


Antimicrobial Peptides and Biomarkers Induced by Ultraviolet Irradiation Have the Potential to Reduce Endodontic Inflammation and Facilitate Tissue Healing


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset of UV-Induced Molecules
2.2. Subset of Endodontic Tissue Molecules
2.3. Analysis
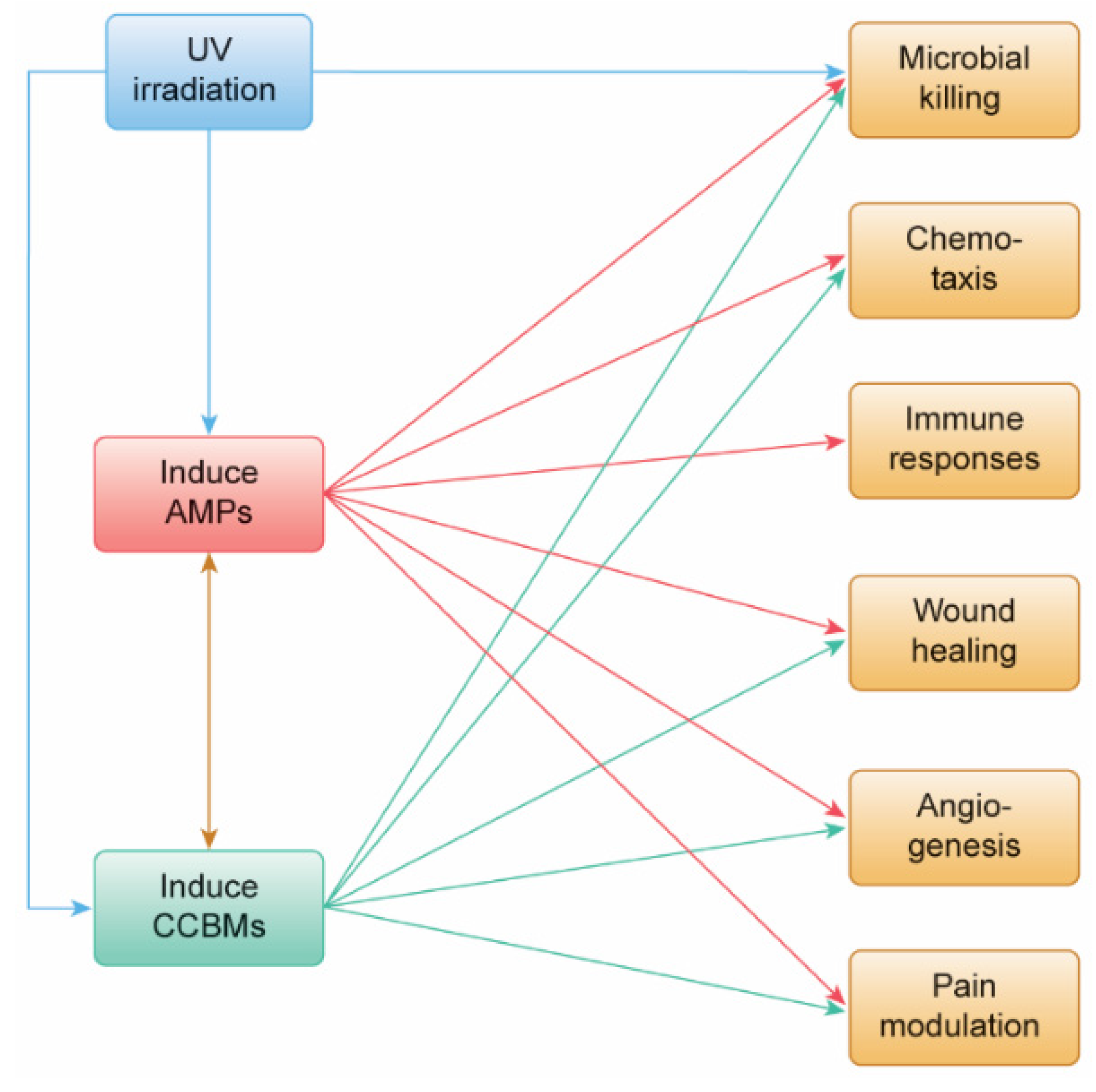
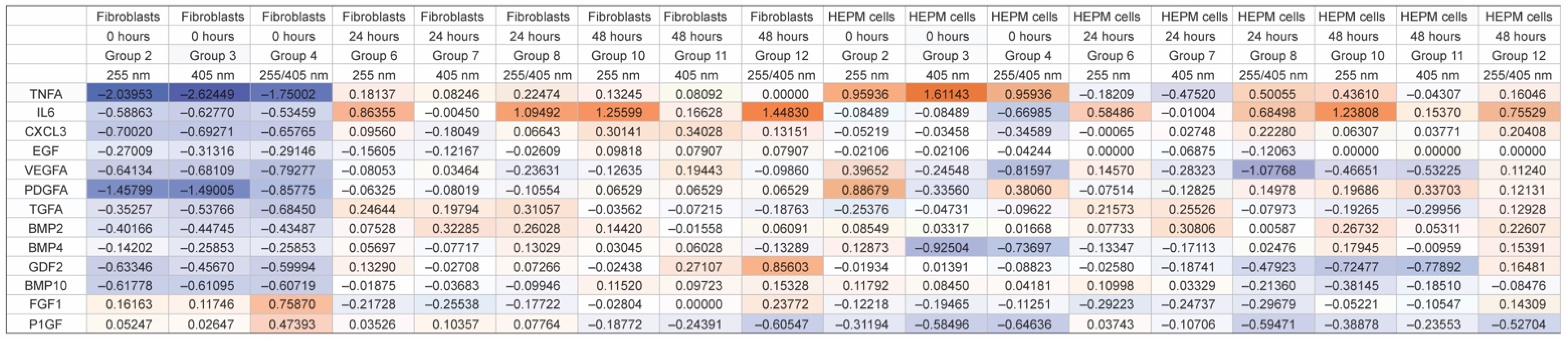
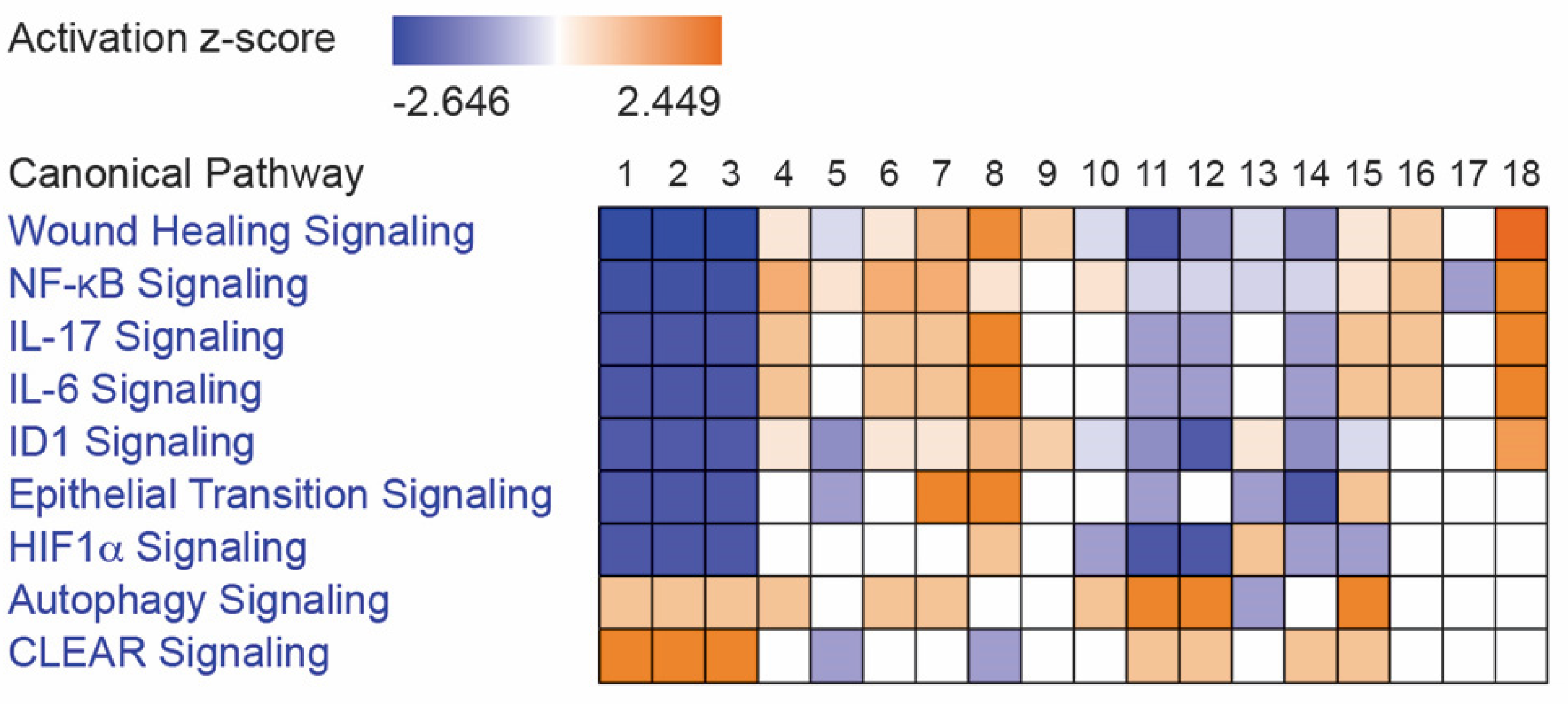
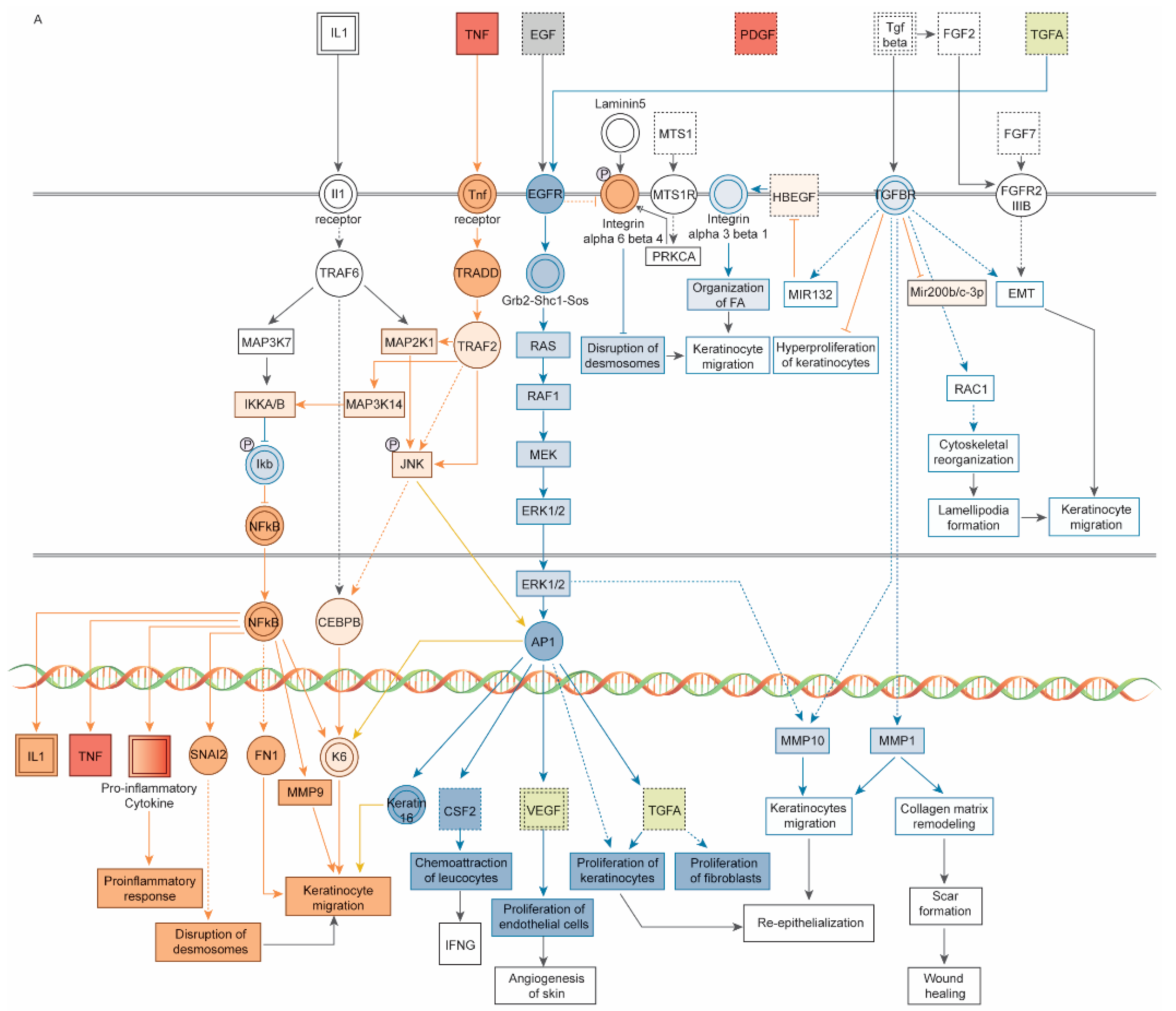
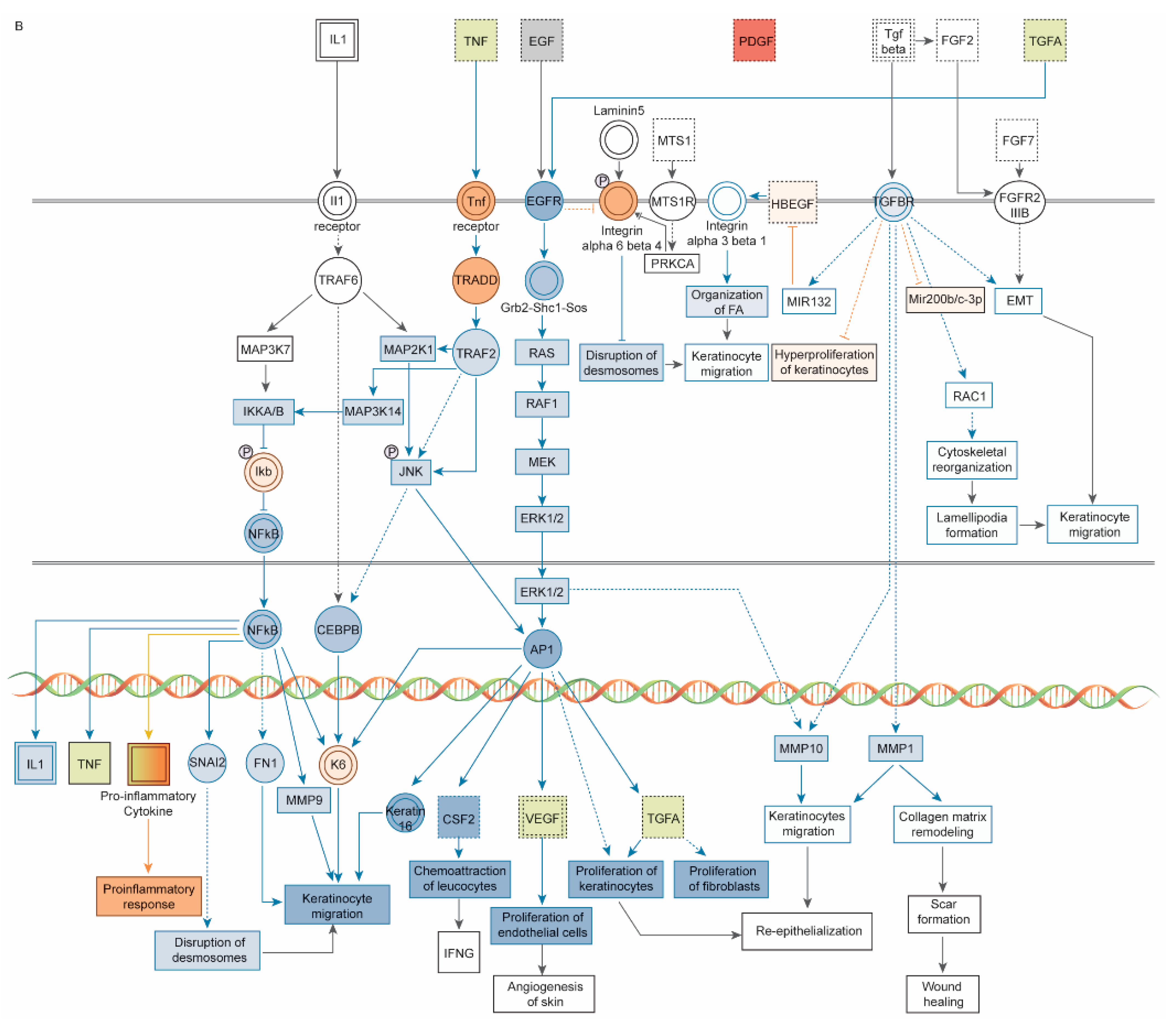
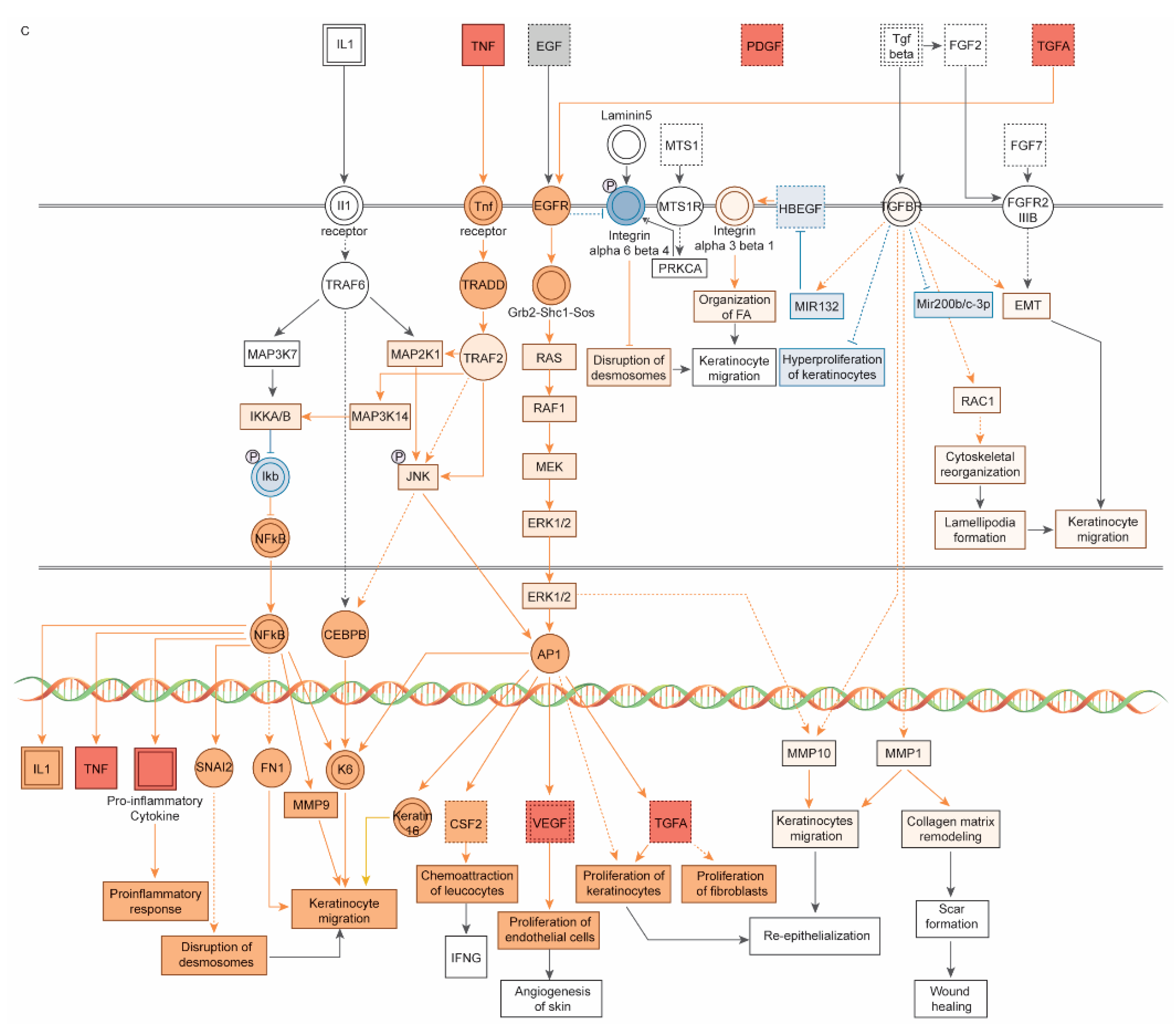
3. Results
3.1. Dataset of 32 UV-Induced Molecules
3.2. Subset of 13 Endodontic Tissue Molecules
3.3. Modulating Endodontic Pain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fouad, A.F. Microbiological Aspects of Traumatic Injuries. J. Endod. 2019, 45, S39–S48. [Google Scholar] [CrossRef] [PubMed]
- Delikan, E.; Caliskan, S.; Cankilic, M.Y.; Aksu, S.; Kesim, B.; Ulger, S.T. Microbiota of endodontically infected primary and permanent teeth. Pediatr. Dent. 2021, 43, 102–110. [Google Scholar] [PubMed]
- Wong, J.; Manoil, D.; Näsman, P.; Belibasakis, G.N.; Neelakantan, P. Microbiological aspects of root canal infections and disinfection strategies: An update review on the current knowledge and challenges. Front. Oral Health 2021, 2, 672887. [Google Scholar] [CrossRef]
- Siqueira, J.F., Jr.; Rocas, I.N. Present status and future directions: Microbiology of endodontic infections. Int. Endod. J. 2022, 55, 512–530. [Google Scholar] [CrossRef] [PubMed]
- Metzger, Z.; Better, H.; Abramovitz, I. Immediate root canal disinfection with ultraviolet light: An ex vivo feasibility study. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2007, 104, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Morio, K.A.; Thayer, E.L.; Bates, A.M.; Brogden, K.A. 255-nm light-emitting diode kills Enterococcus faecalis and induces the production of cellular biomarkers in human embryonic palatal mesenchyme cells and gingival fibroblasts. J. Endod. 2019, 45, 774–783.e6. [Google Scholar] [CrossRef]
- Metzger, Z.; Dotan, M.; Better, H.; Abramovitz, I. Sensitivity of oral bacteria to 254 nm ultraviolet light. Int. Endod. J. 2007, 40, 120–127. [Google Scholar] [CrossRef]
- Morio, K.A.; Sternowski, R.H.; Brogden, K.A. Dataset of endodontic microorganisms killed at 265 nm wavelength by an ultraviolet C light emitting diode in root canals of extracted, instrumented teeth. Data Brief 2022, 40, 107750. [Google Scholar] [CrossRef]
- Ou, Y.; Petersen, P.M. Application of ultraviolet light sources for in vivo disinfection. Jpn. J. Appl. Phys. 2021, 60, 100501. [Google Scholar] [CrossRef]
- Vieyra-Garcia, P.A.; Wolf, P. A deep dive into UV-based phototherapy: Mechanisms of action and emerging molecular targets in inflammation and cancer. Pharmacol. Ther. 2021, 222, 107784. [Google Scholar] [CrossRef]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Morykwas, M.J.; Mark, M.W. Effects of ultraviolet light on fibroblast fibronectin production and lattice contraction. Wounds 1998, 10, 111–117. [Google Scholar]
- Dennler, S.; Itoh, S.; Vivien, D.; Dijke, P.; Huet, S.; Gauthier, J.M. Direct binding of Smad3 and Smad4 to critical TGF beta-inducible elements in the promoter of human plasminogen activator inhibitor-type 1 gene. EMBO J. 1998, 17, 3091–3100. [Google Scholar] [CrossRef] [PubMed]
- Hagai, T.; Chen, X.; Miragaia, R.J.; Rostom, R.; Gomes, T.; Kunowska, N.; Henriksson, J.; Park, J.E.; Proserpio, V.; Donati, G.; et al. Gene expression variability across cells and species shapes innate immunity. Nature 2018, 563, 197–202. [Google Scholar] [CrossRef]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A.; Bates, A.M.; Fischer, C.L. Antimicrobial peptides in host defense: Functions beyond antimicrobial activity. In Antimicrobial Peptides—Role in Human Health and Disease; Harder, J., Schroeder, J.M., Kaufmann, S.H., Mercer, A.A., Weber, B., Eds.; Birkhauser Advances in Infectious Diseases; Springer International Publishing: Cham, Switzerland, 2016; pp. 129–146. [Google Scholar] [CrossRef]
- Drayton, M.; Deisinger, J.P.; Ludwig, K.C.; Raheem, N.; Muller, A.; Schneider, T.; Straus, S.K. Host defense peptides: Dual antimicrobial and immunomodulatory action. Int. J. Mol. Sci. 2021, 22, 11172. [Google Scholar] [CrossRef]
- Takahashi, M.; Umehara, Y.; Yue, H.; Trujillo-Paez, J.V.; Peng, G.; Nguyen, H.L.T.; Ikutama, R.; Okumura, K.; Ogawa, H.; Ikeda, S.; et al. The antimicrobial peptide human beta-defensin-3 accelerates wound healing by promoting angiogenesis, cell migration, and proliferation through the FGFR/JAK2/STAT3 signaling pathway. Front. Immunol. 2021, 12, 712781. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Human antimicrobial peptides and proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef]
- Mallbris, L.; Edstrom, D.W.; Sundblad, L.; Granath, F.; Stahle, M. UVB upregulates the antimicrobial protein hCAP18 mRNA in human skin. J. Investig. Dermatol. 2005, 125, 1072–1074. [Google Scholar] [CrossRef]
- Glaser, R.; Navid, F.; Schuller, W.; Jantschitsch, C.; Harder, J.; Schroder, J.M.; Schwarz, A.; Schwarz, T. UV-B radiation induces the expression of antimicrobial peptides in human keratinocytes in vitro and in vivo. J. Allergy Clin. Immunol. 2009, 123, 1117–1123. [Google Scholar] [CrossRef]
- Kreuter, A.; Hyun, J.; Skrygan, M.; Sommer, A.; Bastian, A.; Altmeyer, P.; Gambichler, T. Ultraviolet A1-induced downregulation of human beta-defensins and interleukin-6 and interleukin-8 correlates with clinical improvement in localized scleroderma. Br. J. Dermatol. 2006, 155, 600–607. [Google Scholar] [CrossRef]
- Gambichler, T.; Skrygan, M.; Tomi, N.S.; Altmeyer, P.; Kreuter, A. Changes of antimicrobial peptide mRNA expression in atopic eczema following phototherapy. Br. J. Dermatol. 2006, 155, 1275–1278. [Google Scholar] [CrossRef]
- Park, K.C.; Jung, H.C.; Hwang, J.H.; Youn, S.W.; Ahn, J.S.; Park, S.B.; Kim, K.H.; Chung, J.H.; Youn, J.I. GM-CSF production by epithelial cell line: Upregulation by ultraviolet A. Photodermatol. Photoimmunol. Photomed. 1997, 13, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Yada, Y.; Kimura, M.; Morisaki, N. Granulocyte/macrophage colony-stimulating factor is an intrinsic keratinocyte-derived growth factor for human melanocytes in UVA-induced melanosis. Biochem. J. 1996, 313, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Blaudschun, R.; Sunderkotter, C.; Brenneisen, P.; Hinrichs, R.; Peters, T.; Schneider, L.; Razi-Wolf, Z.; Hunzelmann, N.; Scharffetter-Kochanek, K. Vascular endothelial growth factor causally contributes to the angiogenic response upon ultraviolet B irradiation in vivo. Br. J. Dermatol. 2002, 146, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Oxholm, A.; Oxholm, P.; Staberg, B.; Bendtzen, K. Immunohistological detection of interleukin I-like molecules and tumour necrosis factor in human epidermis before and after UVB-irradiation in vivo. Br. J. Dermatol. 1988, 118, 369–376. [Google Scholar] [CrossRef]
- Crispin, M.K.; Fuentes-Duculan, J.; Gulati, N.; Johnson-Huang, L.M.; Lentini, T.; Sullivan-Whalen, M.; Gilleaudeau, P.; Cueto, I.; Suárez-Fariñas, M.; Lowes, M.A.; et al. Gene profiling of narrowband UVB-induced skin injury defines cellular and molecular innate immune responses. J. Investig. Dermatol. 2013, 133, 692–701. [Google Scholar] [CrossRef]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schroder, J.M.; Wang, J.M.; Howard, O.M.; et al. B-defensins: Linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial peptides human beta-defensins stimulate epidermal keratinocyte migration, proliferation and production of proinflammatory cytokines and chemokines. J. Investig. Dermatol. 2007, 127, 594–604. [Google Scholar] [CrossRef]
- Ramos, R.; Silva, J.P.; Rodrigues, A.C.; Costa, R.; Guardao, L.; Schmitt, F.; Soares, R.; Vilanova, M.; Domingues, L.; Gama, M. Wound healing activity of the human antimicrobial peptide LL37. Peptides 2011, 32, 1469–1476. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Biragyn, A.; Kwak, L.W.; Oppenheim, J.J. Mammalian defensins in immunity: More than just microbicidal. Trends Immunol. 2002, 23, 291–296. [Google Scholar] [CrossRef]
- Chaly, Y.V.; Paleolog, E.M.; Kolesnikova, T.S.; Tikhonov, I.I.; Petratchenko, E.V.; Voitenok, N.N. Neutrophil alpha-defensin human neutrophil peptide modulates cytokine production in human monocytes and adhesion molecule expression in endothelial cells. Eur. Cytokine Netw. 2000, 11, 257–266. [Google Scholar] [PubMed]
- Barabas, N.; Rohrl, J.; Holler, E.; Hehlgans, T. Beta-defensins activate macrophages and synergize in pro-inflammatory cytokine expression induced by TLR ligands. Immunobiology 2013, 218, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Van Wetering, S.; MannesseLazeroms, S.P.G.; Dijkman, J.H.; Hiemstra, P.S. Effect of neutrophil serine proteinases and defensins on lung epithelial cells: Modulation of cytotoxicity and IL-8 production. J. Leukoc. Biol. 1997, 62, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.; Funderburg, N.; Weinberg, A.; Sieg, S. Human beta defensin-3 induces chemokines from monocytes and macrophages: Diminished activity in cells from HIV-infected persons. Immunology 2013, 140, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.G.; Davidson, D.J.; Gold, M.R.; Bowdish, D.; Hancock, R.E. The human antimicrobial peptide LL-37 is a multifunctional modulator of innate immune responses. J. Immunol. 2002, 169, 3883–3891. [Google Scholar] [CrossRef]
- Warnke, P.H.; Voss, E.; Russo, P.A.; Stephens, S.; Kleine, M.; Terheyden, H.; Liu, Q. Antimicrobial peptide coating of dental implants: Biocompatibility assessment of recombinant human beta defensin-2 for human cells. Int. J. Oral Maxillofac. Implant. 2013, 28, 982–988. [Google Scholar] [CrossRef]
- Heilborn, J.D.; Nilsson, M.F.; Kratz, G.; Weber, G.; Sorensen, O.; Borregaard, N.; Stahle-Backdahl, M. The cathelicidin anti-microbial peptide LL-37 is involved in re-epithelialization of human skin wounds and is lacking in chronic ulcer epithelium. J. Investig. Dermatol. 2003, 120, 379–389. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Gallo, R.L. Antimicrobial peptides: Old molecules with new ideas. J. Investig. Dermatol. 2012, 132, 887–895. [Google Scholar] [CrossRef]
- Schwarz, T.; Beissert, S. Milestones in photoimmunology. J. Investig. Dermatol. 2013, 133, E7–E10. [Google Scholar] [CrossRef] [Green Version]
- Matos, T.R.; Sheth, V. The symbiosis of phototherapy and photoimmunology. Clin. Dermatol. 2016, 34, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Bruhs, A.; Schwarz, T. Ultraviolet radiation-induced immunosuppression: Induction of regulatory T cells. Methods Mol. Biol. 2017, 1559, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Hequet, O.; Nosbaum, A.; Guironnet-Paquet, A.; Blasco, E.; Nicolas-Virelizier, E.; Griffith, T.S.; Rigal, D.; Cognasse, F.; Nicolas, J.F.; Vocanson, M. CD8(+) T cells mediate ultraviolet A-induced immunomodulation in a model of extracorporeal photochemotherapy. Eur. J. Immunol. 2020, 50, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, T. Regulatory T cells induced by ultraviolet radiation. Int. Arch. Allergy Immunol. 2005, 137, 187–193. [Google Scholar] [CrossRef]
- Bernard, J.J.; Gallo, R.L.; Krutmann, J. Photoimmunology: How ultraviolet radiation affects the immune system. Nat. Rev. Immunol. 2019, 19, 688–701. [Google Scholar] [CrossRef]
- Patil, C.S.; Kirkwood, K.L. p38 MAPK signaling in oral-related diseases. J. Dent. Res. 2007, 86, 812–825. [Google Scholar] [CrossRef]
- Ji, R.R.; Gereau, R.W.; Malcangio, M.; Strichartz, G.R. MAP kinase and pain. Brain Res. Rev. 2009, 60, 135–148. [Google Scholar] [CrossRef]
- Muthusamy, V.; Piva, T.J. A comparative study of UV-induced cell signalling pathways in human keratinocyte-derived cell lines. Arch. Dermatol. Res. 2013, 305, 817–833. [Google Scholar] [CrossRef]
- Ma, W.; Quirion, R. The ERK/MAPK pathway, as a target for the treatment of neuropathic pain. Expert Opin. Ther. Targets 2005, 9, 699–713. [Google Scholar] [CrossRef]
- Lin, X.; Wang, M.; Zhang, J.; Xu, R. p38 MAPK: A potential target of chronic pain. Curr. Med. Chem. 2014, 21, 4405–4418. [Google Scholar] [CrossRef]
- Bode, A.M.; Dong, Z. Mitogen-activated protein kinase activation in UV-induced signal transduction. Sci. STKE 2003, 2003, 167. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.; Blattner, C.; Knebel, A.; Iordanov, M.; Herrlich, P.; Rahmsdorf, H.J. UV-induced signal transduction. J. Photochem. Photobiol. B. 1997, 37, 1–17. [Google Scholar] [CrossRef]
- Adachi, M.; Gazel, A.; Pintucci, G.; Shuck, A.; Shifteh, S.; Ginsburg, D.; Rao, L.S.; Kaneko, T.; Freedberg, I.M.; Tamaki, K.; et al. Specificity in stress response: Epidermal keratinocytes exhibit specialized UV-responsive signal transduction pathways. DNA Cell Biol. 2003, 22, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.J.; Ahn, S.W.; Hong, C.K.; Ro, B.I. Expressions of beta-defensins in human keratinocyte cell lines. J. Dermatol. Sci. 2001, 27, 183–191. [Google Scholar] [CrossRef]
- Suo, W.; Guo, H.; Wang, X.; Wang, D. Effect of ultraviolet C light on the expression of basic fibroblast growth factor in rat wounds. Chinese J. Phys. Med. Rehabil. 2003, 25, 651–654. [Google Scholar]
- Suo, W.; Wang, X.; Wang, D. Effect of ultraviolet C irradiation on expression of transforming growth factor-β in wound. Chin. J. Rehabil. Theory Pract. 2002, 8, 5–7. [Google Scholar]
- Gallo, R.L.; Staszewski, R.; Sauder, D.N.; Knisely, T.L.; Granstein, R.D. Regulation of GM-CSF and IL-3 production from the murine keratinocyte cell line PAM 212 following exposure to ultraviolet radiation. J. Investig. Dermatol. 1991, 97, 203–209. [Google Scholar] [CrossRef]
- Kirnbauer, R.; Koch, A.; Kurtmann, J.; Schwarz, T.; Urbanski, A.; Luger, T.A. Different effects of UVA and UVB irradiation on epidermal cell-IL6 expression and release. In Proceedings of the ESDR-JSID-SID Tricontinental Meeting, Washington, DC, USA, 26 April 1989; pp. 393–548. [Google Scholar]
- Gallo, R.L.; Brownstein, E.; Granstein, R.D. Secretion of interleukin 3 activity from a transformed murine keratinocyte line after exposure to ultraviolet radiation: Role of membrane signal transduction mechanisms. In Proceedings of the ESDR-JSID-SID Tricontinental Meeting, Washington, DC, USA, 26 April 1989; pp. 393–548. [Google Scholar]
- Clingen, P.H.; Berneburg, M.; Petit-Frere, C.; Woollons, A.; Lowe, J.E.; Arlett, C.F.; Green, M.H. Contrasting effects of an ultraviolet B and an ultraviolet A tanning lamp on interleukin-6, tumour necrosis factor-alpha and intercellular adhesion molecule-1 expression. Br. J. Dermatol. 2001, 145, 54–62. [Google Scholar] [CrossRef]
- Chung, K.Y.; Chang, N.S.; Park, Y.K.; Lee, K.H. Effect of ultraviolet light on the expression of adhesion molecules and T lymphocyte adhesion to human dermal microvascular endothelial cells. Yonsei Med. J. 2002, 43, 165–174. [Google Scholar] [CrossRef]
- Ansel, J.C.; Luger, T.A.; Green, I. The effect of in vitro and in vivo UV irradiation on the production of ETAF activity by human and murine keratinocytes. J. Investig. Dermatol. 1983, 81, 519–523. [Google Scholar] [CrossRef]
- Kupper, T.S.; Chua, A.O.; Flood, P.; McGuire, J.; Gubler, U. Interleukin 1 gene expression in cultured human keratinocytes is augmented by ultraviolet irradiation. J. Clin. Investig. 1987, 80, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, T.; Luger, T.A. New trends in photobiology: Effect of UV irradiation on epidermal cell cytokine production. J. Photochem. Photobiol. B: Biol. 1989, 4, 1–13. [Google Scholar] [CrossRef]
- Rhodes, L.E.; Joyce, M.; West, D.C.; Strickland, I.; Friedmann, P.S. Comparison of changes in endothelial adhesion molecule expression following UVB irradiation of skin and a human dermal microvascular cell line (HMEC-1). Photodermatol. Photoimmunol. Photomed. 1996, 12, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Kennedy-Crispin, M.; Billick, E.; Mitsui, H.; Gulati, N.; Fujita, H.; Gilleaudeau, P.; Sullivan-Whalen, M.; Johnson-Huang, L.M.; Suarez-Farinas, M.; Krueger, J.G. Human keratinocytes’ response to injury upregulates CCL20 and other genes linking innate and adaptive immunity. J. Investig. Dermatol. 2012, 132, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Kirnbauer, R.; Kock, A.; Neuner, P.; Forster, E.; Krutmann, J.; Urbanski, A.; Schauer, E.; Ansel, J.C.; Schwarz, T.; Luger, T.A. Regulation of epidermal cell interleukin-6 production by UV light and corticosteroids. J. Investig. Dermatol. 1991, 96, 484–489. [Google Scholar] [CrossRef]
- Yarosh, D.; Both, D.; Kibitel, J.; Anderson, C.; Elmets, C.; Brash, D.; Brown, D. Regulation of TNFalpha production and release in human and mouse keratinocytes and mouse skin after UV-B irradiation. Photodermatol. Photoimmunol. Photomed. 2000, 16, 263–270. [Google Scholar] [CrossRef]
- Gambichler, T.; Tomi, N.S.; Skrygan, M.; Altmeyer, P.; Kreuter, A. Alterations of TGF-beta/Smad mRNA expression in atopic dermatitis following narrow-band ultraviolet B phototherapy: Results of a pilot study. J. Dermatol. Sci. 2006, 44, 56–58. [Google Scholar] [CrossRef]
- James, L.C.; Moore, A.M.; Wheeler, L.A.; Murphy, G.M.; Dowd, P.M.; Greaves, M.W. Transforming growth factor alpha: In vivo release by normal human skin following UV irradiation and abrasion. Skin Pharmacol. 1991, 4, 61–64. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Entrez Gene Name | Entrez Gene ID (Human) | Location | Function Type |
|---|---|---|---|---|
| BMP10 | Bone morphogenetic protein 10 | 27302 | Extracellular space | Growth factor |
| CAMP | Cathelicidin antimicrobial peptide | 820 | Cytoplasm | Other |
| CCL2 | C-C motif chemokine ligand 2 | 6347 | Extracellular space | Cytokine |
| CCL20 | C-C motif chemokine ligand 20 | 6364 | Extracellular space | Cytokine |
| CSF2 | Colony stimulating factor 2 | 1437 | Extracellular space | Cytokine |
| CXCL1 | C-X-C motif chemokine ligand 1 | 2919 | Extracellular space | Cytokine |
| CXCL2 | C-X-C motif chemokine ligand 2 | 2920 | Extracellular space | Cytokine |
| CXCL3 | C-X-C motif chemokine ligand 3 | 2921 | Extracellular space | Cytokine |
| CXCL8 | C-X-C motif chemokine ligand 8 | 3576 | Extracellular space | Cytokine |
| DEFB1 | Defensin beta 1 | 1672 | Extracellular space | Other |
| DEFB103B | Defensin beta 103B | 55894 | Extracellular space | Other |
| DEFB4A | Defensin beta 4A | 1673 | Extracellular space | Other |
| FGF1 | Fibroblast growth factor 1 | 2246 | Extracellular space | Growth factor |
| FGF2 | Fibroblast growth factor 2 | 2247 | Extracellular space | Growth factor |
| FN1 | Fibronectin 1 | 2335 | Extracellular space | Enzyme |
| ICAM1 | Intercellular adhesion molecule 1 | 3383 | Plasma membrane | Transmembrane receptor |
| IL6 | Interleukin 6 | 3569 | Extracellular space | Cytokine |
| IL10 | Interleukin 10 | 3586 | Extracellular space | Cytokine |
| PI3 | Peptidase inhibitor 3 | 5266 | Extracellular space | Other |
| PIGF | Phosphatidylinositol glycan anchor biosynthesis class F | 5281 | Cytoplasm | Enzyme |
| RNASE7 | Ribonuclease A family member 7 | 84659 | Extracellular space | Enzyme |
| S100A7 | S100 calcium binding protein A7 | 6278 | Cytoplasm | Other |
| S100A8 | S100 calcium binding protein A8 | 6279 | Cytoplasm | Other |
| S100A9 | S100 calcium binding protein A9 | 6280 | Cytoplasm | Other |
| S100A12 | S100 calcium binding protein A12 | 6283 | Cytoplasm | Other |
| SELE | Selectin E | 6401 | Plasma membrane | Transmembrane receptor |
| SMAD3 | SMAD family member 3 | 4088 | Nucleus | Transcription regulator |
| SMAD4 | SMAD family member 4 | 4089 | Nucleus | Transcription regulator |
| TGFA | Transforming growth factor alpha | 7039 | Extracellular space | Growth factor |
| TGFB1 | Transforming growth factor beta 1 | 7040 | Extracellular space | Growth factor |
| TNF | Tumor necrosis factor | 7124 | Extracellular space | Cytokine |
| VEGFA | Vascular endothelial growth factor A | 7422 | Extracellular space | Growth factor |
| IPA Function | p-Value | No. | Identification of Molecules |
|---|---|---|---|
| Canonical Pathway Annotations | |||
| Cellular Stress and Injury | |||
| Wound healing signaling pathway | 3.22 × 10−11 | 9 | CSF2, CXCL8, FGF2, FN1, IL6, TGFA, TGFB1, TNF, VEGFA |
| Cytokine Signaling | |||
| IL17 signaling | 8.43 × 10−20 | 13 | CCL2, CCL20, CSF2, CXCL1, CXCL3, CXCL8, DEFB1, DEFB103A/DEFB103B, DEFB4A/DEFB4B, IL6, TGFB1, TNF, VEGFA |
| IL6 signaling | 2.53 × 10−5 | 4 | CXCL8, IL6, TNF, VEGFA |
| IL10 signaling | 1.23 × 10−4 | 3 | IL10, IL6, TNF |
| IL8 signaling | 1.73 × 10−4 | 4 | CXCL1, CXCL8, ICAM1, VEGFA |
| Cellular Immune Response | |||
| Role of cytokines in mediating communication between immune cells | 8.74 × 10−11 | 6 | CSF2, CXCL8, IL10, IL6, TGFB1, TNF |
| Th1 and Th2 activation pathway | 8.01 × 10−5 | 4 | ICAM1, IL10, IL6, TGFB1 |
| Th1 pathway | 5.81 × 10−4 | 3 | ICAM1, IL10, IL6 |
| Th2 pathway | 8.13 × 10−4 | 3 | ICAM1, IL10, TGFB1 |
| Communication between innate and adaptive immune cells | 7.44 × 10−3 | 5 | CSF2, CXCL8, IL10, IL6, TNF |
| Diseases or Functions Annotations | |||
| Antimicrobial Response, Inflammatory Response | |||
| Antibacterial response | 1.66 × 10−22 | 13 | CAMP, CCL20, DEFB1, DEFB103A/DEFB103B, DEFB4A/DEFB4B, IL10, IL6, RNASE7, S100A12, S100A7, S100A8, S100A9, TNF |
| Antimicrobial Response, Inflammatory Response | |||
| Chemoattraction | 5.09 × 10−23 | 12 | CAMP, CCL2, CCL20, CSF2, CXCL1, CXCL3, CXCL8, DEFB4A/DEFB4B, FN1, TGFB1, TNF, VEGFA |
| Cellular Movement, Hematological System Development and Function, Immune Cell Trafficking, Inflammatory Response | |||
| Chemotaxis | 4.28 × 10−37 | 27 | CAMP, CCL2, CCL20, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, DEFB1, DEFB103A/DEFB103B, DEFB4A/DEFB4B, FGF2, FN1, ICAM1, IL10, IL6, S100A12, S100A7, S100A8, S100A9, SELE, SMAD3, SMAD4, TGFA, TGFB1, TNF, VEGFA |
| Chemotaxis of leukocytes | 1.43 × 10−35 | 24 | CAMP, CCL2, CCL20, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, DEFB1, DEFB103A/DEFB103B, DEFB4A/DEFB4B, FN1, ICAM1, IL10, IL6, S100A12, S100A7, S100A8, S100A9, SELE, SMAD3, TGFB1, TNF, VEGFA |
| Inflammatory Response | |||
| Inflammatory response | 9.34 × 10−29 | 26 | CAMP, CCL2, CCL20, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, DEFB1, DEFB103A/DEFB103B, DEFB4A/DEFB4B, FGF1, FGF2, FN1, ICAM1, IL10, IL6, S100A12, S100A7, S100A8, S100A9, SELE, SMAD3, TGFB1, TNF, VEGFA |
| Proinflammatory response | 4.92 × 10−15 | 7 | CCL2, CXCL3, CXCL8, IL10, IL6, TNF, VEGFA |
| Innate immune response | 5.94 × 10−14 | 10 | CAMP, CXCL1, CXCL8, FN1, IL10, IL6, RNASE7, S100A12, SMAD3, TNF |
| Tissue Development | |||
| Healing of wound | 1.53 × 10−20 | 13 | CSF2, FGF1, FGF2, FN1, ICAM1, IL10, IL6, SMAD3, SMAD4, TGFA, TGFB1, TNF, VEGFA |
| Cell-To-Cell Signaling and Interaction, Cellular Movement, Hematological System Development and Function, Immune Cell Trafficking, Inflammatory Response | |||
| Cell movement of monocytes | 1.37 × 10−31 | 19 | CAMP, CCL2, CCL20, CSF2, CXCL3, CXCL8, DEFB1, DEFB103A/DEFB103B, FN1, ICAM1, IL10, IL6, S100A12, S100A7, SELE, SMAD3, TGFB1, TNF, VEGFA |
| Cell movement of neutrophils | 2.01 × 10−28 | 21 | CAMP, CCL2, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, DEFB1, DEFB103A/DEFB103B, DEFB4A/DEFB4B, FN1, ICAM1, IL10, IL6, S100A12, S100A8, S100A9, SELE, SMAD3, TGFB1, TNF |
| Cell-To-Cell Signaling and Interaction, Cellular Movement, Hematological System Development and Function, Immune Cell Trafficking, Inflammatory Response | |||
| Recruitment of cells | 7.85 × 10−29 | 21 | BMP10, CAMP, CCL2, CCL20, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, DEFB4A/DEFB4B, FGF2, FN1, ICAM1, IL10, IL6, S100A8, SELE, SMAD3, TGFB1, TNF, VEGFA |
| Recruitment of leukocytes | 1.05 × 10−25 | 19 | BMP10, CAMP, CCL2, CCL20, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, DEFB4A/DEFB4B, FN1, ICAM1, IL10, IL6, S100A8, SELE, SMAD3, TGFB1, TNF |
| Cell-To-Cell Signaling and Interaction, Inflammatory Response | |||
| Immune response of cells | 2.70 × 10−18 | 18 | CAMP, CCL2, CCL20, CSF2, CXCL1, CXCL3, CXCL8, FN1, ICAM1, IL10, IL6, S100A12, S100A8, S100A9, SMAD3, TGFB1, TNF, VEGFA |
| Immune response of myeloid cells | 2.85 × 10−17 | 13 | CAMP, CCL2, CSF2, CXCL1, CXCL3, CXCL8, FN1, ICAM1, IL10, IL6, S100A9, TGFB1, TNF |
| Cellular Growth and Proliferation | |||
| Angiogenesis | 1.74 × 10−23 | 24 | BMP10, CAMP, CCL2, CSF2, CXCL1, CXCL2, CXCL8, FGF1, FGF2, FN1, ICAM1, IL10, IL6, PIGF, S100A12, S100A8, S100A9, SELE, SMAD3, SMAD4, TGFA, TGFB1, TNF, VEGFA |
| Proliferation of vascular cells | 3.32 × 10−20 | 16 | CAMP, CCL2, CXCL1, CXCL8, FGF1, FGF2, FN1, IL10, IL6, S100A8, S100A9, SMAD3, SMAD4, TGFB1, TNF, VEGFA |
| Cellular Movement, Hematological System Development and Function, Immune Cell Trafficking | |||
| Cell survival | 1.73 × 10−17 | 24 | CAMP, CCL2, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, DEFB103A/DEFB103B, DEFB4A/DEFB4B, FGF1, FGF2, FN1, ICAM1, IL10, IL6, S100A8, S100A9, SELE, SMAD3, SMAD4, TGFA, TGFB1, TNF, VEGFA |
| Cell viability | 2.74 × 10−15 | 22 | CAMP, CCL2, CSF2, CXCL1, CXCL2, CXCL3, CXCL8, FGF1, FGF2, FN1, ICAM1, IL10, IL6, S100A8, S100A9, SELE, SMAD3, SMAD4, TGFA, TGFB1, TNF, VEGFA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morio, K.A.; Sternowski, R.H.; Zeng, E.; Brogden, K.A. Antimicrobial Peptides and Biomarkers Induced by Ultraviolet Irradiation Have the Potential to Reduce Endodontic Inflammation and Facilitate Tissue Healing. Pharmaceutics 2022, 14, 1979. https://doi.org/10.3390/pharmaceutics14091979
Morio KA, Sternowski RH, Zeng E, Brogden KA. Antimicrobial Peptides and Biomarkers Induced by Ultraviolet Irradiation Have the Potential to Reduce Endodontic Inflammation and Facilitate Tissue Healing. Pharmaceutics. 2022; 14(9):1979. https://doi.org/10.3390/pharmaceutics14091979
Chicago/Turabian StyleMorio, Kimberly A., Robert H. Sternowski, Erliang Zeng, and Kim A. Brogden. 2022. "Antimicrobial Peptides and Biomarkers Induced by Ultraviolet Irradiation Have the Potential to Reduce Endodontic Inflammation and Facilitate Tissue Healing" Pharmaceutics 14, no. 9: 1979. https://doi.org/10.3390/pharmaceutics14091979
APA StyleMorio, K. A., Sternowski, R. H., Zeng, E., & Brogden, K. A. (2022). Antimicrobial Peptides and Biomarkers Induced by Ultraviolet Irradiation Have the Potential to Reduce Endodontic Inflammation and Facilitate Tissue Healing. Pharmaceutics, 14(9), 1979. https://doi.org/10.3390/pharmaceutics14091979