

Current Advances of Plant-Based Vaccines for Neurodegenerative Diseases

 , ,
, ,  ,
,  and
and 
Abstract
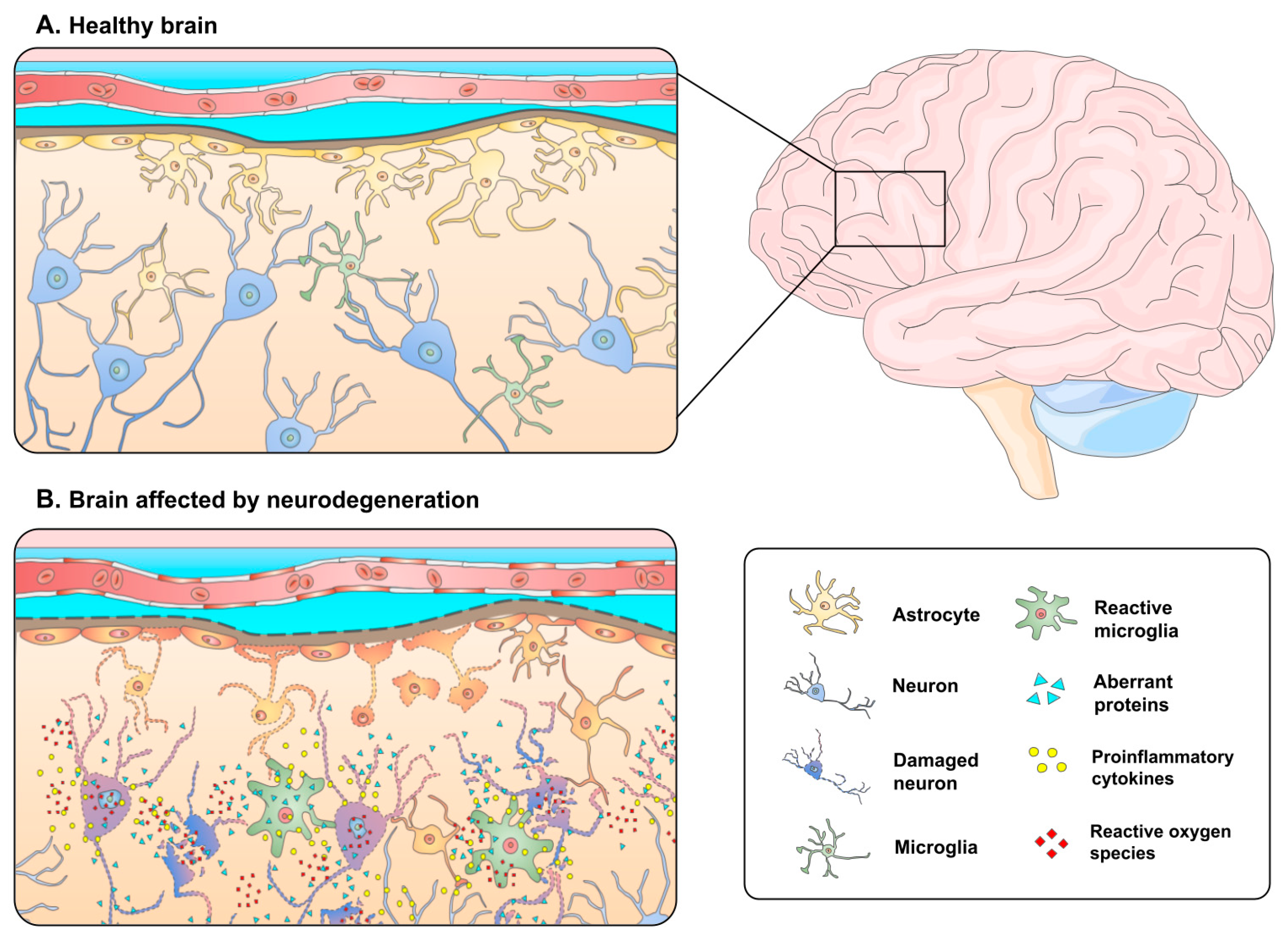
:1. Introduction
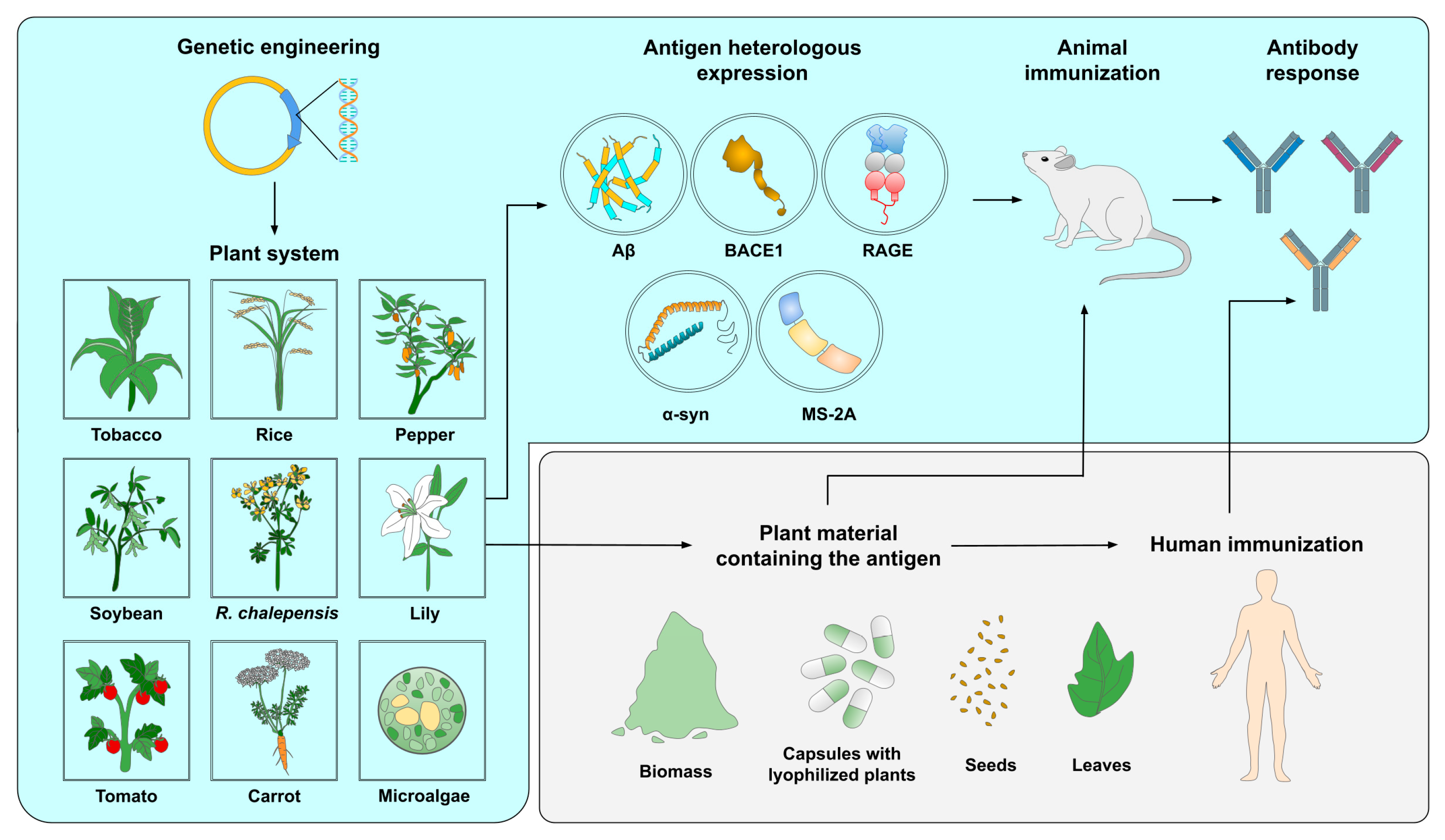
2. Overview of the Present Landscape of Plant-Based Vaccines
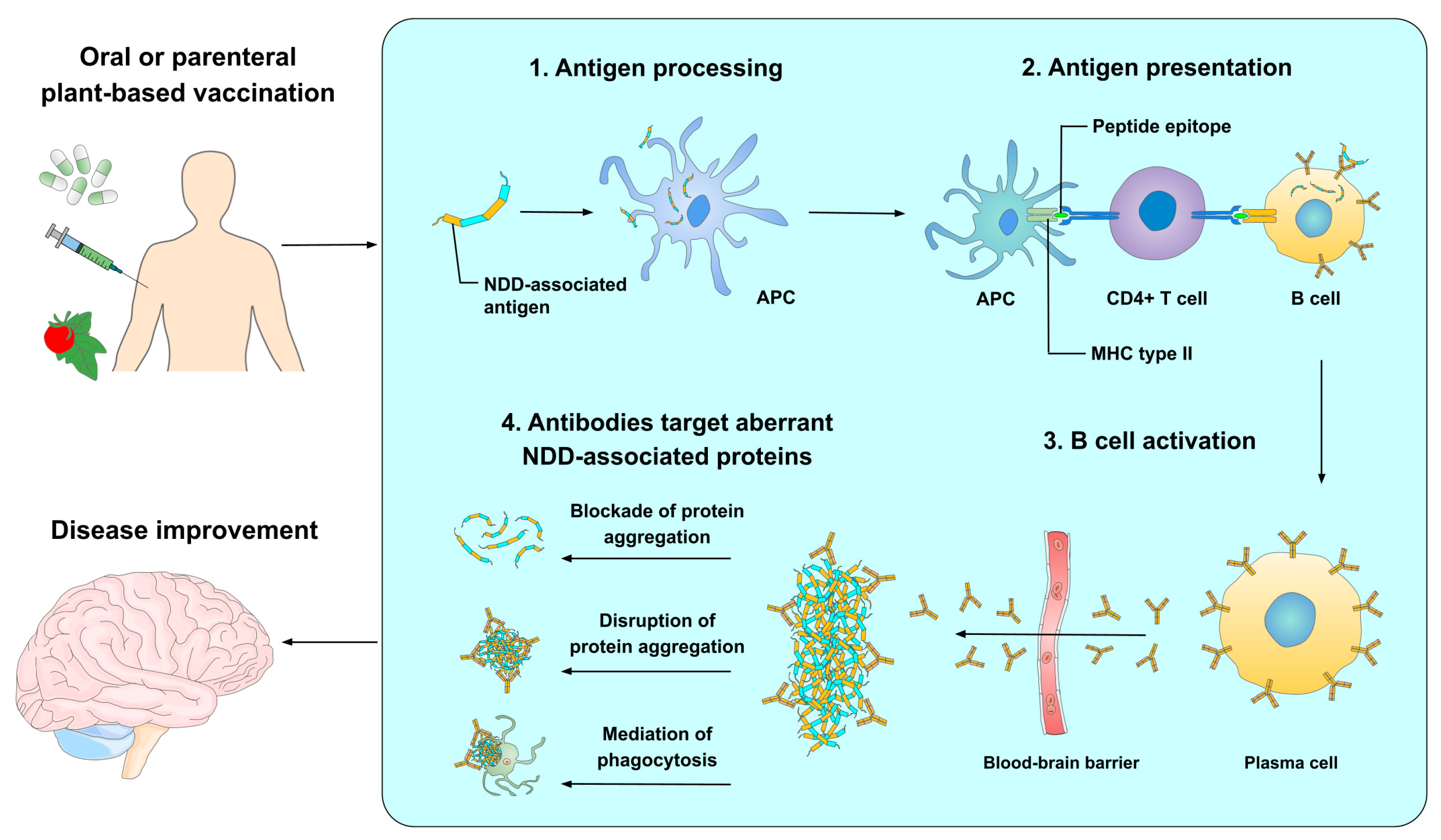
3. Mechanism of Action of Peptide-Based Vaccines for Neurodegenerative Diseases
4. Progress on Plant-Based Vaccines for Alzheimer’s Disease
5. Progress on Plant-Based Vaccines for Parkinson’s Disease
6. Progress on Plant-Based Vaccines for Multiple Sclerosis
7. Phytochemical-Based Treatments for Neurodegenerative Diseases
8. Concluding Remarks
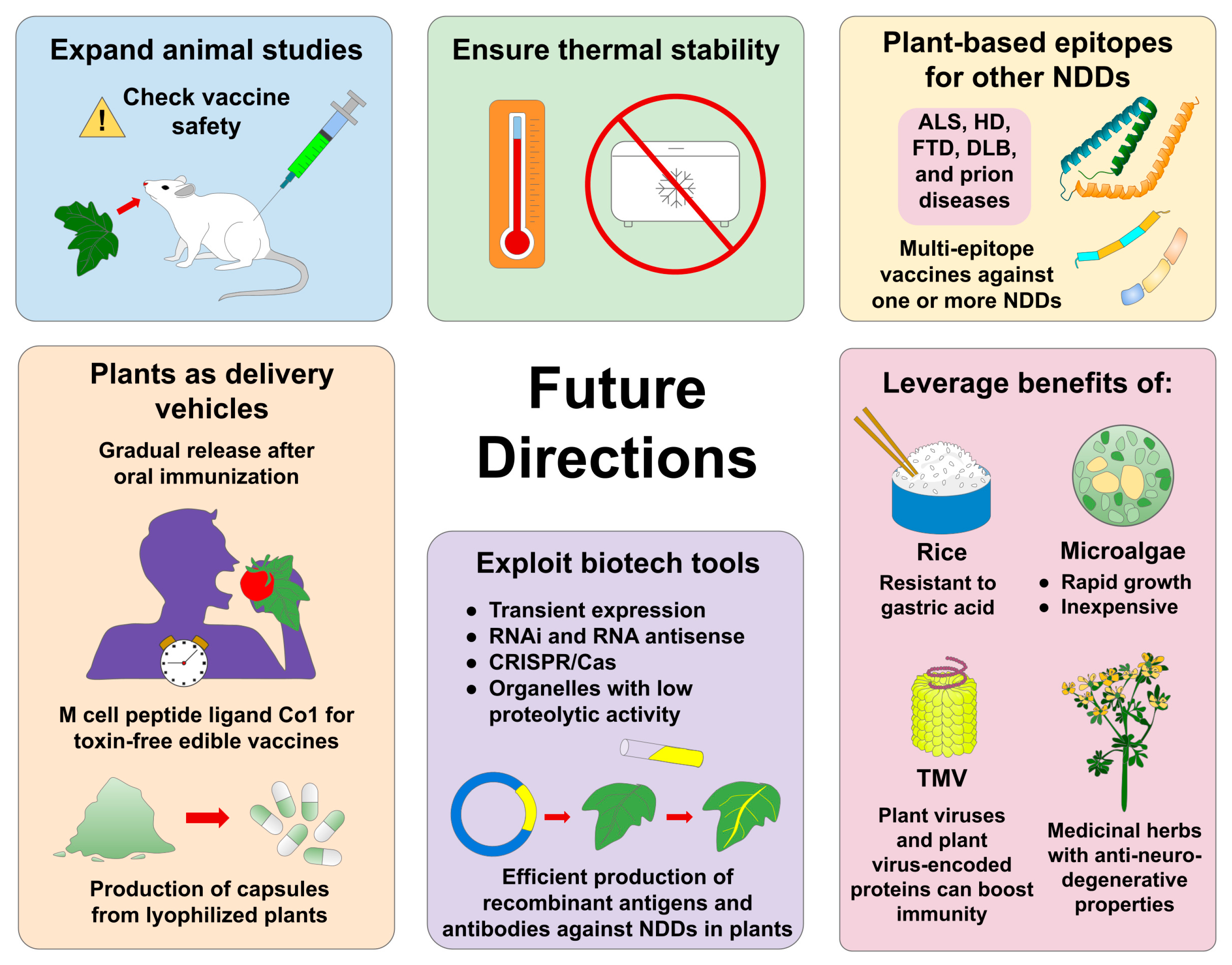
9. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di Paolo, M.; Papi, L.; Gori, F.; Turillazzi, E. Natural Products in Neurodegenerative Diseases: A Great Promise but an Ethical Challenge. Int. J. Mol. Sci. 2019, 20, 5170. [Google Scholar] [CrossRef] [Green Version]
- Amor, S.; Peferoen, L.A.N.; Vogel, D.Y.S.; Breur, M.; van der Valk, P.; Baker, D.; Van Noort, J.M. Inflammation in neurodegenerative diseases—An update. Immunology 2014, 142, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.; Ottolenghi, S.; Zuccotti, G.; Samaja, M.; Carelli, S. Mitochondrial dysfunctions in neurodegenerative diseases: Role in disease pathogenesis, strategies for analysis and therapeutic prospects. Neural Regen. Res. 2022, 17, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Aldewachi, H.; Al-Zidan, R.N.; Conner, M.T.; Salman, M.M. High-Throughput Screening Platforms in the Discovery of Novel Drugs for Neurodegenerative Diseases. Bioengineering 2021, 8, 30. [Google Scholar] [CrossRef]
- Schaffert, L.-N.; Carter, W.G. Do Post-Translational Modifications Influence Protein Aggregation in Neurodegenerative Diseases: A Systematic Review. Brain Sci. 2020, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, S.; Silva, B.; Long, S.; Pinto, M.; Remião, F.; Sousa, E.; Silva, R. Fiscalin Derivatives as Potential Neuroprotective Agents. Pharmaceutics 2022, 14, 1456. [Google Scholar] [CrossRef]
- Aza, A.; Gómez-Vela, M.; Badia, M.; Begoña Orgaz, M.; González-Ortega, E.; Vicario-Molina, I.; Montes-López, E. Listening to families with a person with neurodegenerative disease talk about their quality of life: Integrating quantitative and qualitative approaches. Health Qual. Life Outcomes 2022, 20, 76. [Google Scholar] [CrossRef]
- Galts, C.P.C.; Bettio, L.E.B.; Jewett, D.C.; Yang, C.C.; Brocardo, P.S.; Rodrigues, A.L.S.; Thacker, J.S.; Gil-Mohapel, J. Depression in neurodegenerative diseases: Common mechanisms and current treatment options. Neurosci. Biobehav. Rev. 2019, 102, 56–84. [Google Scholar] [CrossRef]
- Herzog-Krzywoszanska, R.; Krzywoszanski, L. Sleep Disorders in Huntington’s Disease. Front. Psychiatry 2019, 10, 221. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.N.; Belli, A.; Di Pietro, V. Small Non-coding RNAs: New Class of Biomarkers and Potential Therapeutic Targets in Neurodegenerative Disease. Front. Genet. 2019, 10, 364. [Google Scholar] [CrossRef]
- Adan, G.; Mitchell, J.W.; Ziso, B.; Larner, A.J. Diagnosis and Management of Seizures in Neurodegenerative Diseases. Curr. Treat. Options Neurol. 2021, 23, 1. [Google Scholar] [CrossRef]
- Doty, K.R.; Guillot-Sestier, M.V.; Town, T. The role of the immune system in neurodegenerative disorders: Adaptive or maladaptive? Brain Res. 2015, 1617, 155–173. [Google Scholar] [CrossRef] [Green Version]
- Hammond, T.R.; Marsh, S.E.; Stevens, B. Immune Signaling in Neurodegeneration. Immunity 2019, 50, 955–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, J.E.; Veasey, S.C. Impact of sleep disturbances on neurodegeneration: Insight from studies in animal models. Neurobiol. Dis. 2020, 139, 104820. [Google Scholar] [CrossRef] [PubMed]
- Popa-Wagner, A.; Dumitrascu, D.I.; Capitanescu, B.; Petcu, E.B.; Surugiu, R.; Fang, W.-H.; Dumbrava, D.-A. Dietary habits, lifestyle factors and neurodegenerative diseases. Neural Regen. Res. 2020, 15, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, S.; Rana, J.N.; Choi, E.H.; Han, I. Microwave Radiation and the Brain: Mechanisms, Current Status, and Future Prospects. Int. J. Mol. Sci. 2022, 23, 9288. [Google Scholar] [CrossRef]
- Ciccocioppo, F.; Bologna, G.; Ercolino, E.; Pierdomenico, L.; Simeone, P.; Lanuti, P.; Pieragostino, D.; Del Boccio, P.; Marchisio, M.; Miscia, S. Neurodegenerative diseases as proteinopathies-driven immune disorders. Neural Regen. Res. 2020, 15, 850–856. [Google Scholar] [CrossRef]
- Soto, C.; Pritzkow, S. Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1332–1340. [Google Scholar] [CrossRef]
- Marsh, A.P. Molecular mechanisms of proteinopathies across neurodegenerative disease: A review. Neurol. Res. Pract. 2019, 1, 35. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Bandopadhyay, R.; Singh, P.K.; Mishra, P.S.; Sharma, N.; Khurana, N. Neuroinflammation in neurological disorders: Pharmacotherapeutic targets from bench to bedside. Metab. Brain Dis. 2021, 36, 1591–1626. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, G.; Zhang, F. Role of Peripheral Immune Cells-Mediated Inflammation on the Process of Neurodegenerative Diseases. Front. Immunol. 2020, 11, 582825. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Tweedie, D.; Scerba, M.T.; Greig, N.H. Neuroinflammation as a Factor of Neurodegenerative Disease: Thalidomide Analogs as Treatments. Front. Cell Dev. Biol. 2019, 7, 313. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Calvani, R.; Coelho-Júnior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial Dysfunction, Oxidative Stress, and Neuroinflammation: Intertwined Roads to Neurodegeneration. Antioxidants 2020, 9, 647. [Google Scholar] [CrossRef]
- Sahebnasagh, A.; Eghbali, S.; Saghafi, F.; Sureda, A.; Avan, R. Neurohormetic phytochemicals in the pathogenesis of neurodegenerative diseases. Immun. Ageing 2022, 19, 36. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Bravo Vázquez, L.A.; Pérez Uribe, S.; Reyes-Pérez, P.R.; Sharma, A. Current Status of microRNA-Based Therapeutic Approaches in Neurodegenerative Disorders. Cells 2020, 9, 1698. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Kuo, H.C. Functional roles and networks of non-coding RNAs in the pathogenesis of neurodegenerative diseases. J. Biomed. Sci. 2020, 27, 49. [Google Scholar] [CrossRef] [Green Version]
- Pohl, F.; Kong Thoo Lin, P. The Potential Use of Plant Natural Products and Plant Extracts with Antioxidant Properties for the Prevention/Treatment of Neurodegenerative Diseases: In Vitro, In Vivo and Clinical Trials. Molecules 2018, 23, 3283. [Google Scholar] [CrossRef] [Green Version]
- Gorabi, A.M.; Kiaie, N.; Barreto, G.E.; Read, M.I.; Tafti, H.A.; Sahebkar, A. The Therapeutic Potential of Mesenchymal Stem Cell–Derived Exosomes in Treatment of Neurodegenerative Diseases. Mol. Neurobiol. 2019, 56, 8157–8167. [Google Scholar] [CrossRef]
- Shukla, S.; Tekwani, B.L. Histone Deacetylases Inhibitors in Neurodegenerative Diseases, Neuroprotection and Neuronal Differentiation. Front. Pharmacol. 2020, 11, 537. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, H.; Wei, D.; Zhang, X.; Wang, J.; Wu, X.; Chang, J. Mitochondria-targeted nanoparticles in treatment of neurodegenerative diseases. Exploration 2021, 1, 20210115. [Google Scholar] [CrossRef]
- Feng, Y.S.; Tan, Z.X.; Wu, L.Y.; Dong, F.; Zhang, F. The involvement of NLRP3 inflammasome in the treatment of neurodegenerative diseases. Biomed. Pharmacother. 2021, 138, 111428. [Google Scholar] [CrossRef] [PubMed]
- Mortada, I.; Farah, R.; Nabha, S.; Ojcius, D.M.; Fares, Y.; Almawi, W.Y.; Sadier, N.S. Immunotherapies for Neurodegenerative Diseases. Front. Neurol. 2021, 12, 654739. [Google Scholar] [CrossRef] [PubMed]
- Sim, K.-Y.; Im, K.-C.; Park, S.-G. The Functional Roles and Applications of Immunoglobulins in Neurodegenerative Disease. Int. J. Mol. Sci. 2020, 21, 5295. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Iba, M.; Kim, C.; Masliah, E. Immunotherapies for Aging-Related Neurodegenerative Diseases—Emerging Perspectives and New Targets. Neurotherapeutics 2020, 17, 935–954. [Google Scholar] [CrossRef]
- Knecht, L.; Folke, J.; Dodel, R.; Ross, J.A.; Albus, A. Alpha-synuclein Immunization Strategies for Synucleinopathies in Clinical Studies: A Biological Perspective. Neurotherapeutics 2022, 19, 1489–1502. [Google Scholar] [CrossRef]
- Vassilakopoulou, V.; Karachaliou, C.-E.; Evangelou, A.; Zikos, C.; Livaniou, E. Peptide-Based Vaccines for Neurodegenerative Diseases: Recent Endeavors and Future Perspectives. Vaccines 2021, 9, 1278. [Google Scholar] [CrossRef]
- Hovakimyan, A.; Antonyan, T.; Shabestari, S.K.; Svystun, O.; Chailyan, G.; Coburn, M.A.; Carlen-Jones, W.; Petrushina, I.; Chadarevian, J.P.; Zagorski, K.; et al. A MultiTEP platform-based epitope vaccine targeting the phosphatase activating domain (PAD) of tau: Therapeutic efficacy in PS19 mice. Sci. Rep. 2019, 9, 15455. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Hovakimyan, A.; Zagorski, K.; Antonyan, T.; Petrushina, I.; Davtyan, H.; Chailyan, G.; Hasselmann, J.; Iba, M.; Adame, A.; et al. Efficacy and immunogenicity of MultiTEP-based DNA vaccines targeting human α-synuclein: Prelude for IND enabling studies. npj Vaccines 2022, 7, 1. [Google Scholar] [CrossRef]
- Ji, M.; Xie, X.; Liu, D.; Lu, S.; Zhang, L.; Huang, Y.; Liu, R. Engineered hepatitis B core virus-like particle carrier for precise and personalized Alzheimer’s disease vaccine preparation via fixed-point coupling. Appl. Mater. Today 2020, 19, 100575. [Google Scholar] [CrossRef]
- Sha, S.; Xing, X.N.; Wang, T.; Li, Y.; Zhang, R.W.; Shen, X.L.; Cao, Y.P.; Qu, L. DNA vaccines targeting amyloid-β oligomer ameliorate cognitive deficits of aged APP/PS1/tau triple-transgenic mouse models of Alzheimer’s disease. Neural Regen. Res. 2022, 17, 2305–2310. [Google Scholar] [CrossRef] [PubMed]
- García-Arriaza, J.; Marín, M.Q.; Merchán-Rubira, J.; Mascaraque, S.M.; Medina, M.; Ávila, J.; Hernández, F.; Esteban, M. Tauopathy Analysis in P301S Mouse Model of Alzheimer Disease Immunized with DNA and MVA Poxvirus-Based Vaccines Expressing Human Full-Length 4R2N or 3RC Tau Proteins. Vaccines 2020, 8, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.J.; Thijssen, E.; van Brummelen, E.; van der Plas, J.L.; Radanovic, I.; Moerland, M.; Hsieh, E.; Groeneveld, G.J.; Dodart, J.C. A Randomized First-in-Human Study With UB-312, a UBITh® α-Synuclein Peptide Vaccine. Mov. Disord. 2022, 37, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Mantile, F.; Prisco, A. Vaccination against β-Amyloid as a Strategy for the Prevention of Alzheimer’s Disease. Biology 2020, 9, 425. [Google Scholar] [CrossRef] [PubMed]
- Govea-Alonso, D.O.; Cardineau, G.A.; Rosales-Mendoza, S. Principles of Plant-Based Vaccines. In Genetically Engineered Plants as a Source of Vaccines Against Wide Spread Diseases; Rosales-Mendoza, S., Ed.; Springer: New York, NY, USA, 2014; pp. 1–14. [Google Scholar]
- Rahimian, N.; Miraei, H.R.; Amiri, A.; Ebrahimi, M.S.; Nahand, J.S.; Tarrahimofrad, H.; Hamblin, M.R.; Khan, H.; Mirzaei, H. Plant-based vaccines and cancer therapy: Where are we now and where are we going? Pharmacol. Res. 2021, 169, 105655. [Google Scholar] [CrossRef]
- LeBlanc, Z.; Waterhouse, P.; Bally, J. Plant-Based Vaccines: The Way Ahead? Viruses 2021, 13, 5. [Google Scholar] [CrossRef]
- Dhama, K.; Natesan, S.; Iqbal Yatoo, M.; Patel, S.K.; Tiwari, R.; Saxena, S.K.; Harapan, H. Plant-based vaccines and antibodies to combat COVID-19: Current status and prospects. Hum. Vaccines Immunother. 2020, 16, 2913–2920. [Google Scholar] [CrossRef]
- Owczarek, B.; Gerszberg, A.; Hnatuszko-Konka, K. A Brief Reminder of Systems of Production and Chromatography-Based Recovery of Recombinant Protein Biopharmaceuticals. Biomed Res. Int. 2019, 2019, 4216060. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; Rubio-Infante, N.; Zarazúa, S.; Govea-Alonso, D.O.; Martel-Gallegos, G.; Moreno-Fierros, L. Plant-based vaccines for Alzheimer’s disease: An overview. Expert Rev. Vaccines 2014, 13, 429–441. [Google Scholar] [CrossRef]
- Ishiura, S.; Yoshida, T. Plant-based vaccines for Alzheimer’s disease. Proc. Jpn. Acad. Ser. B. Phys. Biol. Sci. 2019, 95, 290–294. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Euym, J.W.; Kim, M.S.; Lee, B.C.; Mook-Jung, I.; Jeon, J.H.; Joung, H. Expression of human β-amyloid peptide in transgenic potato. Plant Sci. 2003, 165, 1445–1451. [Google Scholar] [CrossRef]
- Youm, J.W.; Kim, H.; Han, J.H.L.; Jang, C.H.; Ha, H.J.; Mook-Jung, I.; Jeon, J.H.; Choi, C.Y.; Kim, Y.H.; Kim, H.S.; et al. Transgenic potato expressing Aβ reduce Aβ burden in Alzheimer’s disease mouse model. FEBS Lett. 2005, 579, 6737–6744. [Google Scholar] [CrossRef] [Green Version]
- Youm, J.W.; Jeon, J.H.; Kim, H.; Kim, Y.H.; Ko, K.; Joung, H.; Kim, H.S. Transgenic tomatoes expressing human beta-amyloid for use as a vaccine against Alzheimer’s disease. Biotechnol. Lett. 2008, 30, 1839–1845. [Google Scholar] [CrossRef] [Green Version]
- Arevalo-Villalobos, J.I.; Rosales-Mendoza, S.; Zarazua, S. Immunotherapies for neurodegenerative diseases: Current status and potential of plant-made biopharmaceuticals. Expert Rev. Vaccines 2017, 16, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Velázquez, A.; López-Quesada, A.; Ceballo-Cámara, Y.; Cabrera-Herrera, G.; Tiel-González, K.; Mirabal-Ortega, L.; Pérez-Martínez, M.; Pérez-Castillo, R.; Rosabal-Ayán, Y.; Ramos-González, O.; et al. Tobacco seeds as efficient production platform for a biologically active anti-HBsAg monoclonal antibody. Transgenic Res. 2015, 24, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Berlanga, B.; Pniewski, T. Plant-Based Vaccines in Combat against Coronavirus Diseases. Vaccines 2022, 10, 138. [Google Scholar] [CrossRef]
- Wong-Arce, A.; González-Ortega, O.; Rosales-Mendoza, S. Plant-Made Vaccines in the Fight Against Cancer. Trends Biotechnol. 2017, 35, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, S.; Hefferon, K.; Makhzoum, A.; Abouhaidar, M. Combating Human Viral Diseases: Will Plant-Based Vaccines Be the Answer? Vaccines 2021, 9, 761. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Rai, V.; Xiao, Y. Cold chain and virus-free oral polio booster vaccine made in lettuce chloroplasts confers protection against all three poliovirus serotypes. Plant Biotechnol. J. 2019, 17, 1357–1368. [Google Scholar] [CrossRef] [Green Version]
- Ward, B.J.; Séguin, A.; Couillard, J.; Trépanier, S.; Landry, N. Phase III: Randomized observer-blind trial to evaluate lot-to-lot consistency of a new plant-derived quadrivalent virus like particle influenza vaccine in adults 18–49 years of age. Vaccine 2021, 39, 1528–1533. [Google Scholar] [CrossRef]
- Paolino, K.M.; Regules, J.A.; Moon, J.E.; Ruck, R.C.; Bennett, J.W.; Remich, S.A.; Mills, K.T.; Lin, L.; Washington, C.N.; Fornillos, G.A.; et al. Safety and immunogenicity of a plant-derived recombinant protective antigen (rPA)-based vaccine against Bacillus anthracis: A Phase 1 dose-escalation study in healthy adults. Vaccine 2022, 40, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Frisio, D.G.; Ventura, V. Global Innovation Trends for Plant-Based Vaccines Production: A Patent Analysis. Plants 2021, 10, 2558. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.J.; Gobeil, P.; Séguin, A.; Atkins, J.; Boulay, I.; Charbonneau, P.Y.; Couture, M.; D’Aoust, M.A.; Dhaliwall, J.; Finkle, C.; et al. Phase 1 randomized trial of a plant-derived virus-like particle vaccine for COVID-19. Nat. Med. 2021, 27, 1071–1078. [Google Scholar] [CrossRef]
- Charland, N.; Gobeil, P.; Pillet, S.; Boulay, I.; Séguin, A.; Makarkov, A.; Heizer, G.; Bhutada, K.; Mahmood, A.; Trépanier, S.; et al. Safety and immunogenicity of an AS03-adjuvanted plant-based SARS-CoV-2 vaccine in Adults with and without Comorbidities. NPJ Vaccines 2022, 7, 142. [Google Scholar] [CrossRef]
- Hager, K.J.; Pérez Marc, G.; Gobeil, P.; Diaz, R.S.; Heizer, G.; Llapur, C.; Makarkov, A.I.; Vasconcellos, E.; Pillet, S.; Riera, F.; et al. Efficacy and Safety of a Recombinant Plant-Based Adjuvanted Covid-19 Vaccine. N. Engl. J. Med. 2022, 386, 2084–2096. [Google Scholar] [CrossRef]
- Kumar, M.; Kumari, N.; Thakur, N.; Bhatia, S.K.; Saratale, G.D.; Ghodake, G.; Mistry, B.M.; Alavilli, H.; Kishor, D.S.; Du, X.; et al. A Comprehensive Overview on the Production of Vaccines in Plant-Based Expression Systems and the Scope of Plant Biotechnology to Combat against SARS-CoV-2 Virus Pandemics. Plants 2021, 10, 1213. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Mendoza, S.; Salazar-González, J.A. Do microRNAs play a role in the activity of plant-based vaccines? Expert Rev. Vaccines 2017, 16, 529–533. [Google Scholar] [CrossRef]
- Nevagi, R.J.; Toth, I.; Skwarczynski, M. Peptide-based vaccines. In Peptide Applications in Biomedicine, Biotechnology and Bioengineering; Koutsopoulos, S., Ed.; Woodhead Publishing: Duxford, UK, 2018; pp. 327–358. [Google Scholar]
- Ringel, O.; Vieillard, V.; Debré, P.; Eichler, J.; Büning, H.; Dietrich, U. The Hard Way towards an Antibody-Based HIV-1 Env Vaccine: Lessons from Other Viruses. Viruses 2018, 10, 197. [Google Scholar] [CrossRef] [Green Version]
- Purcell, A.W.; McCluskey, J.; Rossjohn, J. More than one reason to rethink the use of peptides in vaccine design. Nat. Rev. Drug Discov. 2007, 6, 404–414. [Google Scholar] [CrossRef]
- Berger, A. Th1 and Th2 responses: What are they? BMJ 2000, 321, 424. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Timares, L.; Elmets, C.A. Host Defenses in Skin. In Clinical Immunology: Principles and Practice; Rich, R.R., Fleisher, T.A., Shearer, W.T., Schroeder, H.W., Jr., Frew, A.J., Weyand, C.M., Eds.; Elsevier: London, UK, 2019; pp. 273–283.e1. [Google Scholar]
- Marciani, D.J. Alzheimer’s disease vaccine development: A new strategy focusing on immune modulation. J. Neuroimmunol. 2015, 287, 54–63. [Google Scholar] [CrossRef]
- Marciani, D.J. A retrospective analysis of the Alzheimer’s disease vaccine progress—The critical need for new development strategies. J. Neurochem. 2016, 137, 687–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abudula, T.; Bhatt, K.; Eggermont, L.J.; O’Hare, N.; Memic, A.; Bencherif, S.A. Supramolecular Self-Assembled Peptide-Based Vaccines: Current State and Future Perspectives. Front. Chem. 2020, 8, 598160. [Google Scholar] [CrossRef]
- Valiukas, Z.; Ephraim, R.; Tangalakis, K.; Davidson, M.; Apostolopoulos, V.; Feehan, J. Immunotherapies for Alzheimer’s Disease—A Review. Vaccines 2022, 10, 1527. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-P.; Downs, D.; Huang, X.-P.; Da, H.; Fung, K.-M.; Tang, J. Amyloid-beta reduction by memapsin 2 (beta-secretase) immunization. FASEB J. 2007, 21, 3184–3196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.X.; Tian, Y.; Wang, Z.T.; Ma, Y.H.; Tan, L.; Yu, J.T. The Epidemiology of Alzheimer’s Disease Modifiable Risk Factors and Prevention. J. Prev. Alzheimer’s Dis. 2021, 8, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Au, R.; Mattke, S.; van der Flier, W.M.; Aisen, P.; Apostolova, L.; Chen, C.; Cho, M.; De Santi, S.; Gao, P.; et al. Designing the next-generation clinical care pathway for Alzheimer’s disease. Nat. Aging 2022, 2, 692–703. [Google Scholar] [CrossRef]
- Rahman, M.M.; Lendel, C. Extracellular protein components of amyloid plaques and their roles in Alzheimer’s disease pathology. Mol. Neurodegener. 2021, 16, 59. [Google Scholar] [CrossRef]
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer’s Disease. Mol. Psychiatry 2021, 26, 5481–5503. [Google Scholar] [CrossRef]
- Ishii-Katsuno, R.; Nakajima, A.; Katsuno, T.; Nojima, J.; Futai, E.; Sasagawa, N.; Yoshida, T.; Watanabe, Y.; Ishiura, S. Reduction of amyloid β-peptide accumulation in Tg2576 transgenic mice by oral vaccination. Biochem. Biophys. Res. Commun. 2010, 399, 593–599. [Google Scholar] [CrossRef]
- Stratmann, T. Cholera Toxin Subunit B as Adjuvant––An Accelerator in Protective Immunity and a Break in Autoimmunity. Vaccines 2015, 3, 579–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 2022, 22, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Youm, J.W.; Jeon, J.H.; Kim, H.; Min, S.R.; Kim, M.S.; Joung, H.; Jeong, W.J.; Kim, H.S. High-level expression of a human β-site APP cleaving enzyme in transgenic tobacco chloroplasts and its immunogenicity in mice. Transgenic Res. 2010, 19, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Vitti, A.; Piazzolla, G.; Condelli, V.; Nuzzaci, M.; Lanorte, M.T.; Boscia, D.; De Stradis, A.; Antonaci, S.; Piazzolla, P.; Tortorella, C. Cucumber mosaic virus as the expression system for a potential vaccine against Alzheimer’s disease. J. Virol. Methods 2010, 169, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Oono, Y.; Wakasa, Y.; Hirose, S.; Yang, L.; Sakuta, C.; Takaiwa, F. Analysis of ER stress in developing rice endosperm accumulating β-amyloid peptide. Plant Biotechnol. J. 2010, 8, 691–718. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.J.; Yang, J.Y.; Hsieh, H.L.; Wu, S.H. Processing bodies control the selective translation for optimal development of Arabidopsis young seedlings. Proc. Natl. Acad. Sci. USA 2019, 116, 6451–6456. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Kimura, E.; Koike, S.; Nojima, J.; Futai, E.; Sasagawa, N.; Watanabe, Y.; Ishiura, S. Transgenic Rice Expressing Amyloid β-peptide for Oral Immunization. Int. J. Biol. Sci. 2011, 7, 301–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nojima, J.; Maeda, A.; Aoki, S.; Suo, S.; Yanagihara, D.; Watanabe, Y.; Yoshida, T.; Ishiura, S. Effect of rice-expressed amyloid β in the Tg2576 Alzheimer’s disease transgenic mouse model. Vaccine 2011, 29, 6252–6258. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Y-Maze for Assessment of Spatial Working and Reference Memory in Mice. In Pre-Clinical Models. Methods in Molecular Biology; Guest, P.C., Ed.; Humana Press: New York, NY, USA, 2019; Volume 1916, pp. 105–111. [Google Scholar]
- Nojima, J.; Ishii-Katsuno, R.; Futai, E.; Sasagawa, N.; Watanabe, Y.; Yoshida, T.; Ishiura, S. Production of Anti-Amyloid β Antibodies in Mice Fed Rice Expressing Amyloid β. Biosci. Biotechnol. Biochem. 2011, 75, 396–400. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Youm, J.W.; Moon, K.B.; Ha, J.H.; Kim, Y.H.; Joung, H.; Jeon, J.H. Expression analysis of human β-secretase in transgenic tomato fruits. Protein Expr. Purif. 2012, 82, 125–131. [Google Scholar] [CrossRef]
- Maruyama, N.; Fujiwara, K.; Yokoyama, K.; Cabanos, C.; Hasegawa, H.; Takagi, K.; Nishizawa, K.; Uki, Y.; Kawarabayashi, T.; Shouji, M.; et al. Stable accumulation of seed storage proteins containing vaccine peptides in transgenic soybean seeds. J. Biosci. Bioeng. 2014, 118, 441–447. [Google Scholar] [CrossRef] [PubMed]
- McLaurin, J.; Cecal, R.; Kierstead, M.E.; Tian, X.; Phinney, A.L.; Manea, M.; French, J.E.; Lambermon, M.H.L.; Darabie, A.A.; Brown, M.E.; et al. Therapeutically effective antibodies against amyloid-β peptide target amyloid-β residues 4-10 and inhibit cytotoxicity and fibrillogenesis. Nat. Med. 2002, 8, 1263–1269. [Google Scholar] [CrossRef]
- Romero-Maldonado, A.; Monreal-Escalante, E.; Rosales-Mendoza, S. Expression in plants of two new antigens with implications in Alzheimer’s disease immunotherapy. Plant Cell Tissue Organ Cult. 2016, 126, 361–370. [Google Scholar] [CrossRef]
- Teixeira, A.F.; Fernandes, L.G.V.; Cavenague, M.F.; Takahashi, M.B.; Santos, J.C.; Passalia, F.J.; Daroz, B.B.; Kochi, L.T.; Vieira, M.L.; Nascimento, A.L.T.O. Adjuvanted leptospiral vaccines: Challenges and future development of new leptospirosis vaccines. Vaccine 2019, 37, 3961–3973. [Google Scholar] [CrossRef]
- Zeltins, A.; West, J.; Zabel, F.; El Turabi, A.; Balke, I.; Haas, S.; Maudrich, M.; Storni, F.; Engeroff, P.; Jennings, G.T.; et al. Incorporation of tetanus-epitope into virus-like particles achieves vaccine responses even in older recipients in models of psoriasis, Alzheimer’s and cat allergy. NPJ Vaccines 2017, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Castro, R.; Acero Galindo, G.; García Salcedo, Y.; Uribe Campero, L.; Vazquez Perez, V.; Carrillo-Tripp, M.; Gevorkian, G.; Gomez Lim, M.A. Plant-based chimeric HPV-virus-like particles bearing amyloid-β epitopes elicit antibodies able to recognize amyloid plaques in APP-tg mouse and Alzheimer’s disease brains. Inflammopharmacology 2018, 26, 817–827. [Google Scholar] [CrossRef]
- Uribe-Campero, L.; Monroy-García, A.; Durán-Meza, A.L.; Villagrana-Escareño, M.V.; Ruíz-García, J.; Hernández, J.; Núñez-Palenius, H.G.; Gómez-Lim, M.A. Plant-based porcine reproductive and respiratory syndrome virus VLPs induce an immune response in mice. Res. Vet. Sci. 2015, 102, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Berlanga, B.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Efficient Expression of an Alzheimer’s Disease Vaccine Candidate in the Microalga Schizochytrium sp. Using the Algevir System. Mol. Biotechnol. 2018, 60, 362–368. [Google Scholar] [CrossRef]
- Kawarabayashi, T.; Terakawa, T.; Takahashi, A.; Hasegawa, H.; Narita, S.; Sato, K.; Nakamura, T.; Seino, Y.; Hirohata, M.; Baba, N.; et al. Oral Immunization with Soybean Storage Protein Containing Amyloid-β 4-10 Prevents Spatial Learning Decline. J. Alzheimer’s Dis. 2019, 70, 487–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Watanabe, Y.; Ishiura, S. Production of the herb Ruta chalepensis L. expressing amyloid β-GFP fusion protein. Proc. Jpn. Acad. Ser. B. Phys. Biol. Sci. 2019, 95, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savica, R.; Boeve, B.F.; Mielke, M.M. When Do α-Synucleinopathies Start? An Epidemiological Timeline. JAMA Neurol. 2018, 75, 503–509. [Google Scholar] [CrossRef]
- Serratos, I.N.; Hernández-Pérez, E.; Campos, C.; Aschner, M.; Santamaría, A. An Update on the Critical Role of α-Synuclein in Parkinson’s Disease and Other Synucleinopathies: From Tissue to Cellular and Molecular Levels. Mol. Neurobiol. 2022, 59, 620–642. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Yin, D.; Zhao, H.; Zhang, L. The immunology of Parkinson’s disease. Semin. Immunopathol. 2022, 44, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Chopade, P.; Chopade, N.; Zhao, Z.; Mitragotri, S.; Liao, R.; Chandran Suja, V. Alzheimer’s and Parkinson’s disease therapies in the clinic. Bioeng. Transl. Med. 2023, 8, e10367. [Google Scholar] [CrossRef] [PubMed]
- Koga, S.; Sekiya, H.; Kondru, N.; Ross, O.A.; Dickson, D.W. Neuropathology and molecular diagnosis of Synucleinopathies. Mol. Neurodegener. 2021, 16, 83. [Google Scholar] [CrossRef]
- Mao, Q.; Qin, W.; Zhang, A.; Ye, N. Recent advances in dopaminergic strategies for the treatment of Parkinson’s disease. Acta Pharmacol. Sin. 2020, 41, 471–482. [Google Scholar] [CrossRef]
- Arevalo-Villalobos, J.I.; Govea-Alonso, D.O.; Monreal-Escalante, E.; Zarazúa, S.; Rosales-Mendoza, S. LTB-Syn: A recombinant immunogen for the development of plant-made vaccines against synucleinopathies. Planta 2017, 245, 1231–1239. [Google Scholar] [CrossRef]
- Arevalo-Villalobos, J.I.; Govea Alonso, D.O.; Rosales-Mendoza, S. Using carrot cells as biofactories and oral delivery vehicles of LTB-Syn: A low-cost vaccine candidate against synucleinopathies. J. Biotechnol. 2020, 309, 75–80. [Google Scholar] [CrossRef]
- Fox, J.L. First plant-made biologic approved. Nat. Biotechnol. 2012, 30, 472. [Google Scholar] [CrossRef]
- Hauser, S.L.; Cree, B.A.C. Treatment of Multiple Sclerosis: A Review. Am. J. Med. 2020, 133, 1380–1390.e2. [Google Scholar] [CrossRef]
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [Green Version]
- McGinley, M.P.; Goldschmidt, C.H.; Rae-Grant, A.D. Diagnosis and Treatment of Multiple Sclerosis: A Review. JAMA 2021, 325, 765–779. [Google Scholar] [CrossRef]
- Gholamzad, M.; Ebtekar, M.; Ardestani, M.S.; Azimi, M.; Mahmodi, Z.; Mousavi, M.J.; Aslani, S. A comprehensive review on the treatment approaches of multiple sclerosis: Currently and in the future. Inflamm. Res. 2019, 68, 25–38. [Google Scholar] [CrossRef]
- Arevalo-Villalobos, J.I.; Govea-Alonso, D.O.; Bañuelos-Hernández, B.; González-Ortega, O.; Zarazúa, S.; Rosales-Mendoza, S. Inducible expression of antigens in plants: A study focused on peptides related to multiple sclerosis immunotherapy. J. Biotechnol. 2020, 318, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, R.; Ji, E.; Kim, S.Y. Phytochemicals That Regulate Neurodegenerative Disease by Targeting Neurotrophins: A Comprehensive Review. Biomed Res. Int. 2015, 2015, 814068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surguchov, A.; Bernal, L.; Surguchev, A.A. Phytochemicals as Regulators of Genes Involved in Synucleinopathies. Biomolecules 2021, 11, 624. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, Y.; Chen, Z.; Leng, S.X. Connection between Systemic Inflammation and Neuroinflammation Underlies Neuroprotective Mechanism of Several Phytochemicals in Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2018, 2018, 1972714. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, M.; Nawaz, A.; Naz, F.; Ullah, F.; Sadiq, A.; Islam, Z.U. Phytochemicals-based Therapeutics against Alzheimer’s Disease: An Update. Curr. Top. Med. Chem. 2022, 22, 1811–1820. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Rathinasamy, B.; Lohanathan, B.P.; Thiyagarajan, V.; Weng, C.-F. Neuroprotective Role of Phytochemicals. Molecules 2018, 23, 2485. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, R.; Azam, S.; Cho, D.-Y.; Su-Kim, I.; Choi, D.-K. Natural Phytochemicals as Novel Therapeutic Strategies to Prevent and Treat Parkinson’s Disease: Current Knowledge and Future Perspectives. Oxid. Med. Cell. Longev. 2021, 2021, 6680935. [Google Scholar] [CrossRef]
- Mojaverrostami, S.; Bojnordi, M.N.; Ghasemi-Kasman, M.; Ebrahimzadeh, M.A.; Hamidabadi, H.G. A Review of Herbal Therapy in Multiple Sclerosis. Adv. Pharm. Bull. 2018, 8, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, R.V.; Kumar, J.R.; Egbuna, C.; Ifemeje, J.C. Phytochemicals as therapeutic interventions in neurodegenerative diseases. In Phytochemicals as Lead Compounds for New Drug Discovery; Egbuna, C., Kumar, S., Ifemeje, J.C., Ezzat, S.M., Kaliyaperumal, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 161–178. [Google Scholar]
- Zahedipour, F.; Hosseini, S.A.; Henney, N.C.; Barreto, G.E.; Sahebkar, A. Phytochemicals as inhibitors of tumor necrosis factor alpha and neuroinflammatory responses in neurodegenerative diseases. Neural Regen. Res. 2022, 17, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Darvish, E.; Narimani, F.; Moradi, S.Z.; Abbaszadeh, F.; Khan, H. The regulatory role of non-coding RNAs and their interactions with phytochemicals in neurodegenerative diseases: A systematic review. Brief. Funct. Genom. 2023, elac055. [Google Scholar] [CrossRef] [PubMed]
- Lofthouse, S. Immunological aspects of controlled antigen delivery. Adv. Drug Deliv. Rev. 2002, 54, 863–870. [Google Scholar] [CrossRef]
- Quakkelaar, E.D.; Melief, C.J.M. Experience with Synthetic Vaccines for Cancer and Persistent Virus Infections in Nonhuman Primates and Patients. Adv. Immunol. 2012, 114, 77–106. [Google Scholar] [CrossRef]
- Schillberg, S.; Finnern, R. Plant molecular farming for the production of valuable proteins—Critical evaluation of achievements and future challenges. J. Plant Physiol. 2021, 258–259, 153359. [Google Scholar] [CrossRef]
- Chung, Y.H.; Church, D.; Koellhoffer, E.C.; Osota, E.; Shukla, S.; Rybicki, E.P.; Pokorski, J.K.; Steinmetz, N.F. Integrating plant molecular farming and materials research for next-generation vaccines. Nat. Rev. Mater. 2022, 7, 372–388. [Google Scholar] [CrossRef]
- Gaobotse, G.; Venkataraman, S.; Mmereke, K.M.; Moustafa, K.; Hefferon, K.; Makhzoum, A. Recent Progress on Vaccines Produced in Transgenic Plants. Vaccines 2022, 10, 1861. [Google Scholar] [CrossRef]
- Massa, S.; Presenti, O.; Benvenuto, E. Engineering Plants for the Future: Farming with Value-Added Harvest. In Progress in Botany; Cánovas, F., Lüttge, U., Matyssek, R., Pretzsch, H., Eds.; Springer: Cham, Switzerland, 2018; pp. 65–108. [Google Scholar]
- Shanmugaraj, B.; Bulaon, C.J.I.; Malla, A.; Phoolcharoen, W. Biotechnological Insights on the Expression and Production of Antimicrobial Peptides in Plants. Molecules 2021, 26, 4032. [Google Scholar] [CrossRef]
- Klocko, A.L. Genetic Containment for Molecular Farming. Plants 2022, 11, 2436. [Google Scholar] [CrossRef]
- Kaur, M.; Manchanda, P.; Kalia, A.; Ahmed, F.K.; Nepovimova, E.; Kuca, K.; Abd-Elsalam, K.A. Agroinfiltration Mediated Scalable Transient Gene Expression in Genome Edited Crop Plants. Int. J. Mol. Sci. 2021, 22, 10882. [Google Scholar] [CrossRef] [PubMed]
- Karki, U.; Fang, H.; Guo, W.; Unnold-Cofre, C.; Xu, J. Cellular engineering of plant cells for improved therapeutic protein production. Plant Cell Rep. 2021, 40, 1087–1099. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Timko, M.P. Improving Protein Quantity and Quality—The Next Level of Plant Molecular Farming. Int. J. Mol. Sci. 2022, 23, 1326. [Google Scholar] [CrossRef]
- Dowlath, S.; Campbell, K.; Al-Barwani, F.; Young, V.L.; Young, S.L.; Walker, G.F.; Ward, V.K. Dry Formulation of Virus-Like Particles in Electrospun Nanofibers. Vaccines 2021, 9, 213. [Google Scholar] [CrossRef]
- Kim, E.; Attia, Z.; Cormet-Boyaka, E.; Boyaka, P.N. Toxin-Based Modulators for Regulation of Mucosal Immune Responses. In Mucosal Vaccines: Innovation for Preventing Infectious Diseases; Kiyono, H., Pascual, D.W., Eds.; Academic Press: London, UK, 2020; pp. 185–201. [Google Scholar]
- Kim, T.G.; Kim, M.Y.; Huy, N.X.; Kim, S.H.; Yang, M.S. M Cell-Targeting Ligand and Consensus Dengue Virus Envelope Protein Domain III Fusion Protein Production in Transgenic Rice Calli. Mol. Biotechnol. 2013, 54, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.Y.; Kim, M.Y. Evaluation of the oral immunogenicity of M cell-targeted tetravalent EDIII antigen for development of plant-based edible vaccine against dengue infection. Plant Cell Tissue Organ Cult. 2019, 137, 1–10. [Google Scholar] [CrossRef]
- Kiyono, H.; Yuki, Y.; Nakahashi-Ouchida, R.; Fujihashi, K. Mucosal vaccines: Wisdom from now and then. Int. Immunol. 2021, 33, 767–774. [Google Scholar] [CrossRef]
- Burnett, M.J.B.; Burnett, A.C. Therapeutic recombinant protein production in plants: Challenges and opportunities. Plants People Planet 2020, 2, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Specht, E.A.; Mayfield, S.P. Algae-based oral recombinant vaccines. Front. Microbiol. 2014, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Cid, R.; Bolívar, J. Platforms for production of protein-based vaccines: From classical to next-generation strategies. Biomolecules 2021, 11, 1072. [Google Scholar] [CrossRef]
- Chandran, R.; Abrahamse, H. Identifying Plant-Based Natural Medicine against Oxidative Stress and Neurodegenerative Disorders. Oxid. Med. Cell. Longev. 2020, 2020, 8648742. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Zhu, C.; Kittur, F.S.; He, M.; Arning, E.; Zhang, J.; Johnson, A.J.; Jawa, G.S.; Thomas, M.D.; Ding, T.T.; et al. A plant-based mutant huntingtin model-driven discovery of impaired expression of GTPCH and DHFR. Cell. Mol. Life Sci. 2022, 79, 553. [Google Scholar] [CrossRef]
- Nworji, O.F. Characterisation of Transgenic Tobacco Plants Expressing Synthetic Mouse Prion Protein. PhD Thesis, University of East London Health, Sport and Bioscience, London, UK, 2016. [Google Scholar]
- Trujillo, E.; Rosales-Mendoza, S.; Angulo, C. A multi-epitope plant-made chimeric protein (LTBentero) targeting common enteric pathogens is immunogenic in mice. Plant Mol. Biol. 2020, 102, 159–169. [Google Scholar] [CrossRef]
- Folke, J.; Ferreira, N.; Brudek, T.; Borghammer, P.; Van Den Berge, N. Passive Immunization in Alpha-Synuclein Preclinical Animal Models. Biomolecules 2022, 12, 168. [Google Scholar] [CrossRef]
- Valera, E.; Masliah, E. Immunotherapy for neurodegenerative diseases: Focus on α-synucleinopathies. Pharmacol. Ther. 2013, 138, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesco, G.; Lomoio, S. Pathophysiology of neurodegenerative diseases: An interplay among axonal transport failure, oxidative stress, and inflammation? Semin. Immunol. 2022, 59, 101628. [Google Scholar] [CrossRef]
- Parrocha, C.M.T.; Nowick, J.S. Current peptide vaccine and immunotherapy approaches against Alzheimer’s disease. Pept. Sci. 2023, 115, e24289. [Google Scholar] [CrossRef] [PubMed]
- Donini, M.; Marusic, C. Current state-of-the-art in plant-based antibody production systems. Biotechnol. Lett. 2019, 41, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Zahmanova, G.; Aljabali, A.A.; Takova, K.; Toneva, V.; Tambuwala, M.M.; Andonov, A.P.; Lukov, G.L.; Minkov, I. The Plant Viruses and Molecular Farming: How Beneficial They Might Be for Human and Animal Health? Int. J. Mol. Sci. 2023, 24, 1533. [Google Scholar] [CrossRef]
- Friedland, R.P.; Tedesco, J.M.; Wilson, A.C.; Atwood, C.S.; Smith, M.A.; Perry, G.; Zagorski, M.G. Antibodies to Potato Virus Y Bind the Amyloid β Peptide: Immunohistochemical and NMR studies. J. Biol. Chem. 2008, 283, 22550–22556. [Google Scholar] [CrossRef] [Green Version]
- Bamogo, P.K.A.; Brugidou, C.; Sérémé, D.; Tiendrébéogo, F.; Djigma, F.W.; Simpore, J.; Lacombe, S. Virus-based pharmaceutical production in plants: An opportunity to reduce health problems in Africa. Virol. J. 2019, 16, 167. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Vaishnav, R.A.; Roberts, A.M.; Friedland, R.P. Humans Have Antibodies against a Plant Virus: Evidence from Tobacco Mosaic Virus. PLoS ONE 2013, 8, e60621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Vaishnav, R.A.; Roberts, A.M.; Friedland, R.P. Parkinson disease, edible solanaceae, and tobacco mosaic virus. Ann. Neurol. 2014, 75, 162–163. [Google Scholar] [CrossRef]
- Gasanova, T.V.; Petukhova, N.V.; Ivanov, P.A. Chimeric particles of tobacco mosaic virus as a platform for the development of next-generation nanovaccines. Nanotechnologies Russ. 2016, 11, 227–236. [Google Scholar] [CrossRef]
- Gasanova, T.V.; Koroleva, A.A.; Skurat, E.V.; Ivanov, P.A. Complexes Formed via Bioconjugation of Genetically Modified TMV Particles with Conserved Influenza Antigen: Synthesis and Characterization. Biochem. Moscow 2020, 85, 224–233. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Lin, Y.; Pan, D. A multifaceted evaluation of microgliosis and differential cellular dysregulation of mammalian target of rapamycin signaling in neuronopathic Gaucher disease. Front. Mol. Neurosci. 2022, 15, 944883. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Plant Species | Disease-Related Protein/Epitope Expressed | Biological Model | Main Results | Reference |
|---|---|---|---|---|---|
| Alzheimer’s Disease | Pepper (Capiscum annuum L. var. angulosum) | Aβ42 | Tg2576 transgenic and WT B6 mice | Oral immunization prompted the production of anti- Aβ antibodies and the reduction of Aβ levels without inflammatory responses | [83] |
| Tobacco (Nicotiana tabacum L.) | BACE1 | BALB/c mice | Oral immunization triggered a mild production of primary anti-BACE1 antibodies | [86] | |
| Aβ1-15 and Aβ4-15 | - | CMV can be genetically engineered to produce prospective plant-derived vaccines against AD | [87] | ||
| Rice (Oryza sativa L.) | Aβ42 | - | Expression of recombinant Aβ peptide caused ER stress in rice | [88] | |
| Aβ42 | C57BL/6J mice | Oral immunization increased the serum anti-Aβ antibody titer even in those groups fed with boiled rice | [90] | ||
| Aβ42 | Tg2576 transgenic mice | Oral immunization increased the anti-Aβ antibody titer, decreased both intracerebral and serum Aβ levels, and improved mice memory without inflammatory responses | [91] | ||
| Aβ42 | WT B6 mice | Oral immunization triggered the production of anti-Aβ antibodies without inflammatory responses | [93] | ||
| Tomato (Solanum lycopersicum L.) | BACE1 | - | Recombinant BACE1 protein preserves its activity for long periods of storage at cold or room temperature, is stable in low acid conditions, and could be used to produce prospective plant-derived vaccines against AD | [94] | |
| Soybean (Glycine max [L.] Merr.) | Aβ epitope (FRHDSGY) | - | Utilizing seed storage proteins as carriers, the ER may be a promising organelle for the stable accumulation of disease-related peptides | [95] | |
| Tobacco (Nicotiana tabacum L.) | RAGE23-54 and Aβ42 | - | Chimeric proteins containing RAGE and/or Aβ epitopes can be produced in tobacco, maintaining their antigenic properties | [97] | |
| Lily (Lilium sp.) | Aβ1-6 | C57BL/6 and BALB/c mice | Immunization triggered the production of anti-Aβ antibodies | [99] | |
| Not specified | Aβ11-28 | C57BL/6J mice | Chimeric proteins containing the Aβ epitope triggered the production of anti-Aβ antibodies capable of detecting Aβ plaques in APP-tg mouse and AD brains without inflammatory responses | [100] | |
| Microalga Schizochytrium sp. | RAGE23-54 | - | Schizochytrium sp. is a reliable platform for the synthesis of thermostable recombinant proteins with an antigenic activity that could be used to produce prospective plant-derived vaccines against AD | [102] | |
| Soybean (Glycine max [L.] Merr.) | Aβ4-10 | TgCRND8 mice | Oral immunization triggered the production of anti-Aβ antibodies and prevented spatial learning decline without inflammatory responses | [103] | |
| Ruta chalepensis L. | Aβ42 | C57BL/6J mice | Oral immunization triggered the production of anti-Aβ antibodies and since R. chalepensis is rich in bioactive compounds, it could have synergetic effects as a plant-based vaccine system against AD | [104] | |
| Parkinson’s Disease | Tobacco (Nicotiana tabacum L.) | α-syn85-99, α-syn109-126, and α-syn126-140 | BALB/c mice | Oral immunization triggered anti-syn humoral responses targeting brain polypeptides, implying the production of antibodies against α-syn; this system could be used to produce plant-made vaccines against PD | [111] |
| Carrot (Daucus carota L.) | α-syn85-99, α-syn109-126, and α-syn126-140 | BALB/c mice | Oral immunization triggered anti-syn humoral responses; this system could be used to produce plant-made vaccines against PD | [112] | |
| Multiple Sclerosis | Tobacco (Nicotiana tabacum L.) | MS-2A (containing BV5S2, BV6S5, and BV13S1 peptides) | BALB/c mice | Oral immunization triggered anti-MS-2A humoral responses; this system could be used to produce plant-made vaccines against MS | [118] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo-Vázquez, L.A.; Mora-Hernández, E.O.; Rodríguez, A.L.; Sahare, P.; Bandyopadhyay, A.; Duttaroy, A.K.; Paul, S. Current Advances of Plant-Based Vaccines for Neurodegenerative Diseases. Pharmaceutics 2023, 15, 711. https://doi.org/10.3390/pharmaceutics15020711
Bravo-Vázquez LA, Mora-Hernández EO, Rodríguez AL, Sahare P, Bandyopadhyay A, Duttaroy AK, Paul S. Current Advances of Plant-Based Vaccines for Neurodegenerative Diseases. Pharmaceutics. 2023; 15(2):711. https://doi.org/10.3390/pharmaceutics15020711
Chicago/Turabian StyleBravo-Vázquez, Luis Alberto, Erick Octavio Mora-Hernández, Alma L. Rodríguez, Padmavati Sahare, Anindya Bandyopadhyay, Asim K. Duttaroy, and Sujay Paul. 2023. "Current Advances of Plant-Based Vaccines for Neurodegenerative Diseases" Pharmaceutics 15, no. 2: 711. https://doi.org/10.3390/pharmaceutics15020711
APA StyleBravo-Vázquez, L. A., Mora-Hernández, E. O., Rodríguez, A. L., Sahare, P., Bandyopadhyay, A., Duttaroy, A. K., & Paul, S. (2023). Current Advances of Plant-Based Vaccines for Neurodegenerative Diseases. Pharmaceutics, 15(2), 711. https://doi.org/10.3390/pharmaceutics15020711