
Application of DNA Replicons in Gene Therapy and Vaccine Development

Abstract
:1. Introduction
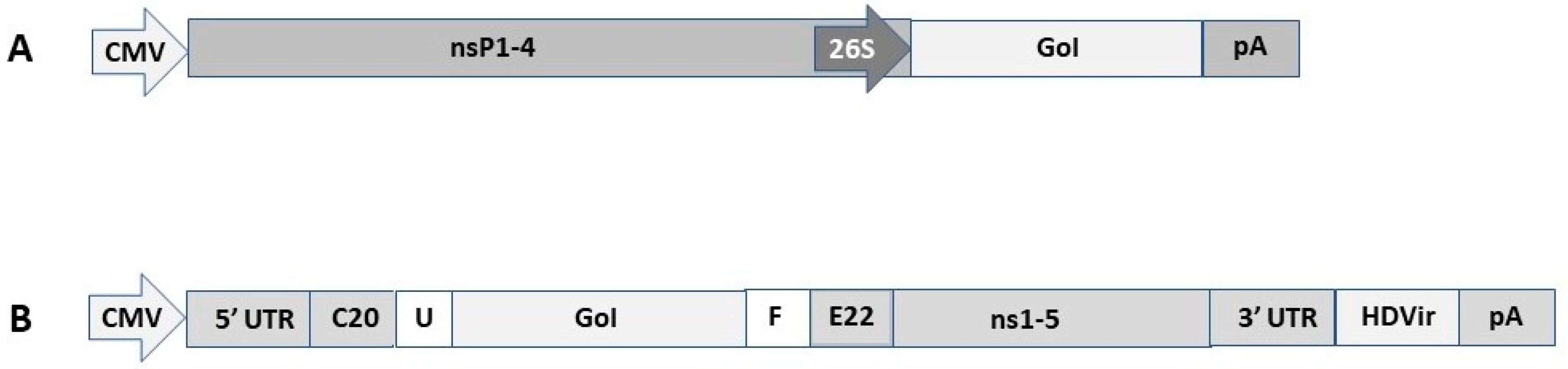
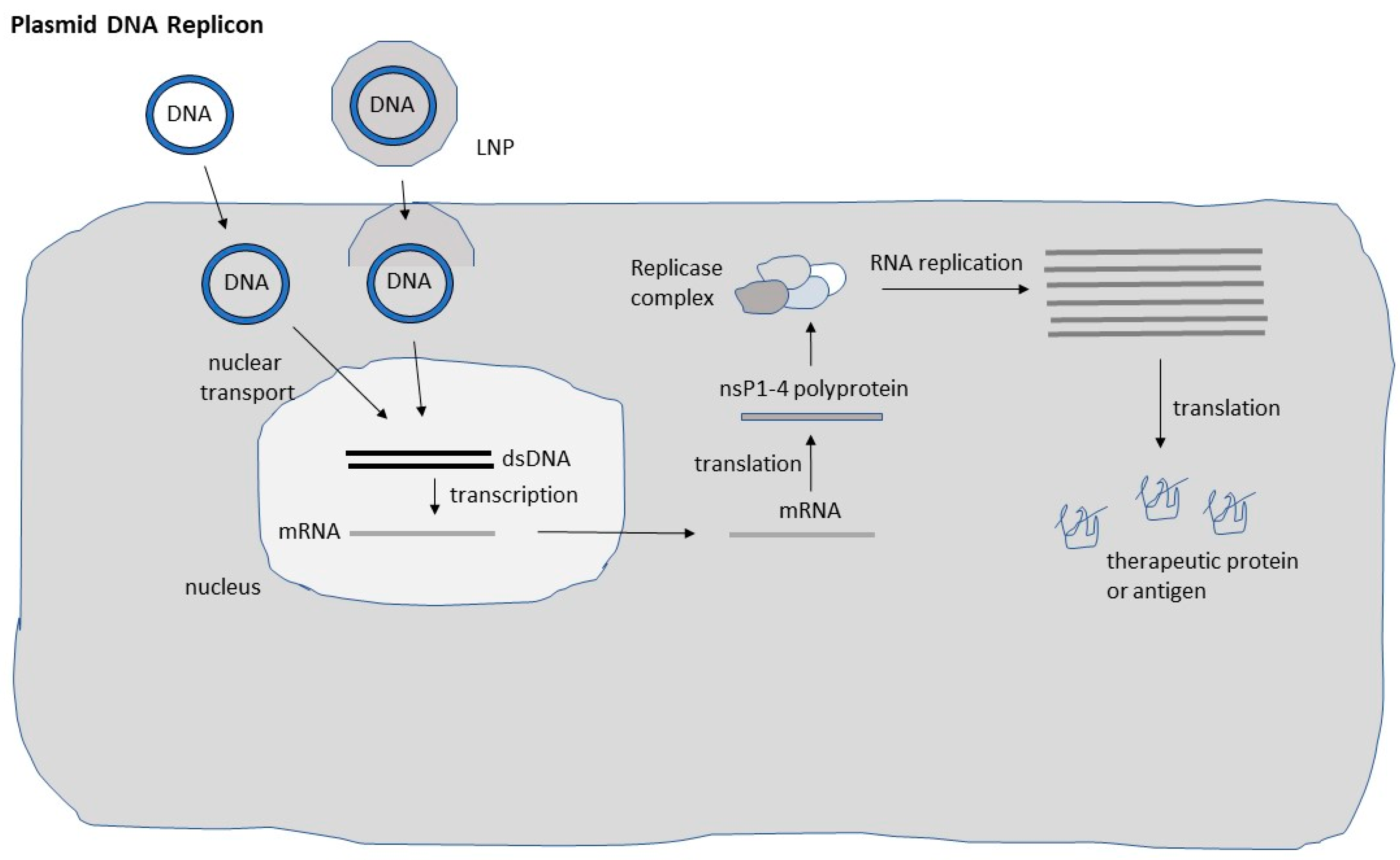
2. DNA Replicon Vectors
3. DNA Replicons and Infectious Diseases
4. DNA Replicons and Cancer
5. Comparison to Conventional DNA Vectors, RNA Replicons and Viral Particles
6. Conclusions
Funding
Conflicts of Interest
References
- Patil, S.D.; Rhodes, D.G.; Burges, D.J. DNA-based therapeutics and DNA delivery systems: A comprehensive review. AAPS J. 2005, 7, E61–E77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hager, S.; Fittler, F.J.; Wagner, E.; Bros, M. Nucleic-Acid Based Approaches for Tumor Therapy. Cells 2020, 9, 2061. [Google Scholar] [CrossRef] [PubMed]
- Vassaux, G.; Martin-Duque, P. Use of suicide genes for cancer gene therapy: Study of the different approaches. Expert Opin. Biol. Ther. 2004, 4, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Cirella, A.; Luri-Rey, C.; Di Trani, C.A.; Teijeira, A.; Olivera, I.; Bolanos, E.; Castanon, E.; Palencia, B.; Brocco, D.; Fernandez-Sendin, M.; et al. Novel strategies exploiting interleukin-12 in cancer immunotherapy. Pharmacol. Ther. 2022, 239, 108189. [Google Scholar] [CrossRef] [PubMed]
- Hodge, K.; Supabphol, S.; Kumar, P.; Poomipak, W.; Pisitkun, T. Recent developments in neoantigen-based cancer vaccines. Asian Pac. J. Allergy Immunol. 2020, 38, 91–101. [Google Scholar]
- Montano-Samaniego, M.; Bravo-Estupinan, D.; Mendez-Guerrero, O.; Alarcon-Hernandez, E.; Ibanez-Hernandez, M. Strategies for Targeting Gene Therapy in Cancer Cells with Tumor-Specific Promoters. Front. Oncol. 2020, 10, 605380. [Google Scholar] [CrossRef]
- Szczyrek, M.; Bitkowska, P.; Chunowski, P.; Czuchryta, P.; Krawwczyk, P.; Milanowski, J. Diet, Microbiome, and Cancer Therapy Immunotherapy—A Comprehensive Review. Nutrients 2021, 13, 2217. [Google Scholar] [CrossRef]
- Anderson, R.J.; Schneider, J. Plasmid DNA and viral vector-based vaccines for the treatment of cancer. Vaccine 2007, 25 (Suppl. S2), B24–B34. [Google Scholar] [CrossRef]
- Lee, J.; Arun Kumar, S.; Jhan, Y.Y.; Bishop, C.J. Engineering DNA vaccines against infectious diseases. Acta Biomater. 2018, 80, 31–47. [Google Scholar] [CrossRef]
- Marino, M.; Scuderi, F.; Provenzano, C.; Bartoccioni, E. Skeletal muscle cells: From local inflammatory response to active immunity. Gene Ther. 2011, 18, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Hengge, U.R.; Chan, E.F.; Foster, R.A.; Walker, P.S.; Vogel, J.C. Cytokine gene expression in epidermis with biological effects following injection of naked DNA. Nat. Genet. 1995, 10, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Lester, G.M.S.; Petishnok, L.C.; Dean, D.A. Cytoplasmic transport and nuclear import of plasmid DNA. Biosci. Rep. 2017, 37, BSR20160616. [Google Scholar] [CrossRef]
- Muramutsu, T.; Nakamura, A.; Park, H.M. In vivo electroporation: A powerful and convenient means of nonviral gene transfer to tissues of living cells. Int. J. Mol. Med. 1998, 1, 55–62. [Google Scholar]
- Mahvi, D.M.; Sheehy, M.J.; Yang, N.S. DNA cancer vaccines: A gene gun approach. Immunol. Cell Biol. 1997, 75, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K.; Boulikas, T. Viral and non-viral vectors in gene therapy. technology development and clinical trials. Technol. Cancer Res. Treat. 2003, 2, 471–486. [Google Scholar] [CrossRef] [Green Version]
- Sunshine, J.C.; Bishop, C.J.; Green, J.J. Advances in polymeric and inorganic vectors for nonviral nucleic acid delivery. Ther. Deliv. 2011, 2, 493–521. [Google Scholar] [CrossRef] [Green Version]
- Lundstrom, K. Self-Replicating RNA Viruses for Vaccine Development against Infectious Diseases and Cancer. Vaccines 2021, 9, 1187. [Google Scholar] [CrossRef]
- Lundstrom, K. Application of Viral Vectors for Vaccine Development with a Special Emphasis on COVID-19. Viruses 2020, 12, 1324. [Google Scholar] [CrossRef]
- Kaufman, H.L.; Kohlhapp, F.J.; Zloza, A. Oncolytic viruses: A new class of immunotherapy drugs. Nat. Rev. Drug Discov. 2015, 14, 642–662. [Google Scholar] [CrossRef]
- Liljestrom, P.; Garoff, H. A new generation of animal cell expression vectors based on the Semliki Forest virus replicon. Biotechnology 1991, 9, 1356–1361. [Google Scholar] [CrossRef]
- DiCiommo, D.P.; Bremner, R. Rapid, high level protein production using DNA-based Semliki Forest virus vectors. J. Biol. Chem. 1998, 273, 18060–18066. [Google Scholar] [CrossRef] [Green Version]
- Pijlman, G.P.; Suhrbier, A.; Khromykh, A.A. Kunjin virus replicons: An RNA-based, non-cytopathic viral vector system for protein production, vaccine and gene therapy applications. Expert Opin. Biol. Ther. 2006, 6, 134–145. [Google Scholar] [CrossRef] [Green Version]
- De Felipe, F. Skipping the co-expression problem: The new 2A “CHYSEL” technology. Genet. Vaccines Ther. 2004, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Lechardeur, D.; Lukacs, G.L. Nucleocytoplasmic transport of plasmid DNA: A perilous journey from the cytoplasm to the nucleus. Hum. Gene Ther. 2006, 17, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.A.; Ludtke, J.J.; Acsadi, G.; Williams, P.; Jani, A. Long-term persistence of plasmid DNA and foreign gene expression in mouse muscle. Hum. Mol. Genet. 1992, 1, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Troilo, P.J.; Wang, X.; Griffiths, T.G.; Pacchione, S.J.; Barnum, A.B.; Harper, L.B.; Pauley, C.J.; Niu, Z.; Denisova, L.; et al. Detection of integration of plasmid DNA into host genomic DNA following intramuscular injection and electroporation. Gene Ther. 2004, 11, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Manam, S.; Ledwith, B.J.; Barnum, A.B.; Troilo, P.J.; Pauley, C.J.; Harper, L.B.; Griffiths, T.G., 2nd; Niu, Z.; Denisova, L.; Follmer, T.T.; et al. Plasmid DNA vaccines: Tissue distribution and effects of DNA sequence, adjuvants and delivery method on integration into host DNA. Intervirology 2000, 43, 273–281. [Google Scholar] [CrossRef]
- Jiao, S.; Williams, P.; Berg, R.K.; Hodgeman, B.A.; Liu, L.; Repetto, G.; Wolff, J.A. Direct gene transfer into nonhuman primate myofibers in vivo. Hum. Gene Ther. 1992, 3, 21–33. [Google Scholar] [CrossRef]
- Mairhofer, J.; Lara, A.R. Advances in host and vector development for the production of plasmid DNA vaccines. Methods Mol. Biol. 2014, 1139, 505–541. [Google Scholar]
- Harvey, T.J.; Anraku, I.; Linedale, R.; Harrich, D.; Mackenzie, J.; Suhrbier, A.; Khromykh, A.A. Kunjin virus replicon vectors for human immunodeficiency virus vaccine development. J. Virol. 2003, 77, 7796–7803. [Google Scholar] [CrossRef] [Green Version]
- Mühlebach, M.D.; Hutzler, S. Development of Recombinant Measles Virus-Based Vaccines. Methods Mol. Biol. 2017, 1581, 151–168. [Google Scholar] [PubMed]
- Lyles, D.S.; Rupprecht, C.E. Rhabdoviridiae. In Fields’ Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1364–1408. [Google Scholar]
- Hariharan, M.J.; Driver, D.A.; Townsend, K.; Brumm, D.; Polo, J.M.; Belli, B.A.; Catton, D.J.; Hsu, D.; Mittelstaedt, D.; McCormack, J.E.; et al. DNA immunization against herpes simplex virus: Enhanced efficacy using a Sindbis virus-based vector. J. Virol. 1998, 72, 950–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasetti, M.F.; Ramirez, K.; Resendiz-Albor, A.; Ulmer, J.; Barry, E.M.; Levine, M.M. Sindbis virus-based measles DNA vaccines protect cotton rats against respiratory measles: Relevance of antibodies, mucosal and systemic antibody-secreting cells, memory B cells, and Th1-type cytokines as correlates of immunity. J. Virol. 2009, 83, 2789–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, J.R.; Kwang, J.; Varthakavi, V.; Lechtenberg, K.F.; Minocha, H.C. Semliki Forest virus vector carrying the bovine viral diarrhea virus NS3 (p80) cDNA induced immune responses in mice and expressed BVDV protein in mammalian cells. Comp. Immunol. Microbiol. Infect. Dis. 1999, 22, 231–246. [Google Scholar] [CrossRef]
- Sun, Y.; Li, N.; Li, H.Y.; Li, M.; Qiu, H.J. Enhanced immunity against classical swine fever in pigs induced by prime-boost immunization using an alphavirus replicon-vectored DNA vaccine and a recombinant adenovirus. Vet. Immunol. Immunopathol. 2010, 137, 20–27. [Google Scholar] [CrossRef]
- Knudsen, M.L.; Ljungberg, K.; Tatoud, R.; Weber, J.; Esteban, M.; Liljeström, P. Alphavirus replicon DNA expressing HIV antigens is an excellent prime for boosting with recombinant Ankara (MVA) or with HIV gp140 protein antigen. PLoS ONE 2015, 10, e0117042. [Google Scholar] [CrossRef]
- Marin, M.Q.; Perez, P.; Ljungberg, K.; Sorzano, C.O.S.; Gomez, C.E.; Liljeström, P.; Esteban, M.; Garcia-Arriaza, J. Potent Anti-Hepatitis C (HCV) T Cell Immune Responses Induced in Mice Vaccinated with DNA-launched RNA Replicons and MVA-HCV. J. Virol. 2019, 93, e00055-19. [Google Scholar] [CrossRef] [Green Version]
- Öhlund, P.; Garcia-Arriaza, J.; Zusinaite, E.; Szurgot, I.; Männik, A.; Kraus, A.; Ustav, M.; Merits, A.; Esteban, M.; Liljeström, P.; et al. DNA-launched RNA replicon vaccines induce potent anti-Ebolavirus immune responses that can be further improved by a recombinant MVA boost. Sci. Rep. 2018, 8, 12459. [Google Scholar] [CrossRef]
- Ren, S.; Wei, Q.; Cai, L.; Yang, X.; Xing, C.; Tan, F.; Leavenworth, J.W.; Liang, S.; Liu, W. Alphavirus Replicon DNA Vectors Expressing Ebola GP and VP40 Antigens Induce Humoral and Cellular Immune Responses in Mice. Front. Microbiol. 2018, 8, 2662. [Google Scholar] [CrossRef] [Green Version]
- Tretyakova, I.; Tibbens, A.; Jokinen, J.D.; Johnson, D.M.; Lukashevich, I.S.; Pushko, P. Novel DNA-launched Venezuelan equine encephalitis virus vaccine with rearranged genome. Vaccine 2019, 37, 3317–3325. [Google Scholar] [CrossRef]
- Tretyakova, I.; Plante, K.S.; Rossi, S.L.; Lawrence, W.S.; Peel, J.E.; Gudjohnsen, S.; Wang, E.; Mirchandani, D.; Tibbens, A.; Lamichhane, T.N.; et al. Venezuelan equine encephalitis vaccine with rearranged genome resists reversion and protects non-human primates from viremia after aerosol challenge. Vaccine 2020, 38, 3378–3386. [Google Scholar] [CrossRef] [PubMed]
- Szurgot, I.; Hanke, L.; Sheward, D.J.; Vidakovics, L.P.; Murrell, B.; McInerney, G.M.; Liljeström, P. DNA-launched replicon vaccines induce potent anti-SARS-CoV-2 immune responses in mice. Sci. Rep. 2021, 11, 3125. [Google Scholar] [CrossRef] [PubMed]
- Kirman, J.R.; Turon, T.; Su, H.; Li, A.; Kraus, C.; Polo, J.M.; Belisle, J.; Morris, S.; Seder, R.A. Enhanced immunogenicity to Mycobacterium tuberculosis by vaccination with an alphavirus plasmid replicon expressing antigen 85A. Infect. Immun. 2003, 71, 575–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmia, N.; Klimstra, W.B.; Mason, C.; Ramsay, A.J. DNA-launched alphavirus replicons encoding a fusion of mycobacterial antigens Acr and Ag85B are immunogenic and protective in a murine model of TB infection. PLoS ONE 2015, 10, e0136635. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Yu, Y.Z.; Yu, W.Y.; Sun, Z.-W. Enhancement of the immunogenicity of DNA replicon vaccine of Clostridium botulinum neurotoxin serotype A by GM-CSF gene adjuvant. Immunopharmacol. Immunotoxicol. 2011, 33, 211–219. [Google Scholar] [CrossRef]
- Yu, Y.-Z.; Guo, J.-P.; An, H.-J.; Zhang, S.M.; Wang, S.; Yu, W.Y.; Sun, Z.W. Potent tetravalent replicon vaccines against botulinum neurotoxins using DNA-based Semliki Forest virus replicon vectors. Vaccine 2012, 31, 2427–2432. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-Z.; Liu, S.; Ma, Y.; Gong, Z.-W.; Wang, S.; Sun, Z.-W. Pentavalent replicon vaccines against botulinum neurotoxins and tetanus toxin using DNA-based Semliki Forest virus replicon vectors. Hum. Vaccines Immunother. 2014, 10, 1874–1879. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-C.; An, H.-J.; Yu, Y.-Z.; Xu, Q. Potentiation of anthrax vaccines using protective antigen-expressing viral replicon vectors. Immunol. Lett. 2015, 163, 206–213. [Google Scholar] [CrossRef]
- Zheng, L.; Hu, Y.; Hua, Q.; Luo, F.; Xie, G.; Li, X.; Lin, J.; Wan, Y.; Ren, S.; Pan, C.; et al. Protective immune response in mice induced by a suicidal DNA vaccine encoding NTPase-II gene of toxoplasma gondii. Acta Trop. 2017, 166, 336–342. [Google Scholar] [CrossRef]
- Savar, N.S.; Vallet, T.; Azizi, M.; Arashkia, A.; Lundstrom, K.; Vignuzzi, M.; Niknam, H.M. Quantitative evaluation of PpSP15- LmSTI1 fusion gene expression following transfection with an alphavirus-derived self-amplifying mRNA and conventional DNA vaccine platforms. Mol. Cell. Probes 2021, 59, 101749. [Google Scholar] [CrossRef]
- Hsu, K.F.; Hung, C.F.; Cheng, W.F.; He, L.; Slater, L.A.; Ling, M.; Wu, T.C. Enhancement of suicidal DNA vaccine potency by linking Mycobacterium tuberculosis heat shock protein 70 to an antigen. Gene Ther. 2001, 8, 376–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Wall, S.; Ljungberg, K.; Ip, P.P.; Boerma, A.; Knudsen, M.L.; Nijman, H.W.; Liljeström, P.; Daemen, T. Potent therapeutic efficacy of an alphavirus replicon DNA vaccine expressing human papilloma virus E6 and E7 antigens. Oncoimmunology 2018, 7, e1487913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.W.; Hung, C.-F.; Juang, J.; He, L.; Hardwick, J.M.; Wu, T.-C. Enhancement of suicidal DNA vaccine potency by delaying suicidal DNA-induced cell death. Gene Ther. 2004, 11, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Lachman, L.B.; Rao, X.M.; Kremer, R.H.; Ozpolat, B.; Kiriakova, G.; Price, J.E. DNA vaccination against neu reduces breast cancer incidence and metastasis in mice. Cancer Gene Ther. 2001, 8, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eralp, Y.; Wang, X.; Wang, J.P.; Maughan, M.F.; Polo, J.M.; Lachman, L.B. Doxorubicin and paclitaxel enhance the antitumor efficacy of vaccines directed against HER2/neu in a murine mammary carcinoma model. Breast Cancer Res. 2004, 6, R275–R283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, J.P.; Rao, X.M.; Price, J.E.; Zhou, H.S.; Lachman, L.B. Prime-boost vaccination with plasmid and adenovirus gene vaccines control HER2/neu+ metastatic breast cancer in mice. Breast Cancer Res. 2005, 7, R580–R588. [Google Scholar] [CrossRef] [Green Version]
- Leitner, W.W.; Hwang, L.N.; deVeer, M.J.; Zhou, A.; Silverman, R.H.; Williams, B.R.G.; Dubensky, T.W.; Ying, H.; Restifo, N.P. Alphavirus-based DNA vaccine breaks immunological tolerance by activating innate antiviral pathways. Nat. Med. 2003, 9, 33–39. [Google Scholar] [CrossRef]
- Leslie, M.C.; Zhao, Y.J.; Lachman, L.B.; Hwu, P.; Wu, G.-J.; Bar-Eli, M. Immunization against MUC18/MCAM, a novel antigen that drives melanoma invasion and metastasis. Gene Ther. 2007, 14, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Wang, W.; Zhu, X.; Wang, Y.; Wu, S.; Wang, Z.; Wang, L.; Du, Z.; Gao, J.; Yu, J. Synergistic antitumor efficacy of combined DNA vaccines targeting tumor cells and angiogenesis. Biochem. Biophys. Res. Commun. 2015, 465, 239–244. [Google Scholar] [CrossRef]
- Varnavski, A.N.; Young, P.R.; Khromykh, A.A. Stable high-level expression of heterologous genes in vitro and in vivo by noncytopathic DNA-based Kunjin virus replicon vectors. J. Virol. 2000, 74, 4394–4403. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, R.; Xanthopoulos, K.G. Induction of antigen-specific immune responses against malignant brain tumors by intramuscular injection of Sindbis DNA encoding gp100 and IL-18. DNA Cell Biol. 2005, 24, 317–324. [Google Scholar] [CrossRef]
- Faham, A.; Herringson, T.; Parish, C.; Suhrbier, A.; Khromykh, A.A.; Altin, J.G. pDNA-lipoplexes engrafted with flagellin-related peptide induce potent immunity and anti-tumour effects. Vaccine 2011, 29, 6911–6919. [Google Scholar] [CrossRef] [PubMed]
- Berglund, P.; Smerdou, C.; Fleeton, M.N.; Tubulekas, I.; Liljeström, P. Enhancing immune responses using suicidal DNA vaccines. Nat. Biotechnol. 1998, 16, 562–565. [Google Scholar] [CrossRef]
- Xiao, S.; Chen, H.; Fang, L.; Liu, C.; Zhang, H.; Jiang, Y.; Hong, W. Comparison of immune responses and protective efficacy of suicidal DNA vaccine and conventional DNA vaccine encoding glycoprotein C of pseudorabies virus in mice. Vaccine 2004, 22, 345–351. [Google Scholar] [CrossRef]
- Kamrud, K.I.; Hooper, J.W.; Elgh, F.; Schmaljohn, C.S. Comparison of the protective efficacy of naked DNA, DNA-based Sindbis replicon, and packaged Sindbis replicon vectors expressing Hantavirus structural genes in hamsters. Virology 1999, 263, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, Y.N. BNT162b2 mRNA COVID-19 Vaccine: First Approval. Drugs 2021, 81, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.; Zaks, T.Z.; Wang, R.-F.; Irvine, K.R.; Kammula, U.S.; Marincola, F.M.; Leitner, W.W.; Restifo, N. Cancer therapy using a self-replicating RNA vaccine. Nat. Med. 1999, 5, 823–827. [Google Scholar] [CrossRef]
- Saxena, S.; Sonwane, A.A.; Dahiya, S.S.; Patel, C.L.; Saini, M.; Rai, A.; Gupta, P.K. Induction of immune responses and protection in mice against rabies using a self-replicating RNA vaccine encoding rabies virus glycoprotein. Vet. Microbiol. 2009, 136, 36–44. [Google Scholar] [CrossRef]
- Geall, A.J.; Verma, A.; Otten, G.R.; Shaw, G.A.; Hekele, A.; Banerjee, K.; Cu, Y.; Beard, C.W.; Brito, L.A.; Krucker, T.; et al. Nonviral delivery of self-amplifying RNA vaccines. Proc. Natl. Acad. Sci. USA 2012, 109, 14604–14609. [Google Scholar] [CrossRef] [Green Version]
- Henao-Restrepo, A.M.; Longini, I.M.; Egger, M.; Dean, N.E.; Edmunds, W.J.; Camacho, A.; Carroll, M.W.; Doumbia, M.; Draguez, B.; Duraffour, S.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine expressing Ebola surface glycoprotein: Interim results from the Guinea ring vaccination cluster-randomised trial. Lancet 2015, 386, 857–866. [Google Scholar] [CrossRef]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ca Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollmann Saphire, E. A vaccine against Ebola virus. Cell 2020, 181, 6. [Google Scholar] [CrossRef] [PubMed]
- Pollock, K.M.; Cheeseman, H.; Szubert, A.J.; Libri, V.; Boffito, M.; Owen, D.; Bern, H.; O’Hara, J.; McFarlane, L.R.; Lemm, N.M.; et al. Safety and immunogenicity of a self-amplifying RNA vaccine against COVID-19: COVAC1, a phase I, dose-ranging trial. EClinicalMedicine 2022, 44, 101262. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Disease | DNA Vector/Target | μg DNA | Findings |
|---|---|---|---|
| Viral | |||
| HSV-1 | SIN/HSV-1 gB | 0.01–3 | Protection in mice against HSV-1 after single injection [33] |
| MV | SIN/MV-H | 100 | 100% protection against MV in cotton rats [34] |
| MV | SIN/MV-H-Fdu | 100 | Protection only after booster with live MV vaccine in rats [34] |
| BVDV | SFV/BVDV p80 | 100 | CTL and CMI responses against BVDV in mice [35] |
| CSFV | SFV/CSFV-E2 + rAdV-E2 | 100 | Heterologous prime-boost strategy superior in pig model [36] |
| HIV | SFV/HIV Env, GagPolNef | 0.2 | Greatly enhanced immune responses after MVA booster [37] |
| HCV | SFV/HCV C, E1, E2, NS2/3 | 0.5–50 | CTL and humoral responses after MVA booster in mice [38] |
| EBOV | SFV/EBOV GP, VP40 | 5 | Binding and neutralizing Abs in mice [39] |
| EBOV | SFV/EBOV GP, VP40 | 10 | Superior humoral, cellular responses after co-injection [40] |
| VEE | VEE/V4020 genome | 100 | Protection against VEE in mice [41] |
| VEE | VEE/V4020 genome | 100 | Protection against VEE in rhesus macaques [42] |
| COVID-19 | SFV/SARS-CoV-2 S | 10 | Neutralizing Abs, superior IgG Abs in mice after 1 injection [43] |
| COVID-19 | SFV/SARS-CoV-2 Secto | 10 | Neutralizing Abs in mice [43] |
| Bacterial | |||
| TB | SIN/M. tuberculosis p85 | 0.5–50 | Specific Ab responses, protection against challenges in mice [44] |
| TB | VEE/Acr-Ag85B fusion | 20 | Inhibition of bacterial growth in lungs and spleen of mice [45] |
| Botulism | SFV/BoNT/A Hc, GM-CSF | 100 | Prolonged survival after BoNT/A challenges in mice [46] |
| Botulism | SFV/BoNT/E, BoNT/F | 100 | Protection against challenge with BoNT/E-BoNT/F mixture [47] |
| Botulism | SFV/BoNT/A, B, E, F | 100 | Protection against 4 BoNT serotypes in mice [48] |
| Tetanus | SFV/TeNT | 100 | Protection against TeNT in mice [48] |
| Anthrax | SFV/anthrax PA | 100 | Protection against B. anthracis A16R strain in mice [49] |
| Protozoan | |||
| TP | SFV/Tg-NPase II | 100 | Protection against acute infection, toxoplasmosis in mice [50] |
| Parasites | |||
| LD | SFV/PpSP15-LmST11 | 0.5–2 | Superior expression from RNA than DNA replicons [51] |
| Cancer | DNA Vector/Target | μg DNA | Findings |
|---|---|---|---|
| Cervical | SFV/HPV E7-Hsp70 | 2 | Potential antitumor activity in metastases [52] |
| Cervical | SFV/HPV E6-E7 | 0.05 | 85% of treated mice becoming tumor-free [53] |
| Cervical | SFV/HPV E7-BCL-xL | 2–20 | Higher immunogenicity, enhanced antitumor activity in mice [54] |
| Breast | SFV/HER2/neu | 100 | Tumor regression, protection against tumors in mice [55] |
| Breast | |||
| Breast | SIN/neu + Dox/Pac | 100 | Substantial tumor regression with Dox or Pac in mice [56] |
| SIN/neu + Ad-neu | 100 | Booster vaccination with Ad-neu prolonged survival in mice [57] | |
| Melanoma | SIN/TRP1 | 3 | Break of tolerance, immunity against melanomas in mice [58] |
| Melanoma | SIN/MCAM/MUC18 | 50 | Protection against B16F10 melanoma challenges in mice [59] |
| Melanoma | SFV/VEGFR2/IL-12 + Survivin/β-hCG Ag | 50 | Enhanced humoral, cellular immune responses, prolonged survival after co-administration of DNA replicons in mice [60] |
| Melanoma | KUN/SIIINFEKL-LPX | 25 | Enhanced antitumor activity in mice [61] |
| Brain | SIN/gp100/IL-18 | 100 | Protective, therapeutic effects, prolonged survival in mice [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lundstrom, K. Application of DNA Replicons in Gene Therapy and Vaccine Development. Pharmaceutics 2023, 15, 947. https://doi.org/10.3390/pharmaceutics15030947
Lundstrom K. Application of DNA Replicons in Gene Therapy and Vaccine Development. Pharmaceutics. 2023; 15(3):947. https://doi.org/10.3390/pharmaceutics15030947
Chicago/Turabian StyleLundstrom, Kenneth. 2023. "Application of DNA Replicons in Gene Therapy and Vaccine Development" Pharmaceutics 15, no. 3: 947. https://doi.org/10.3390/pharmaceutics15030947
APA StyleLundstrom, K. (2023). Application of DNA Replicons in Gene Therapy and Vaccine Development. Pharmaceutics, 15(3), 947. https://doi.org/10.3390/pharmaceutics15030947