
Neonatal Morphine Results in Long-Lasting Alterations to the Gut Microbiome in Adolescence and Adulthood in a Murine Model

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Experimental Design
2.3. 16S rRNA Gene Sequencing
2.4. Bioinformatic Analysis
2.5. Statistical Analysis
3. Results
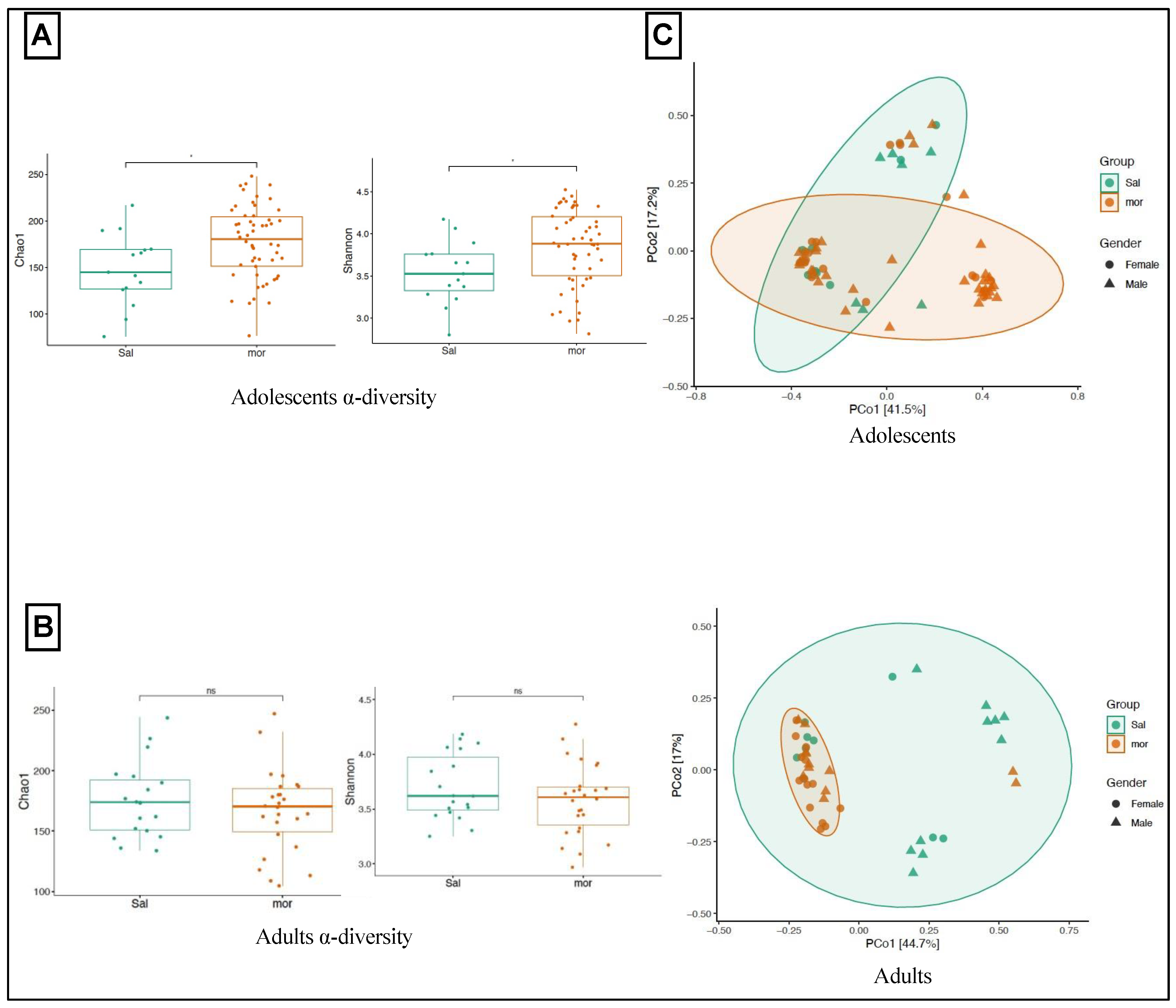
3.1. Impact of Neonatal Morphine Exposure on Gut Microbial Diversity
3.1.1. In Adolescence
3.1.2. In Adulthood
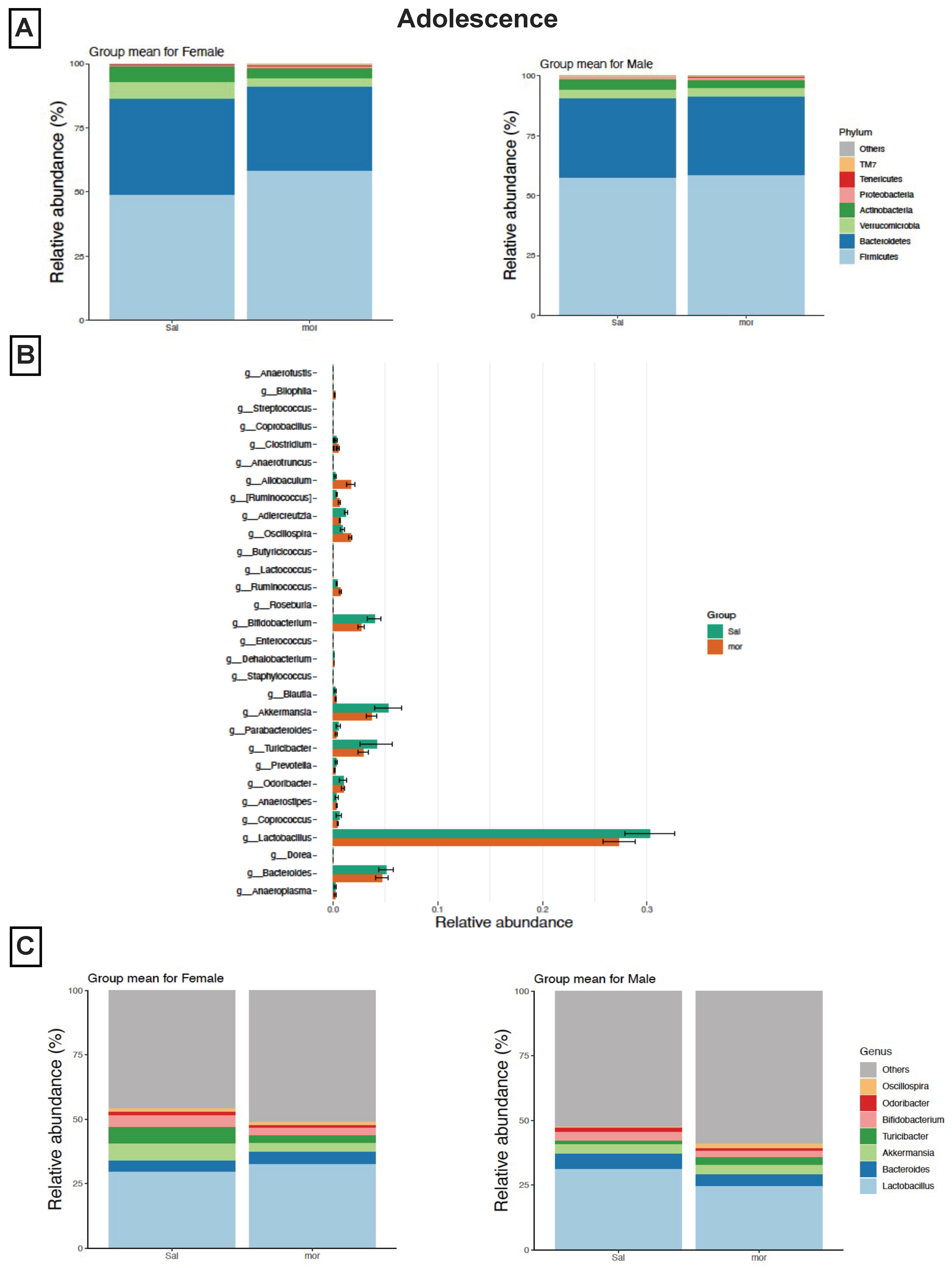
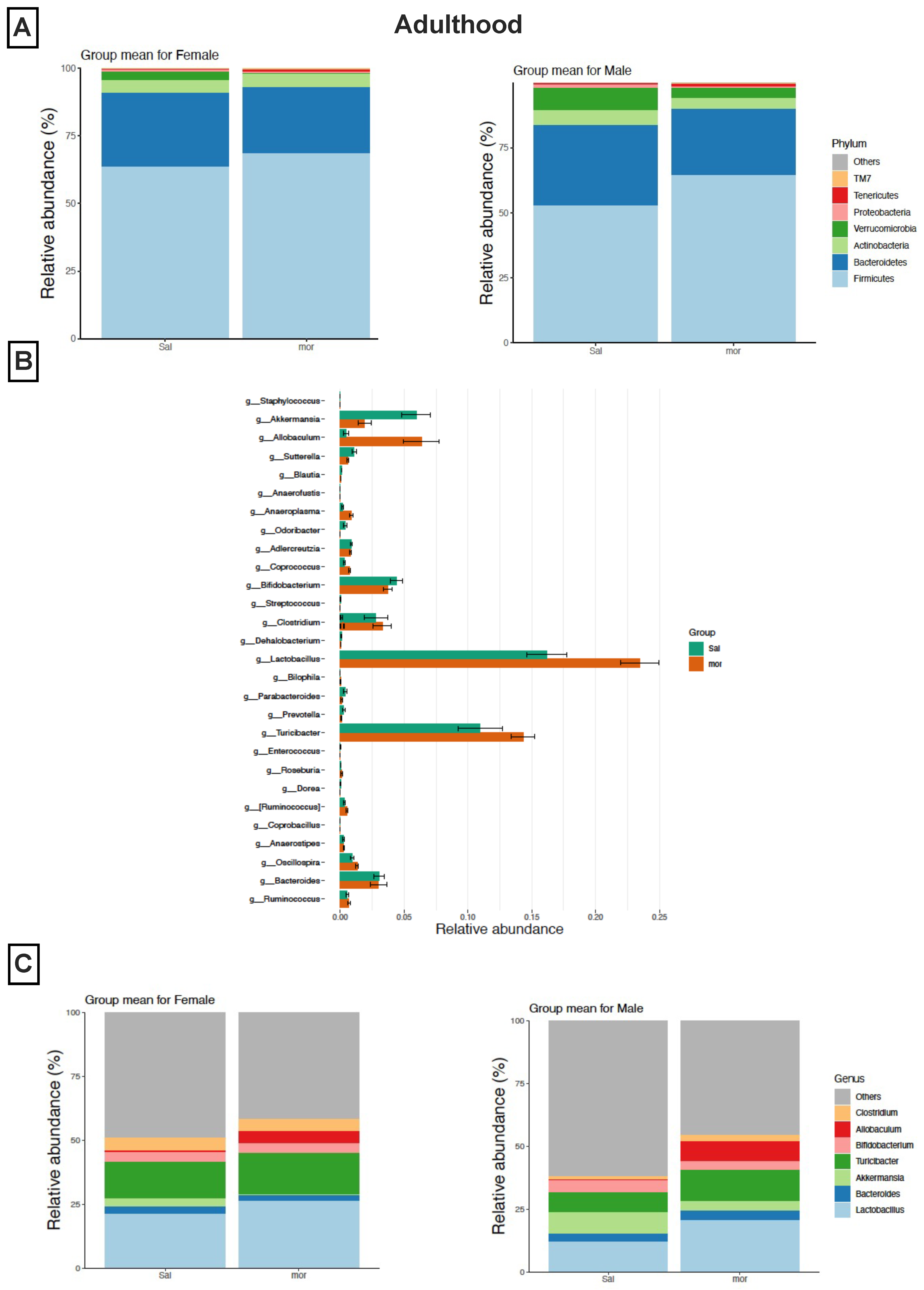
3.2. Impact of Neonatal Morphine Exposure on Gut Microbial Composition
3.2.1. In Adolescence
3.2.2. In Adulthood
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dowell, D.; Haegerich, T.M.; Chou, R. CDC guideline for prescribing opioids for chronic pain-United States, 2016. JAMA—J. Am. Med. Assoc. 2016, 315, 1624–1645. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Preterm Birth|Maternal and Infant Health|Reproductive Health|CDC. In Centers for Disease Control and Prevention; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018. [Google Scholar]
- Hall, R.W.; Anand, K.J.S. Pain management in newborns. Clin. Perinatol. 2014, 41, 895–924. [Google Scholar] [CrossRef] [PubMed]
- Ricardo Buenaventura, M.; Rajive Adlaka, M.; Nalini Sehgal, M. Opioid complications and side effects. Pain Physician 2008, 11, S105–S120. [Google Scholar] [CrossRef]
- Human Microbiome Project Consortium. Structure, Function and Diversity of the Healthy Human Microbiome The Human Microbiome Project Consortium HHS Public Access. Nature 2013, 486, 207–214. [Google Scholar]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Chen, L.H.; Xing, C.; Liu, T. Pain regulation by gut microbiota: Molecular mechanisms and therapeutic potential. Br. J. Anaesth. 2019, 123, 637–654. [Google Scholar] [CrossRef]
- Pacifici, G.M.; Säwe, J.; Kager, L.; Rane, A. Morphine glucuronidation in human fetal and adult liver. Eur. J. Clin. Pharmacol. 1982, 22, 553–558. [Google Scholar] [CrossRef]
- Wang, F.; Roy, S. Gut Homeostasis, Microbial Dysbiosis, and Opioids. Toxicol. Pathol. 2017, 45, 150–156. [Google Scholar] [CrossRef]
- Degruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current understanding of dysbiosis in disease in human and animal models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef]
- Meng, J.; Yu, H.; Ma, J.; Wang, J.; Banerjee, S.; Charboneau, R.; Barke, R.A.; Roy, S. Morphine Induces Bacterial Translocation in Mice by Compromising Intestinal Barrier Function in a TLR-Dependent Manner. PLoS ONE 2013, 8, e54040. [Google Scholar] [CrossRef]
- Wang, F.; Meng, J.; Zhang, L.; Johnson, T.; Chen, C.; Roy, S. Morphine induces changes in the gut microbiome and metabolome in a morphine dependence model. Sci. Rep. 2018, 8, 3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilburger, M.E.; Adler, M.W.; Truant, A.L.; Meissler, J.J.J.; Satishchandran, V.; Rogers, T.J.; Eisenstein, T.K. Morphine induces sepsis in mice. J. Infect. Dis. 1997, 176, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Amaral, F.A.; Sachs, D.; Costa, V.V.; Fagundes, C.T.; Cisalpino, D.; Cunha, T.M.; Ferreira, S.H.; Cunha, F.Q.; Silva, T.A.; Nicoli, J.R.; et al. Commensal microbiota is fundamental for the development of inflammatory pain. Proc. Natl. Acad. Sci. USA 2008, 105, 2193–2197. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, S.; Felice, V.; Nally, K.; Savignac, H.; Claesson, M.; Scully, P.; Woznicki, J.; Hyland, N.; Shanahan, F.; Quigley, E.; et al. Disturbance of the gut microbiota in early-life selectively affects visceral pain in adulthood without impacting cognitive or anxiety-related behaviors in male rats. Neuroscience 2014, 277, 885–901. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Wang, Y.; Zhang, P.; Yuan, Y.; Zhang, Y.; Chen, G. Gut microbiota regulates neuropathic pain: Potential mechanisms and therapeutic strategy. J. Headache Pain 2020, 21, 103. [Google Scholar] [CrossRef]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef]
- Zhang, G.H.; Sweitzer, S.M. Neonatal morphine enhances nociception and decreases analgesia in young rats. Brain Res. 2008, 1199, 82–90. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nature Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- De Wit, N.; Derrien, M.; Bosch-Vermeulen, H.; Oosterink, E.; Keshtkar, S.; Duval, C.; de Vogel-van den Bosch, J.; Kleerebezem, M.; Müller, M.; Van Der Meer, R. Saturated fat stimulates obesity and hepatic steatosis and affects gut microbiota composition by an enhanced overflow of dietary fat to the distal intestine. Am. J. Physiol. Gastrointest Liver Physiol. 2012, 303, G589–G599. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Grecco, G.G.; Gao, Y.; Gao, H.; Liu, Y.; Atwood, B.K. Prenatal opioid administration induces shared alterations to the maternal and offspring gut microbiome: A preliminary analysis. Drug Alcohol Depend. 2021, 227, 108914. [Google Scholar] [CrossRef]
- Abu, Y.; Tao, J.; Dutta, R.; Yan, Y.; Vitari, N.; Kolli, U.; Roy, S. Brief Hydromorphone Exposure During Pregnancy Sufficient to Induce Maternal and Neonatal Microbial Dysbiosis. J. Neuroimmune Pharmacol. 2021, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Menni, C.; Jackson, M.A.; Pallister, T.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Gut microbiome diversity and high-fibre intake are related to lower long-term weight gain. Int. J. Obes. 2017, 41, 1099–1105. [Google Scholar] [CrossRef]
- Banerjee, S.; Sindberg, G.; Wang, F.; Meng, J.; Sharma, U.; Zhang, L.; Dauer, P.; Chen, C.; Dalluge, J.; Johnson, T.; et al. Opioid-induced gut microbial disruption and bile dysregulation leads to gut barrier compromise and sustained systemic inflammation. Mucosal. Immunol. 2016, 9, 1418–1428. [Google Scholar] [CrossRef]
- Xuejun, L.; Xie, Z.; Wang, H.; Shen, Z.; Guo, Y.; Gao, Y.; Chen, X.; Wu, Q.; Li, X.; Wang, K. Bacterial Diversity of Intestinal Microbiota in Patients with Substance Use Disorders Revealed by 16S rRNA Gene Deep Sequencing. Sci. Rep. 2017, 7, 3628. [Google Scholar] [CrossRef]
- Ren, M.; Lotfipour, S. Dose- and Sex-Dependent Bidirectional Relationship between Intravenous Fentanyl Self-Administration and Gut Microbiota. Microorganisms 2022, 10, 1127. [Google Scholar] [CrossRef]
- Kullberg, R.F.J.; Wiersinga, W.J.; Haak, B.W. Gut microbiota and sepsis: From pathogenesis to novel treatments. Curr. Opin. Gastroenterol. 2021, 37, 578–585. [Google Scholar] [CrossRef]
- Abu, Y.; Vitari, N.; Yan, Y.; Roy, S. Opioids and Sepsis: Elucidating the Role of the Microbiome and microRNA-146. Int. J. Mol. Sci. 2022, 23, 1097. [Google Scholar] [CrossRef]
- Bassetti, M.; Bandera, A.; Gori, A. Therapeutic Potential of the Gut Microbiota in the Management of Sepsis. Crit. Care 2020, 24, 105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Li, C.; Shen, L.; Wang, S.; Li, X.; Fu, C.; Li, T.; Liu, B.; Gu, Y.; Wang, W.; et al. The Intestinal Effect of Atorvastatin: Akkermansia muciniphila and Barrier Function. Front. Microbiol. 2022, 12, 797062. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, S.; Abbasi, A.; Somi, M.H.; Moaddab, S.Y.; Nikniaz, L.; Kafil, H.S.; Leylabadlo, H.E. Akkermansia muciniphila: From its critical role in human health to strategies for promoting its abundance in human gut microbiome. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kang, S.W.; Lee, J.H.; Park, S.H.; Lee, J.S. The evolution and competitive strategies of Akkermansia muciniphila in gut. Gut Microbes 2022, 14, 1. [Google Scholar] [CrossRef]
- Claesson, M.J.; van Sinderen, D.; O’Toole, P.W. The genus Lactobacillus—A genomic basis for understanding its diversity. FEMS Microbiol. Lett. 2007, 269, 22–28. [Google Scholar] [CrossRef]
- Sanders, M.E. Probiotics: Considerations for human health. Nutr. Rev. 2003, 61, 91–99. [Google Scholar] [CrossRef]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef]
- Handelmann, G.E.; Dow-Edwards, D. Modulation of brain development by morphine: Effects on central motor systems and behavior. Peptides 1985, 6, 29–34. [Google Scholar] [CrossRef]
- Borrelli, K.N.; Yao, E.J.; Yen, W.W.; Phadke, R.A.; Ruan, Q.T.; Chen, M.M.; Kelliher, J.C.; Langan, C.R.; Scotellaro, J.L.; Babbs, R.K.; et al. Sex differences in behavioral and brainstem transcriptomic neuroadaptations following neonatal opioid exposure in outbred mice. eNeuro 2021, 8, 5. [Google Scholar] [CrossRef]
- Craig, M.M.; Bajic, D. Long-term behavioral effects in a rat model of prolonged postnatal morphine exposure. Behav. Neurosci. 2015, 129, 643–655. [Google Scholar] [CrossRef]
- Sultan, S.; El-Mowafy, M.; Elgaml, A.; Ahmed, T.A.E.; Hassan, H.; Mottawea, W. Metabolic Influences of Gut Microbiota Dysbiosis on Inflammatory Bowel Disease. Front. Physiol. 2021, 12, 715506. [Google Scholar] [CrossRef]
- Anand, K.J.S.; Hickey, P.R. Pain and Its Effects in the Human Neonate and Fetus. New Engl. J. Med. 1987, 317, 1321–1329. [Google Scholar] [CrossRef]
- Slater, R.; Cantarella, A.; Gallella, S.; Worley, A.; Boyd, S.; Meek, J.; Fitzgerald, M. Cortical pain responses in human infants. J. Neurosci. 2006, 26, 3662–3666. [Google Scholar] [CrossRef]
- Dührsen, L.; Simons, S.H.; Dzietko, M.; Genz, K.; Bendix, I.; Boos, V.; Sifringer, M.; Tibboel, D.; Felderhoff-Mueser, U. Effects of repetitive exposure to pain and morphine treatment on the neonatal rat brain. Neonatology 2012, 103, 35–43. [Google Scholar] [CrossRef]
- Cruz-Aguliar, R.M.; Wantia, N.; Clavel, T.; Vehreschild, M.J.; Buch, T.; Bajbouj, M.; Haller, D.; Busch, D.; Schmid, R.M.; Stein-Thoeringer, C.K. An Open-Labeled Study on Fecal Microbiota Transfer in Irritable Bowel Syndrome Patients Reveals Improvement in Abdominal Pain Associated with the Relative Abundance of Akkermansia Muciniphila. Digestion 2019, 100, 127–138. [Google Scholar] [CrossRef]
- Horvath, A.; Dziechciarz, P.; Szajewska, H. Meta-analysis: Lactobacillus rhamnosus GG for abdominal pain-related functional gastrointestinal disorders in childhood. Aliment. Pharmacol. Ther. 2011, 33, 1302–1310. [Google Scholar] [CrossRef]
- Saulnier, D.M.; Riehle, K.; Mistretta, T.A.; Diaz, M.A.; Mandal, D.; Raza, S.; Weidler, E.M.; Qin, X.; Coarfa, C.; Milosavljevic, A.; et al. Gastrointestinal microbiome signatures of pediatric patients with irritable bowel syndrome. Gastroenterology 2011, 141, 1782–1791. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoine, D.; Singh, P.K.; Tao, J.; Roy, S. Neonatal Morphine Results in Long-Lasting Alterations to the Gut Microbiome in Adolescence and Adulthood in a Murine Model. Pharmaceutics 2022, 14, 1879. https://doi.org/10.3390/pharmaceutics14091879
Antoine D, Singh PK, Tao J, Roy S. Neonatal Morphine Results in Long-Lasting Alterations to the Gut Microbiome in Adolescence and Adulthood in a Murine Model. Pharmaceutics. 2022; 14(9):1879. https://doi.org/10.3390/pharmaceutics14091879
Chicago/Turabian StyleAntoine, Danielle, Praveen Kumar Singh, Junyi Tao, and Sabita Roy. 2022. "Neonatal Morphine Results in Long-Lasting Alterations to the Gut Microbiome in Adolescence and Adulthood in a Murine Model" Pharmaceutics 14, no. 9: 1879. https://doi.org/10.3390/pharmaceutics14091879
APA StyleAntoine, D., Singh, P. K., Tao, J., & Roy, S. (2022). Neonatal Morphine Results in Long-Lasting Alterations to the Gut Microbiome in Adolescence and Adulthood in a Murine Model. Pharmaceutics, 14(9), 1879. https://doi.org/10.3390/pharmaceutics14091879