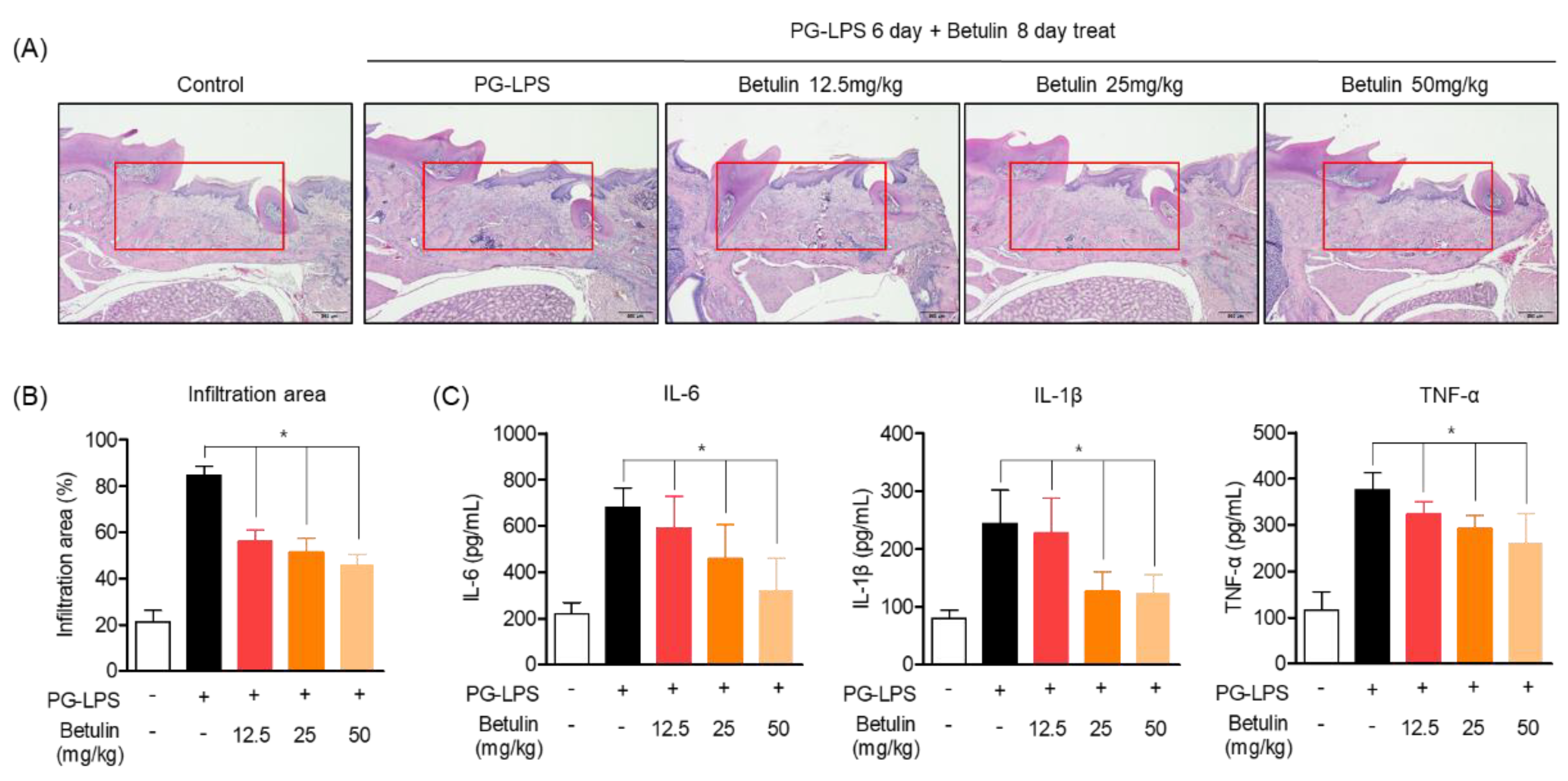
3.7. Inhibitory Effect of Betulin on Periodontal Tissue Infiltration in PG-LPS-Induced Periodontitis In Vivo Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Listyarifah, D.; Al-Samadi, A.; Salem, A.; Syaify, A.; Salo, T.; Tervahartiala, T.; Ainola, M. Infection and apoptosis associated with inflammation in periodontitis: An immunohistologic study. Oral Dis. 2017, 23, 1144–1154. [Google Scholar] [CrossRef]
- Jorgen, S.; Casey, C. The oral microflora and human periodontal disease. In Medical Importance of the Normal Microflora; Springer: New York, NY, USA, 1999; pp. 102–127. [Google Scholar] [CrossRef]
- Seckl, J.R.; Walker, B.R. Minireview: 11β-Hydroxysteroid DehydrogenaseType 1—A Tissue-Specific Amplifier of Glucocorticoid Action. Endocrinology 2001, 142, 1371–1376. [Google Scholar] [CrossRef]
- Stewart, P.M.; Krozowski, Z.S. 11b-Hydroxysteroid dehydrogenase. Vitam. Horm. 1999, 57, 249–324. [Google Scholar]
- Wang, M. The role of glucocorticoid action in the pathophysiology of the metabolic syndrome. Nutr. Metab. 2005, 2, 3. [Google Scholar] [CrossRef]
- Rask, E.; Olsson, T.; Söderberg, S.; Andrew, R.; Livingstone, D.E.; Johnson, O.; Walker, B.R. Tissue-Specific Dysregulation of Cortisol Metabolism in Human Obesity. J. Clin. Endocrinol. Metab. 2001, 86, 1418–1421. [Google Scholar] [CrossRef]
- Paulsen, S.K.; Pedersen, S.B.; Fisker, S.; Richelsen, B. 11β-HSD Type 1 Expression in Human Adipose Tissue: Impact of Gender, Obesity, and Fat Localization. Obesity 2007, 15, 1954–1960. [Google Scholar] [CrossRef]
- Nuotio-Antar, A.M.; Hachey, D.L.; Hasty, A.H. Carbenoxolone Treatment Attenuates Symptoms of Metabolic Syndrome and Atherogenesis in Obese, Hyperlipidemic Mice. Am. J. Physiol. Endocrinol. Metab. 2007, 293, 1517–1528. [Google Scholar] [CrossRef]
- Rosenstock, J.; Banarer, S.; Fonseca, V.A.; Inzucchi, S.E.; Sun, W.; Yao, W.; Hollis, G.; Flores, R.; Levy, R.; Williams, W.V.; et al. The 11-β-Hydroxysteroid Dehydrogenase Type 1 Inhibitor INCB13739 Improves Hyperglycemia in Patients with Type 2 Diabetes Inadequately Controlled by Metformin Monotherapy. Diabetes Care 2010, 33, 1516–1522. [Google Scholar] [CrossRef]
- Zbánková, S.; Bryndová, J.; Leden, P.; Kment, M.; Svec, A.; Pácha, J. 11β-hydroxysteroid dehydrogenase 1 and 2 expression in colon from patients with ulcerative colitis. J. Gastroenterol. Hepatol. 2007, 22, 1019–1023. [Google Scholar] [CrossRef]
- Hardy, R.; Rabbitt, E.H.; Filer, A.; Emery, P.; Hewison, M.; Stewart, P.M. Local and systemic glucocorticoid metabolism in inflammatory arthritis. Ann. Rheum. Dis. 2008, 67, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Nakata, T.; Fujita, A.; Umeda, M.; Yoshida, H.; Inami, K.; Masuzaki, H. The increased ratio of 11β-hydroxysteroid dehydrogenase type 1 versus 11β-hydroxysteroid dehydrogenase type 2 in chronic periodontitis irrespective of obesity. Springerplus. 2016, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- Seibel, M.J.; Cooper, M.S.; Zhou, H. Glucocorticoid-induced osteoporosis: Mechanisms, management, and future perspectives. Lancet Diabetes Endocrinol. 2013, 1, 59–70. [Google Scholar] [CrossRef]
- Huh, J.E.; Baek, Y.H.; Kim, Y.J.; Lee, J.D.; Choi, D.Y.; Park, D.S. Protective effects of butanol fraction from Betula platyphylla var. japonica on cartilage alterations in a rabbit collagenase-induced osteoarthritis. J. Ethnopharmacol. 2009, 123, 515–521. [Google Scholar] [CrossRef]
- Fuchen, S.; Jingwen, L.; Takayoshi, K.; Shaoquan, N. Resources of the white birch (Betula platyphylla) for sap production and its ecological characteristics in northeast China. Eurasian J. For. Res. 2001, 2, 31–38. [Google Scholar]
- Matsuda, H.; Ishikado, A.; Nishida, N.; Ninomiya, K.; Fujiwara, H.; Kobayashi, Y.; Yoshikawa, M. Hepatoprotective, superoxide scavenging, and antioxidative activities of aromatic constituents from the bark of Betula platyphylla var. japonica. Bioorganic Med. Chem. Lett. 1998, 8, 2939–2944. [Google Scholar] [CrossRef]
- Jiang, H.; Shen, Y.; Yasuda, E.; Chiba, M.; Terazawa, M. Phenolic glucosides from Inner bark of shirakamba birch, Betula platyphylla Sukatchev var. japonica Hara. Eurasian J. For. Res. 2001, 3, 49–54. [Google Scholar]
- Zhang, Z.; Hong, S. Determination of betulin content from bark of Betula platyphylla Suk by HPLC. Chem. Ind. For. Prod. 2004, 24, 61–63. [Google Scholar]
- Zhang, Y.; Yu, T.; Wang, Y. Extraction of botulin from bark of Betula platyphylla by supercritical carbon dioxide extraction. J. For. Res. 2003, 14, 202–204. [Google Scholar] [CrossRef]
- Alakurtti, S.; Makela, T.; Koskimies, S.; Yli-Kauhaluoma, J. Pharmacological properties of the ubiquitous natural product botulin. Eur. J. Pharm. Sci. 2006, 29, 1–13. [Google Scholar] [CrossRef]
- Wu, Q.; Li, H.; Qiu, J.; Feng, H. Betulin protects mice from bacterial pneumonia andacute lung injury. Microb. Pathog. 2014, 75, 21–28. [Google Scholar] [CrossRef]
- Szuster-Ciesielska, A.; Plewka, K.; Daniluk, J.; Kandefer-Szerszen, M. Betulin and betulinic acid attenuate ethanol-induced liver stellate cell activation by inhibiting reactive oxygen species (ROS), cytokine (TNF-alpha, TGF-beta) production and by influencing intracellular signaling. Toxicology 2011, 280, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Song, D.; Wang, T.; Xia, X.; Hu, W.; Han, F. LFP-20, a porcine lactoferrin peptide, ameliorates LPS-induced inflammation via the MyD88/NF-kappaB and MyD88/MAPK signaling pathways. Dev. Comp. Immunol. 2015, 52, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, Y.; Zou, P.; Wang, J.; He, W.; Shi, D. Synthesis and biological evaluation of a novel class of curcumin analogs as anti-inflammatory agents for prevention and treatment of sepsis in mouse model. Drug Des. Dev. Ther. 2015, 9, 1663–1678. [Google Scholar] [CrossRef] [Green Version]
- So, H.M.; Eom, H.J.; Lee, D.; Kim, S.; Kang, K.S.; Lee, K.; Baek, K.H.; Park, J.Y.; Kim, K.H. Bioactivity evaluations of betulin identified from the bark of Betula platyphylla var. japonica for cancer therapy. Arch. Pharmacal Res. 2018, 41, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Ishisaka, A.; Ansai, T.; Soh, I.; Inenaga, K.; Yoshida, A.; Shigeyama, C.; Awano, S.; Hamasaki, T.; Sonoki, K.; Takata, Y.; et al. Association of salivary levels of cortisol and dehydroepiandrosterone with periodontitis in older Japanese adults. J. Periodontol. 2007, 78, 1767–1773. [Google Scholar] [CrossRef]
- Cooper, M.S.; Rabbitt, E.H.; Goddard, P.E.; Bartlett, W.A.; Hewison, M.; Stewart, P.M. Osteoblastic 11beta-hydroxysteroid dehydrogenase type 1 activity increases with age and glucocorticoid exposure. J. Bone Miner. Res. 2002, 17, 979–986. [Google Scholar] [CrossRef]
- Justesen, J.; Mosekilde, L.; Holmes, M.; Stenderup, K.; Gasser, J.; Mullins, J.J.; Seckl, J.R.; Kassem, M. Mice deficient in 11beta-hydroxysteroid dehydrogenase type 1 lack bone marrow adipocytes, but maintain normal bone formation. Endocrinology 2004, 145, 1916–1925. [Google Scholar] [CrossRef]
- Cho, Y.J.; Huh, J.E.; Kim, D.Y.; Kim, N.J.; Lee, J.D.; Baek, Y.H.; Cho, E.M.; Yang, H.R.; Choi, D.Y.; Park, D.S. Effect of Betula platyphylla var. japonica on proteoglycan release, type II collagen degradation, and matrix metalloproteinase expression in rabbit articular cartilage explants. Biol. Pharm. Bull. 2006, 29, 1408–1413. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, Z.; Liu, W.; Han, X.; Zhao, M. Betulin attenuates lung and liver injuries in sepsis. Int. Immunopharmacol. 2016, 30, 50–56. [Google Scholar] [CrossRef]
- Cho, N.K.; Kim, D.H.; Sung, S.H. Simultaneous Determination of Platyphylloside, Aceroside VIII and Betulin in Betula platyphylla bark by HPLC-DAD. Korean J. Pharmacogn. 2014, 45, 294–299. [Google Scholar]
- Park, S.B.; Park, J.S.; Jung, W.H.; Kim, H.Y.; Kwak, H.J.; Ahn, J.H.; Choi, K.J.; Yoon-Ju Na, Y.J.; Choi, S.; Rhee, S.D.; et al. Anti-inflammatory effect of a selective 11b-hydroxysteroid dehydrogenase type 1 inhibitor via the stimulation of heme oxygenase-1 in LPS-activated mice and J774.1 murine macrophages. J. Pharmacol. Sci. 2016, 131, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Assuma, R.; Oates, T.; Cochran, D.; Amar, S.; Graves, D.T. IL-1 and TNF antagonists inhibit the inflammatory response and bone loss in experimental periodontitis. J. Immunol. 1998, 160, 403–409. [Google Scholar] [PubMed]
- Nakata, T.; Makoto, U.; Masuzaki, H.; Sawai, H. The expression of 11β-hydroxysteroid dehydrogenase type 1 is increased in experimental periodontitis in rats. BMC Oral Health 2016, 16, 108. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.E.; Hyun, K.Y. Inhibitory effect of Acer tegmentosum maxim extracts on P. gingivalis LPS induced Periodontitis. Arch. Oral Biol. 2020, 109, 104529. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence (5′→3′) | |
|---|---|---|
| 11β-hsd1 | Forward | GGGGTACCTTTTTCCCCGCTCTACTGATAACT |
| Reverse | TTCTCGAGCCGACAGGGAGCTGGCCTGAAGACT | |
| c/ebp β | Forward | AGAAGACCGTGGACAAGCACAG |
| Reverse | CTCCAGGACCTTGTGCTGCGT | |
| c/ebpα | Forward | CGGACTTGGTGCGTCTAAGATG |
| Reverse | GCATTGGAGCGGTGAGTTTG | |
| il-6 | Forward | AGTGAGGAACAAGCCAGAGC |
| Reverse | GTCAGGGGTGGTTATTGCAT | |
| il-1β | Forward | AACCTCTTCGAGGCACAAGG |
| Reverse | GTCCTGGAAGGAGCACTTCAT | |
| tnf-α | Forward | GCCTCTTCTCCTTCCTGATCGT |
| Reverse | TGAGGGTTTGCTACAACATGGG | |
| alp | Forward | TGCAGTACGAGCTGAACAGG |
| Reverse | GTCAATTCTGCCTCCTTCCA | |
| opn | Forward | TCAGCTGGATGACCAGAGTG |
| Reverse | TTGGGGTCTACAACCAGCAT | |
| runx2 | Forward | TCTTAGAACAAATTCTGCCCTTT |
| Reverse | TGCTTTGGTCTTGAAATCACA | |
| gapdh | Forward | TGTTCGTCATGGGTGTGAAC |
| Reverse | GTCTTCTGGGTGGCAGTGAT | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-N.; Jeong, G.-S. Inhibitory Effect of Periodontitis through C/EBP and 11β-Hydroxysteroid Dehydrogenase Type 1 Regulation of Betulin Isolated from the Bark of Betula platyphylla. Pharmaceutics 2022, 14, 1868. https://doi.org/10.3390/pharmaceutics14091868
Kim E-N, Jeong G-S. Inhibitory Effect of Periodontitis through C/EBP and 11β-Hydroxysteroid Dehydrogenase Type 1 Regulation of Betulin Isolated from the Bark of Betula platyphylla. Pharmaceutics. 2022; 14(9):1868. https://doi.org/10.3390/pharmaceutics14091868
Chicago/Turabian StyleKim, Eun-Nam, and Gil-Saeng Jeong. 2022. "Inhibitory Effect of Periodontitis through C/EBP and 11β-Hydroxysteroid Dehydrogenase Type 1 Regulation of Betulin Isolated from the Bark of Betula platyphylla" Pharmaceutics 14, no. 9: 1868. https://doi.org/10.3390/pharmaceutics14091868
APA StyleKim, E. -N., & Jeong, G. -S. (2022). Inhibitory Effect of Periodontitis through C/EBP and 11β-Hydroxysteroid Dehydrogenase Type 1 Regulation of Betulin Isolated from the Bark of Betula platyphylla. Pharmaceutics, 14(9), 1868. https://doi.org/10.3390/pharmaceutics14091868