

Chemical Constituents from the Roots of Angelica reflexa That Improve Glucose-Stimulated Insulin Secretion by Regulating Pancreatic β-Cell Metabolism


 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Plant Material
2.3. Genetic Analysis
2.4. Extraction and Isolation
2.4.1. Koseonolin A (1)
2.4.2. Koseonolin B (2)
2.4.3. Isohydroxylomatin (3)
2.5. Computational Methods
2.6. Cell Culture and Cell Viability Assay
2.7. GSIS Assay and ADP/ATP Ratio Assay
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
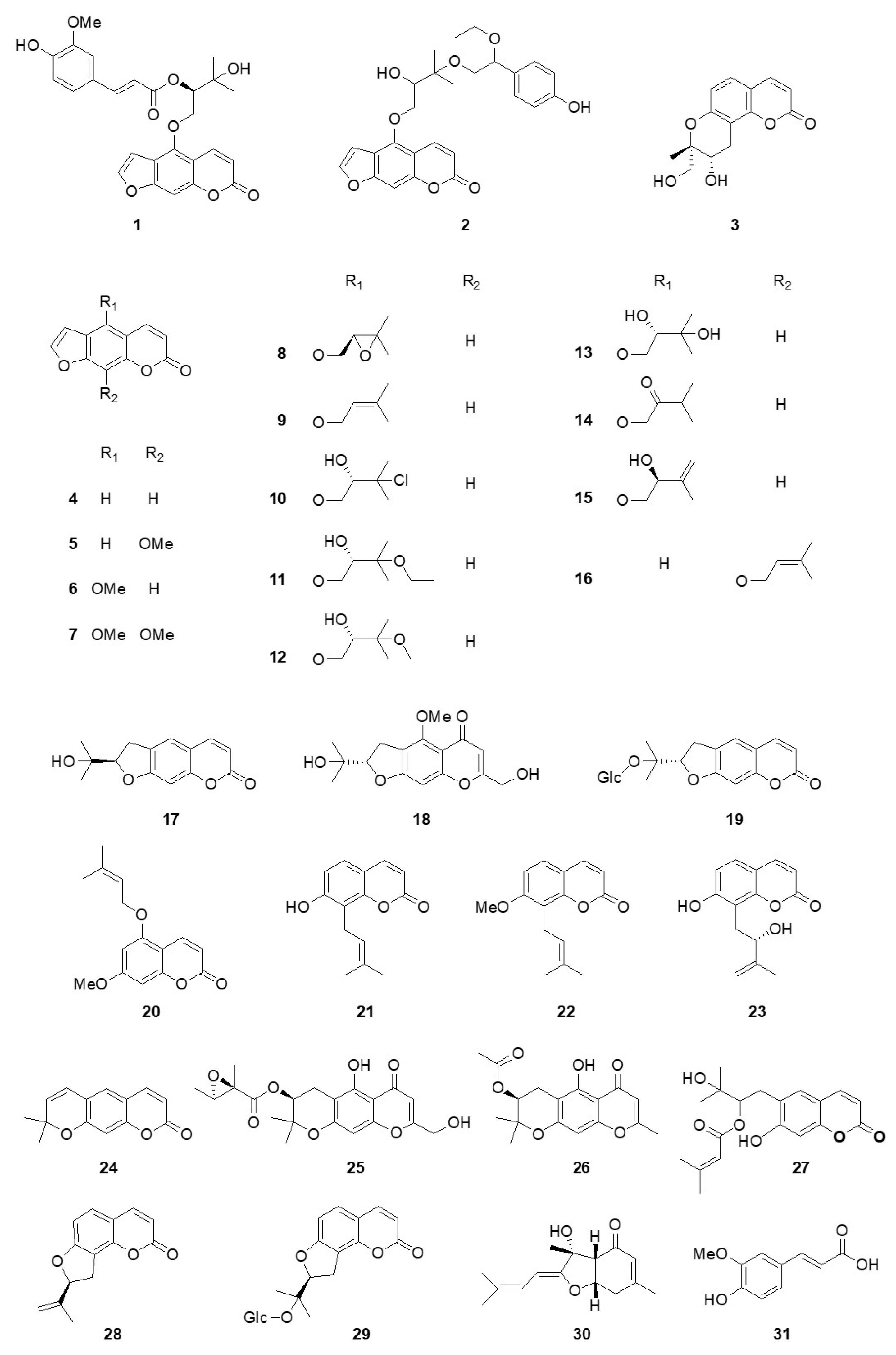
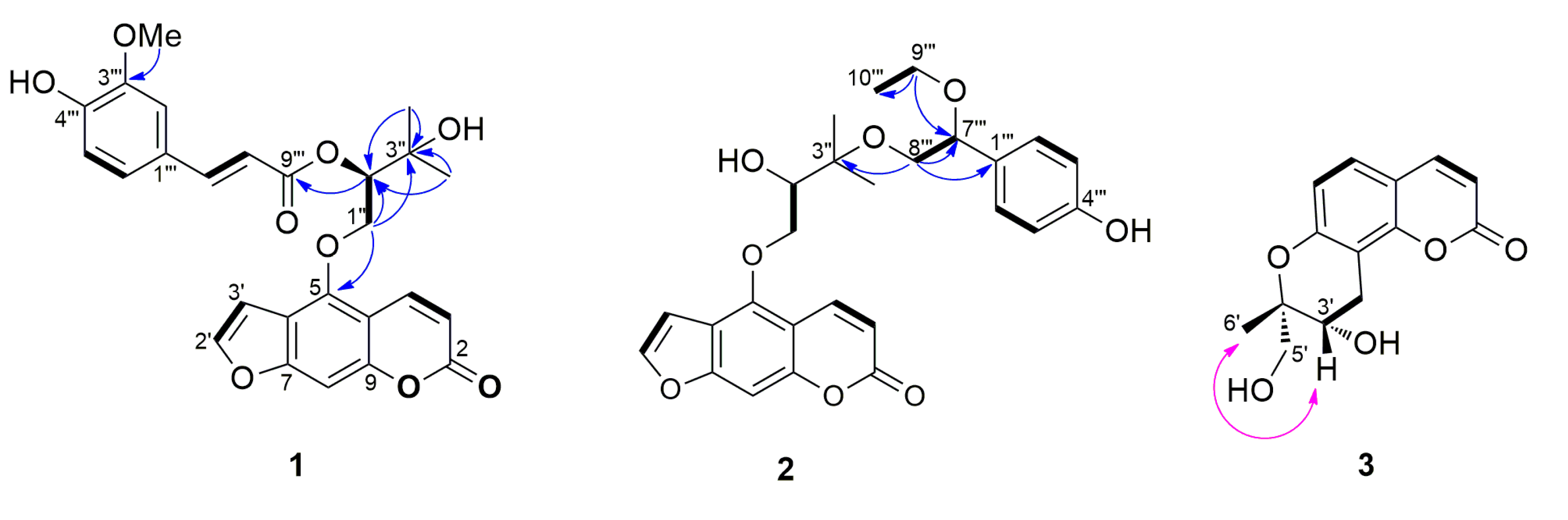
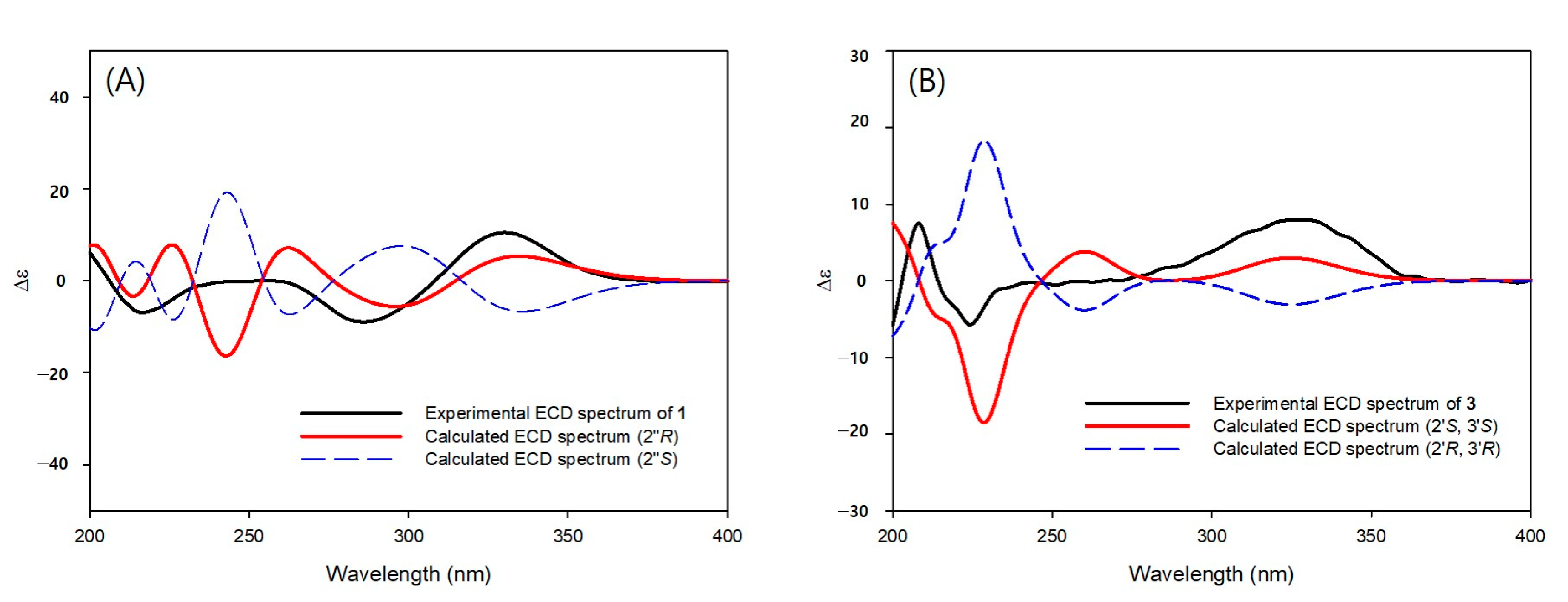
3.1. Structural Elucidation of Isolated Compounds
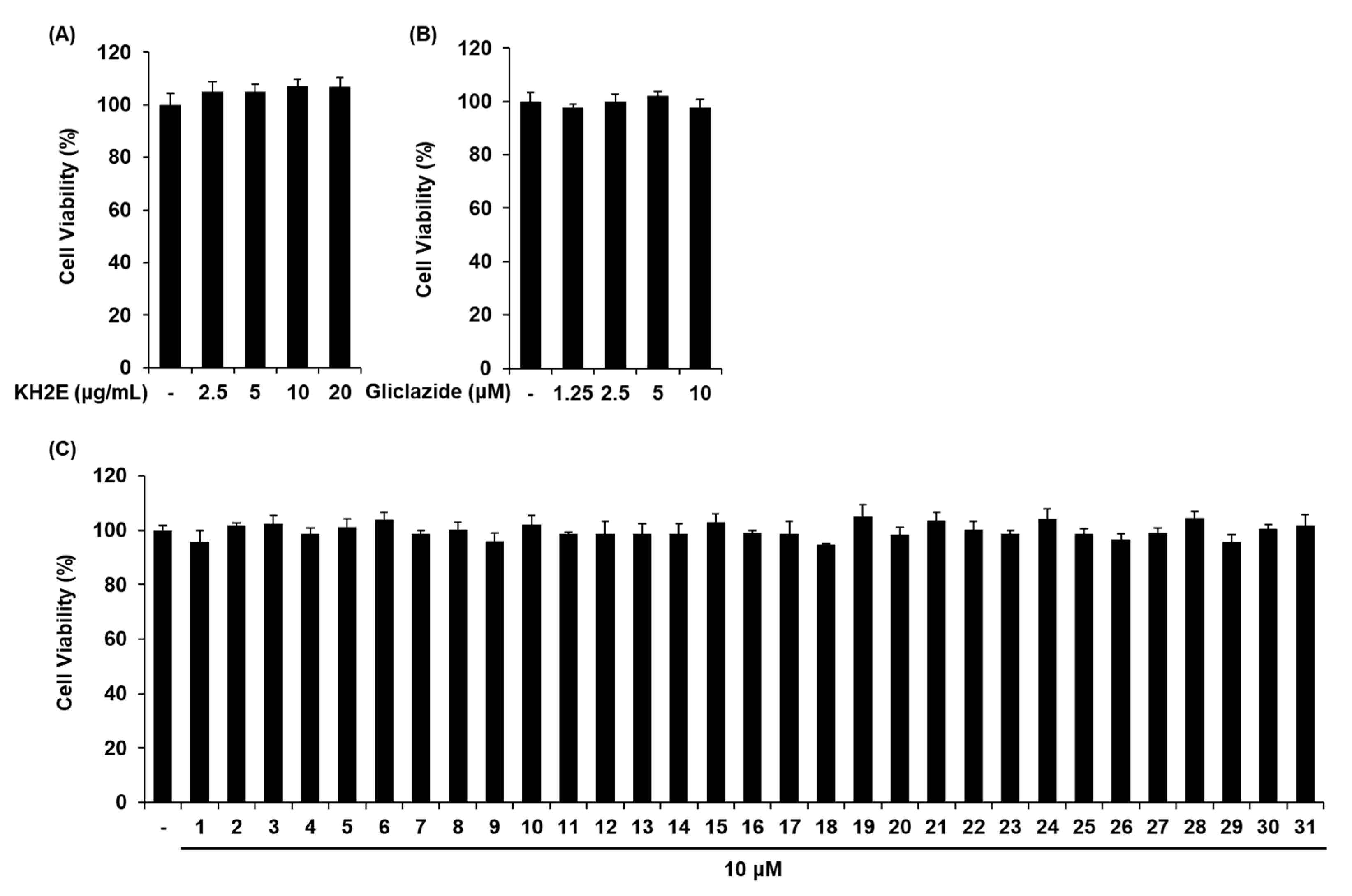
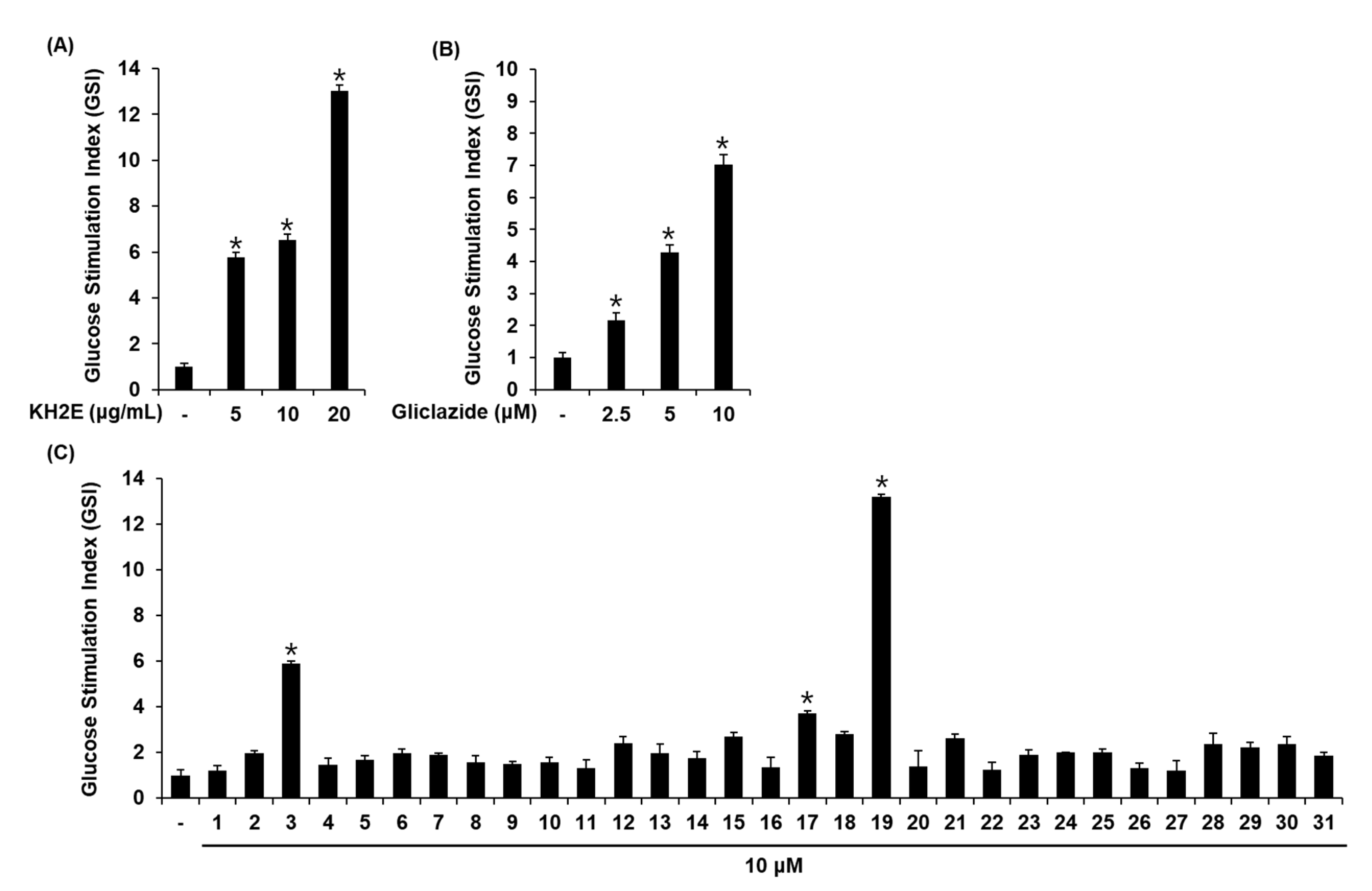
3.2. Effect of KH2E and Compounds 1–31 on GSIS
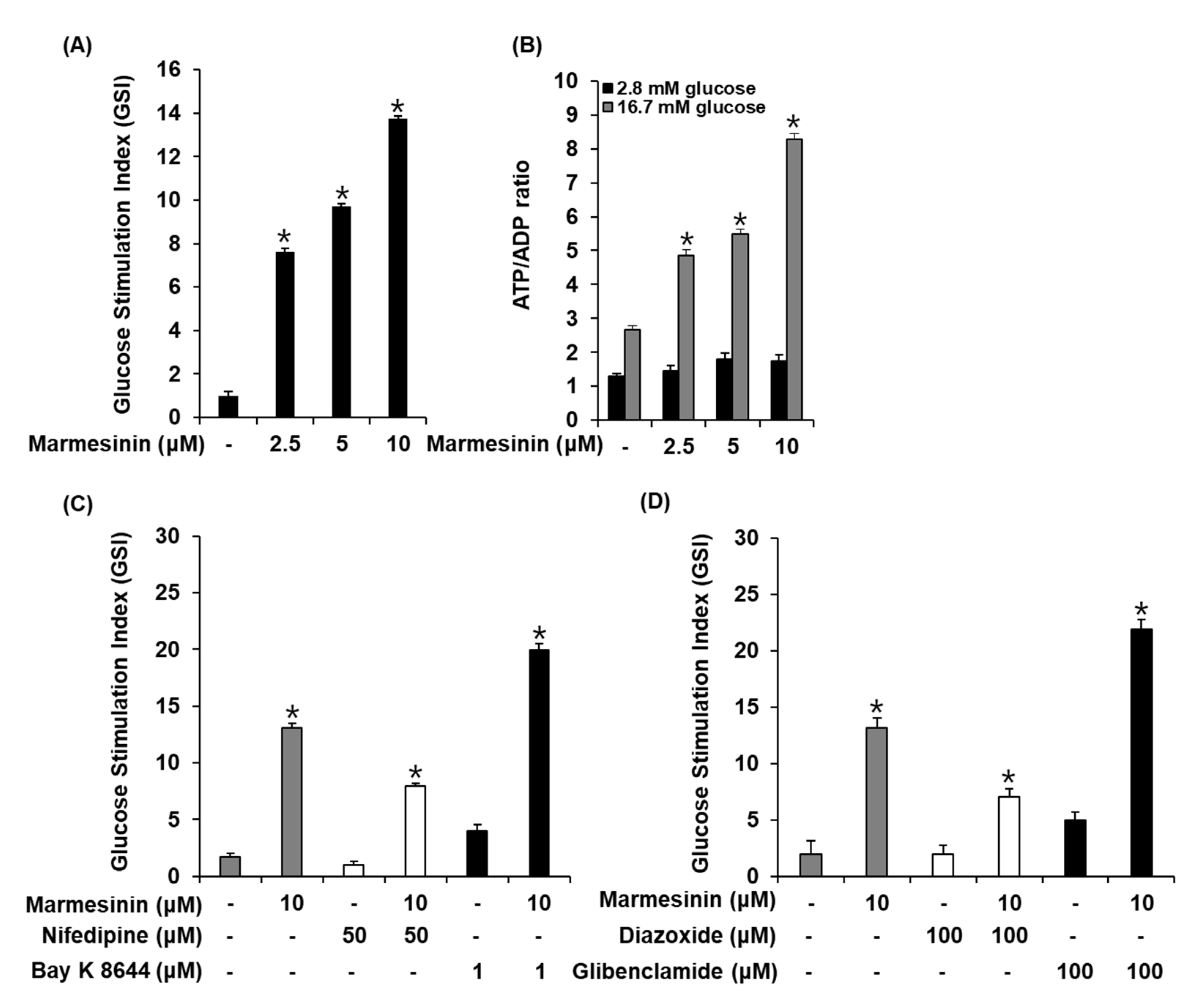
3.3. Effect of Marmesinin (19) on GSIS
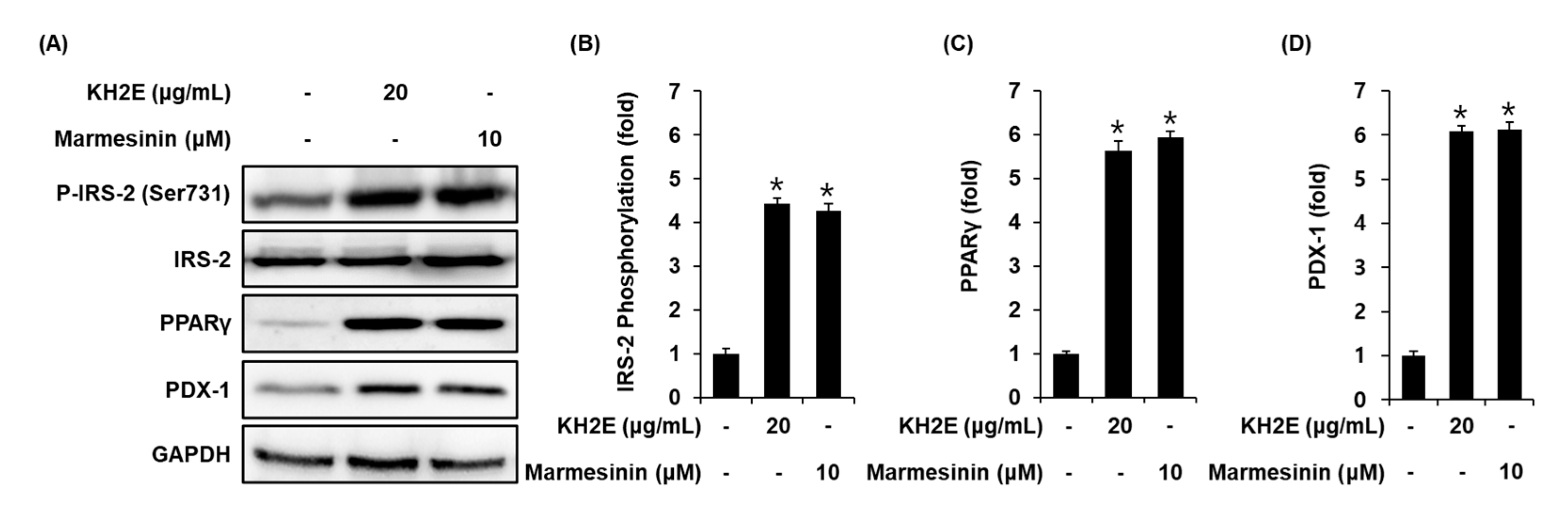
3.4. Effect of KH2E and Marmesinin (19) on the Protein Expression of P-IRS-2 (Ser731), IRS-2, PPARγ, and PDX-1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Saisho, Y. β-cell dysfunction: Its critical role in prevention and management of type 2 diabetes. World J. Diabetes 2015, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Pearson, E.R. Dorothy Hodgkin Lecture 2021: Drugs, genes and diabetes. Diabet. Med. 2021, 38, e14726. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Nowsherwan, N.; Abbass, M.; Ali, A.; Afridi, H.; Gul, W. Comparison of efficacy of combination of metformin plus modified-release gliclazide with combination of metformin plus sitagliptin in patients with type-2 diabetes mellitus. Am. J. Health Med. Nurs. Pract. 2021, 6, 38–44. [Google Scholar] [CrossRef]
- Vivó-Barrachina, L.; Rojas-Chacón, M.J.; Navarro-Salazar, R.; Belda-Sanchis, V.; Pérez-Murillo, J.; Peiró-Puig, A.; Herran-González, M.; Pérez-Bermejo, M. The Role of Natural Products on Diabetes Mellitus Treatment: A Systematic Review of Randomized Controlled Trials. Pharmaceutics 2022, 14, 101. [Google Scholar] [CrossRef]
- Hosseini Nasab, N.; Shah, F.H.; Kim, S.J. Pharmacological Role of Ostericum koreanum: A Short Viewpoint. Nat. Prod. Commun. 2021, 16, 1–8. [Google Scholar] [CrossRef]
- Lee, C.-K.; Woo, W.-S. Coumarin Constituents from the Roos of Angelica koreana Max. Korean J. Pharmacogn. 1982, 13, 10–13. [Google Scholar]
- Lee, B.Y.; Kwak, M.; Han, J.E.; Jung, E.-H.; Nam, G.-H. Ganghwal is a new species, Angelica reflexa. J. Species Res. 2013, 2, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Seo, Y.H.; Lee, A.Y.; Ryu, S.M.; Choi, G.; Moon, B.C.; Lee, J. Comparative Analysis of HPLC Component Patterns of Angelica reflexa roots by two cultivation types distributed in Korea. Korean Herb. Med. Inform. 2020, 8, 205–213. [Google Scholar]
- Kang, T.J.; Lee, S.Y.; Singh, R.P.; Agarwal, R.; Yim, D.S. Anti-tumor activity of oxypeucedanin from Ostericum koreanum against human prostate carcinoma DU145 cells. Acta Oncol. 2009, 48, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Shin, S. In vitro effects of essential oils from Ostericum koreanum against antibiotic-resistant Salmonella spp. Arch. Pharmacal Res. 2005, 28, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-J.; Bae, G.-S.; Kim, D.-Y.; Seo, S.-W.; Park, K.-B.; Kim, B.-J.; Song, J.-M.; Lee, K.-Y.; Na, C.; Shin, B.-C. Inhibitory effect of extract from Ostericum koreanum on LPS-induced proinflammatory cytokines production in RAW264. 7 cells. Korea J. Herbol. 2008, 23, 127–134. [Google Scholar]
- Park, Y.J.; Kim, H.J.; Lee, S.J.; Choi, H.-Y.; Jin, C.; Lee, Y.S. A new chromone, 11-hydroxy-sec-O-glucosylhamaudol from Ostericum koreanum. Chem. Pharm. Bull. 2007, 55, 1065–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, W.-S.; Kim, H.-H.; Ahn, D.-K.; Rhee, J.-S.; Ham, I.-H.; Choi, H.-Y. Effects of Angelicae koreanae radix on the vasomotor responses and focal cerebral ischemic damage by MCAO. Korea J. Herbol. 2004, 19, 147. [Google Scholar]
- Lee, S.-K.; Cho, H.-K.; Cho, S.-H.; Kim, S.-S.; Nahm, D.-H.; Park, H.-S. Occupational asthma and rhinitis caused by multiple herbal agents in a pharmacist. Ann. Allergy Asthma Immunol. 2001, 86, 469–474. [Google Scholar] [CrossRef]
- Jeon, J.-H.; Yang, J.-Y.; Chung, N.; Lee, H.-S. Contact and fumigant toxicities of 3-methylphenol isolated from Ostericum koreanum and its derivatives against house dust mites. J. Agric. Food Chem. 2012, 60, 12349–12354. [Google Scholar] [CrossRef]
- Song, J.-E.; Yang, J.-Y.; Lee, H.-S. Comparison with volatile compounds derived from essential oils of Ostericum spp. roots in Korea. J. Essent. Oil Bear. Plants 2015, 18, 1417–1420. [Google Scholar] [CrossRef]
- Lee, M.-K.; Ling, J.-H.; Chun, M.-H.; Jeong, J.-H.; Na, Y.-C.; Lee, K.-W.; Jung, J.H.; Hong, J. Simultaneous determination of biological marker compounds in Ostericum koreanum by HPLC method and discrimination by principal component analysis. Bull. Korean Chem. Soc. 2008, 29, 2465–2470. [Google Scholar]
- Jung, H.W.; Mahesh, R.; Park, J.H.; Boo, Y.C.; Park, K.M.; Park, Y.-K. Bisabolangelone isolated from Ostericum koreanum inhibits the production of inflammatory mediators by down-regulation of NF-κB and ERK MAP kinase activity in LPS-stimulated RAW264. 7 cells. Int. Immunopharmacol. 2010, 10, 155–162. [Google Scholar] [CrossRef]
- Kang, S.-W.; Kim, H.-K.; Lee, W.-J.; Ahn, Y.-J. Toxicity of bisabolangelone from Ostericum koreanum roots to Dermatophagoides farinae and Dermatophagoides pteronyssinus (Acari: Pyroglyphidae). J. Agric. Food Chem. 2006, 54, 3547–3550. [Google Scholar] [CrossRef]
- Mahesh, R.; Won Jung, H.; Hong Park, J.; Park, Y.-K. In vitro antioxidant capacity and neuronal cell toxicity of roots of Ostericum koreanum Maximowicz. E-J. Chem. 2011, 8, 1451–1455. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-Y.; Ahn, S.-J.; Baek, J.M.; Yoon, K.-H.; Lee, M.S.; Oh, J. Ostericum koreanum reduces LPS-induced bone loss through inhibition of Osteoclastogenesis. Am. J. Chin. Med. 2015, 43, 495–512. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ryu, S.M.; Nam, H.-h.; Kim, J.S.; Song, J.-h.; Seo, Y.H.; Kim, H.S.; Lee, A.Y.; Kim, W.J.; Lee, D.; Moon, B.C.; et al. Chemical Constituents of the Egg Cases of Tenodera angustipennis (Mantidis ootheca) with Intracellular Reactive Oxygen Species Scavenging Activity. Biomolecules 2021, 11, 556. [Google Scholar] [CrossRef]
- Kwon, M.-Y.; Park, E.; Lee, S.-J.; Chung, S.W. Heme oxygenase-1 accelerates erastin-induced ferroptotic cell death. Oncotarget 2015, 6, 24393. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.R.; Rhyu, D.Y. Anti-diabetic effects of Caulerpa lentillifera: Stimulation of insulin secretion in pancreatic β-cells and enhancement of glucose uptake in adipocytes. Asian Pac. J. Trop. Biomed. 2014, 4, 575–580. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Chen, P.-Y.; Wu, C.-C.; Tsai, I.-L.; Chen, I.-S. Chemical constituents and anti-platelet aggregation activity from the root of Peucedanum formosanum. J. Food Drug Anal. 2008, 16, 10. [Google Scholar] [CrossRef]
- Lee, H.K.; Oh, S.R.; Kwon, O.K.; Ahn, K.S.; Lee, J.; Kim, J.C.; Min, B.S.; Joung, H. Isolation of coumarins and ferulate from the roots of Angelica purpuraefolia and the antitumor activity of khellactone. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2007, 21, 406–409. [Google Scholar]
- Luz, R.F.; Vieira, I.J.; Braz-Filho, R.; Moreira, V.F. 13 C-NMR Data from Coumarins from Moraceae Family. Am. J. Anal. Chem. 2015, 6, 851. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.; Kim, H.P.; Kim, M.J.; Chun, W. Acetylcholinesterase inhibitors from Angelica polymorpha stem. Nat. Prod. Sci. 2017, 23, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Marumoto, S.; Miyazawa, M. Microbial transformation of isopimpinellin by Glomerella cingulata. J. Oleo Sci. 2011, 60, 575–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Ito, Y. Preparative isolation of imperatorin, oxypeucedanin and isoimperatorin from traditional Chinese herb “bai zhi” Angelica dahurica (Fisch. ex Hoffm) Benth. et Hook using multidimensional high-speed counter-current chromatography. J. Chromatogr. A 2006, 1115, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harkar, S.; Razdan, T.; Waight, E. Steroids, chromone and coumarins from Angelica officinalis. Phytochemistry 1984, 23, 419–426. [Google Scholar] [CrossRef]
- Shalaby, N.M.; Abd-Alla, H.I.; Aly, H.F.; Albalawy, M.A.; Shaker, K.H.; Bouajila, J. Preliminary in vitro and in vivo evaluation of antidiabetic activity of Ducrosia anethifolia Boiss. and its linear furanocoumarins. BioMed Res. Int. 2014, 2014, 480545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.Y.; Lee, S.; Jin, J.L.; Yun-Choi, H.S. Platelet anti-aggregatory effects of coumarins from the roots of Angelica genuflexa and A. gigas. Arch. Pharmacal Res. 2003, 26, 723–726. [Google Scholar] [CrossRef]
- Ganbaatar, J.; Shults, E.; Radnaeva, L.; Taraskin, V. Coumarins of Angelica deccurens. Mong. J. Chem. 2013, 14, 25–27. [Google Scholar] [CrossRef]
- Liu, R.; Wu, S.; Sun, A. Separation and purification of four chromones from radix saposhnikoviae by high-speed counter-current chromatography. Phytochem. Anal. Int. J. Plant Chem. Biochem. Tech. 2008, 19, 206–211. [Google Scholar] [CrossRef]
- Hong, G.U.; Lee, J.-Y.; Kang, H.; Kim, T.Y.; Park, J.Y.; Hong, E.Y.; Shin, Y.H.; Jung, S.H.; Chang, H.-B.; Kim, Y.H. Inhibition of osteoarthritis-related molecules by Isomucronulatol 7-O-β-d-glucoside and ecliptasaponin A in IL-1β-Stimulated chondrosarcoma cell model. Molecules 2018, 23, 2807. [Google Scholar] [CrossRef] [Green Version]
- Ito, C.; Fujiwara, K.; Kajita, M.; Ju-ichi, M.; Takemura, Y.; Suzuki, Y.; Tanaka, K.; Omura, M.; Furukawa, H. New coumarins from Citrus plants. Chem. Pharm. Bull. 1991, 39, 2509–2513. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Zhang, T.; Ito, Y. Preparative isolation of osthol and xanthotoxol from Common Cnidium Fruit (Chinese traditional herb) using stepwise elution by high-speed counter-current chromatography. J. Chromatogr. A 2004, 1033, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-H.; Chen, Y.-C.; Hwang, T.-L.; Shu, C.-W.; Sung, P.-J.; Lim, Y.-P.; Kuo, W.-L.; Chen, J.-J. New coumarins and anti-inflammatory constituents from the fruits of Cnidium monnieri. Int. J. Mol. Sci. 2014, 15, 9566–9578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.W.; Cheng, M.J.; Peng, C.F.; Chen, I.S. Secondary metabolites and antimycobacterial activities from the roots of Ficus nervosa. Chem. Biodivers. 2010, 7, 1814–1821. [Google Scholar] [CrossRef] [PubMed]
- de Melo Cazal, C.; de Cássia Domingues, V.; Batalhão, J.R.; Bueno, O.C.; Rodrigues Filho, E.; da Silva, M.F.G.F.; Vieira, P.C.; Fernandes, J.B. Isolation of xanthyletin, an inhibitor of ants’ symbiotic fungus, by high-speed counter-current chromatography. J. Chromatogr. A 2009, 1216, 4307–4312. [Google Scholar] [CrossRef] [PubMed]
- Skalicka-Woźniak, K.; Mroczek, T.; Garrard, I.; Głowniak, K. Isolation of the new minor constituents dihydropyranochromone and furanocoumarin from fruits of Peucedanum alsaticum L. by high-speed counter-current chromatography. J. Chromatogr. A 2009, 1216, 5669–5675. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Taguchi, H.; Endo, T.; YOSIOKA, I. The constituents of Ledebouriella seseloides Wolff. I. Structures of three new chromones. Chem. Pharm. Bull. 1982, 30, 3555–3562. [Google Scholar] [CrossRef] [Green Version]
- Karakaya, S.; Gözcü, S.; Güvenalp, Z.; Özbek, H.; Yuca, H.; Dursunoğlu, B.; Kazaz, C.; Kılıç, C.S. The α-amylase and α-glucosidase inhibitory activities of the dichloromethane extracts and constituents of Ferulago bracteata roots. Pharm. Biol. 2018, 56, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, S.; Muro, S.; Kobayashi, M.; Miyazawa, M.; Hirai, Y. Absolute structures of some naturally occurring isopropenyldihydrobenzofurans, remirol, remiridiol, angenomalin, and isoangenomalin. J. Org. Chem. 2003, 68, 6274–6278. [Google Scholar] [CrossRef] [PubMed]
- VanWagenen, B.C.; Huddleston, J.; Cardellina, J.H. Native American food and medicinal plants, 8. Water-soluble constituents of Lomatium dissectum. J. Nat. Prod. 1988, 51, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Prachayasittikul, S.; Suphapong, S.; Worachartcheewan, A.; Lawung, R.; Ruchirawat, S.; Prachayasittikul, V. Bioactive metabolites from Spilanthes acmella Murr. Molecules 2009, 14, 850–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sur, T.K.; Hazra, A.K.; Bhattacharyya, D.; Hazra, A. Antiradical and antidiabetic properties of standardized extract of Sunderban mangrove, Rhizophora mucronata. Pharmacogn. Mag. 2015, 11, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salahuddin, M.; Jalalpure, S.S.; Gadge, N.B. Antidiabetic activity of aqueous bark extract of Cassia glauca in streptozotocin-induced diabetic rats. Can. J. Physiol. Pharmacol. 2010, 88, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Ji, Y.; Li, W.; Liu, Y.; Fu, R.; Xiang, M. Protective effects of the total coumarin fraction of Urtica dentata on experimental diabetic nephropathy in vitro and in vivo. Planta Med. 2015, 81, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yao, Y.; Li, L. Coumarins as potential antidiabetic agents. J. Pharm. Pharmacol. 2017, 69, 1253–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousof Ali, M.; Jung, H.A.; Choi, J.S. Anti-diabetic and anti-Alzheimer’s disease activities of Angelica decursiva. Arch. Pharmacal Res. 2015, 38, 2216–2227. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Liao, C.; Chen, J.; Zhou, M. In vitro and in vivo growth inhibition of human leukemia cells by Nodakenetin are mediated via mitochondrial apoptosis, cell cycle arrest and inhibition of cell migration and invasion. J. BUON 2019, 24, 1219–1224. [Google Scholar]
- Bharti, S.K.; Krishnan, S.; Kumar, A.; Kumar, A. Antidiabetic phytoconstituents and their mode of action on metabolic pathways. Ther. Adv. Endocrinol. Metab. 2018, 9, 81–100. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, M.S.; Lee, B.H.; Kim, J.-K.; Ahn, E.-K.; Ko, H.-J.; Cho, Y.-R.; Lee, S.-J.; Bae, G.-U.; Kim, Y.K. Marmesin-mediated suppression of VEGF/VEGFR and integrin β1 expression: Its implication in non-small cell lung cancer cell responses and tumor angiogenesis. Oncol. Rep. 2017, 37, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhong, S.; Wu, H.; Wu, Q. In vitro anti-cancer effect of marmesin by suppression of PI3K/Akt pathway in esophagus cancer cells. Esophagus 2022, 19, 163–174. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, J.-K.; Ahn, E.-K.; Ko, H.-J.; Cho, Y.-R.; Lee, C.H.; Kim, Y.K.; Bae, G.-U.; Oh, J.S.; Seo, D.-W. Marmesin is a novel angiogenesis inhibitor: Regulatory effect and molecular mechanism on endothelial cell fate and angiogenesis. Cancer Lett. 2015, 369, 323–330. [Google Scholar] [CrossRef]
- Thaifa, M.; Roshna, S.; Arya, U.; Babu, A.G. A review on diabetes mellitus and diabetic neuropathy: A plant based approach. J. Pharmacogn. Phytochem. 2017, 6, 506–510. [Google Scholar]
- Kang, S.Y.; Kim, Y.C. Neuroprotective coumarins from the root of Angelica gigas: Structure-activity relationships. Arch. Pharmacal Res. 2007, 30, 1368–1373. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.-I.; Lee, J.-H.; Lee, Y.-C.; Kim, K.-S. Antiplasmodial and cytotoxic activity of coumarin derivatives from dried roots of Angelica gigas Nakai in vitro. Immunopharmacol. Immunotoxicol. 2011, 33, 663–666. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Deng, S.; Nie, H.; Huang, Y.; Liu, B.; Yang, R.; Huang, S.; Zhou, D.; Chen, H.; Li, J. Two new coumarins from the bark of Streblus indicus (Bur.) Corner. Nat. Prod. Res. 2017, 31, 1052–1058. [Google Scholar] [CrossRef]
- Oloya, B.; Namukobe, J.; Heydenreich, M.; Ssengooba, W.; Schmidt, B.; Byamukama, R. Antimycobacterial activity of the extract and isolated compounds from the stem bark of Zanthoxylum leprieurii Guill. and Perr. Nat. Prod. Commun. 2021, 16, 1–8. [Google Scholar] [CrossRef]
- Vimal, V.; Devaki, T. Linear furanocoumarin protects rat myocardium against lipidperoxidation and membrane damage during experimental myocardial injury. Biomed. Pharmacother. 2004, 58, 393–400. [Google Scholar] [CrossRef]
- Alizadeh, M.; Jalal, M.; Hamed, K.; Saber, A.; Kheirouri, S.; Pourteymour Fard Tabrizi, F.; Kamari, N. Recent updates on anti-inflammatory and antimicrobial effects of furan natural derivatives. J. Inflamm. Res. 2020, 13, 451–463. [Google Scholar] [CrossRef]
- He, J.; Zhu, N.-L.; Kong, J.; Peng, P.; Li, L.-F.; Wei, X.-L.; Jiang, Y.-Y.; Zhang, Y.-L.; Bian, B.-L.; She, G.-M. A newly discovered phenylethanoid glycoside from Stevia rebaudiana Bertoni affects insulin secretion in rat INS-1 islet β cells. Molecules 2019, 24, 4178. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Rebsdorf, A.; Anker, C.; Gregersen, S.; Hermansen, K.; Geuns, J.M.; Jeppesen, P.B. Steviol glucuronide, a metabolite of steviol glycosides, potently stimulates insulin secretion from isolated mouse islets: Studies in vitro. Endocrinol. Diabetes Metab. 2019, 2, e00093. [Google Scholar] [CrossRef]
- Adki, K.M.; Kulkarni, Y.A. Glycosides from natural sources in the treatment of diabetes mellitus. Struct. Health Eff. Nat. Prod. Diabetes Mellit. 2021, 1, 81–102. [Google Scholar]
- Ashcroft, F.M. ATP-sensitive potassium channelopathies: Focus on insulin secretion. J. Clin. Investig. 2005, 115, 2047–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seino, S. Cell signalling in insulin secretion: The molecular targets of ATP, cAMP and sulfonylurea. Diabetologia 2012, 55, 2096–2108. [Google Scholar] [CrossRef] [PubMed]
- Kusunoki, M.; Hayashi, M.; Shoji, T.; Uba, T.; Tanaka, H.; Sumi, C.; Matsuo, Y.; Hirota, K. Propofol inhibits stromatoxin-1-sensitive voltage-dependent k+ channels in pancreatic β-cells and enhances insulin secretion. PeerJ 2019, 7, e8157. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, L.; Li, Y.; Chen, H.; Sun, Z. Nifedipine protects INS-1 β-cell from high glucose-induced ER stress and apoptosis. Int. J. Mol. Sci. 2011, 12, 7569–7580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardy, G.; Virsolvy, A.; Quignard, J.; Ravier, M.; Bertrand, G.; Dalle, S.; Cros, G.; Magous, R.; Richard, S.; Oiry, C. Quercetin induces insulin secretion by direct activation of L-type calcium channels in pancreatic beta cells. Br. J. Pharmacol. 2013, 169, 1102–1113. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Hwang, Y.-C.; Koo, S.-H.; Park, K.S.; Lee, M.-S.; Kim, K.-W.; Lee, M.-K. PPAR-γ activation increases insulin secretion through the up-regulation of the free fatty acid receptor GPR40 in pancreatic β-cells. PLoS ONE 2013, 8, e50128. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-i.; Ahn, Y.-h. Role of peroxisome proliferator-activated receptor-γ in the glucose-sensing apparatus of liver and β-cells. Diabetes 2004, 53, S60–S65. [Google Scholar] [CrossRef] [Green Version]
- Kushner, J.A.; Ye, J.; Schubert, M.; Burks, D.J.; Dow, M.A.; Flint, C.L.; Dutta, S.; Wright, C.V.; Montminy, M.R.; White, M.F. Pdx1 restores β cell function in Irs2 knockout mice. J. Clin. Investig. 2002, 109, 1193–1201. [Google Scholar] [CrossRef]
- Suzuki, R.; Tobe, K.; Terauchi, Y.; Komeda, K.; Kubota, N.; Eto, K.; Yamauchi, T.; Azuma, K.; Kaneto, H.; Taguchi, T. Pdx1 expression in Irs2-deficient mouse β-cells is regulated in a strain-dependent manner. J. Biol. Chem. 2003, 278, 43691–43698. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 a,c | 2 a,d | 3 b,c | ||||
|---|---|---|---|---|---|---|
| Position | δC, Type | δH Multi (J in Hz) | δC, Type | δH Multi (J in Hz) | δC, Type | δH Multi (J in Hz) |
| 2 | 161.1, C | 161.3, C | 163.4, C | |||
| 3 | 112.9, CH2 | 6.24, d (9.9) | 112.8, CH2 | 6.30, d (9.7) | 112.5, CH | 6.19, d (9.5) |
| 4 | 139.3, CH | 8.12, d (9.9), | 139.6, CH | 8.24, d (9.7), | 146.5, CH | 7.86, d (9.5) |
| 5 | 148.6, C | 148.9, C | 130.5, CH | 7.41, d (8.3) | ||
| 6 | 113.3, C | 114.1, C | 108.1, CH | 6.78, d (8.3) | ||
| 7 | 158.1, C | 158.1, C | 165.8, C | |||
| 8 | 94.4, CH | 7.14, s | 94.5, CH | 7.48, s | 115.4, C | |
| 9 | 152.6, C | 152.6, C | 152.7, C | |||
| 10 | 107.0, C | 107.4, C | 114.6, C | |||
| 2′ | 145.1, CH | 7.61, d (2.2) | 145.0, CH | 7.61, d (2.0) | 74.6, C | |
| 3′ | 104.9, CH | 7.01, d (2.2) | 104.9, CH | 6.95, d (2.0) | 88.7, CH | 5.02, dd (9.9, 8.0) |
| 4′ | - | - | - | - | 68.2, CH2 | 3.32, dd (16.1, 9.9), 3.43, dd (16.1, 8.0) |
| 5′ | - | - | - | - | 27.6, CH2 | 3.53, d (10.5), 3.72, d (10.5) |
| 6′ | - | - | - | - | 19.7, CH3 | 1.21, s |
| 1″ | 71.7, CH2 | 4.69, dd (10.2, 8.0), 4.85, dd (10.2, 2.6) | 74.3, CH2 | 4.31, dd (9.9, 8.0), 4.48 dd (9.9, 3.0) | - | - |
| 2″ | 77.4, CH | 5.44, dd (8.0, 2.6) | 77.4, CH | 3.93, dd (8.0, 3.0) | - | - |
| 3″ | 71.7, C | 71.7, C | - | - | ||
| 4″ | 26.5, CH3 | 1.38, s | 26.5, CH3 | 1.21, s | - | - |
| 5″ | 26.6, CH3 | 1.41, s | 26.6, CH3 | 1.19, s | - | - |
| 1′′′ | 126.5, C | 131.8, C | - | - | ||
| 2′′′ | 109.4, CH2 | 7.02, d (1.1) | 128.3, CH | 7.19, d (8.3) | - | - |
| 3′′′ | 146.8, C | 115.3, CH | 6.80, d (8.2) | - | - | |
| 4′′′ | 148.4, C | 155.4, C | - | - | ||
| 5′′′ | 114.8, CH | 6.94, d (8.2) | 115.3, CH | 6.80, d (8.2) | - | - |
| 6′′′ | 123.3, CH | 7.08, dd (8.2, 1.3) | 128.3, CH | 7.19, d (8.3) | - | - |
| 7′′′ | 146.4, CH | 7.67, d (15.9) | 80.8, CH | 4.34, t (5.7) | - | - |
| 8′′′ | 114.2, CH | 6.31, d (15.9) | 66.3, CH2 | 3.52, dd (9.7, 4.9), 3.67 dd (9.7, 6.4) | - | - |
| 9′′′ | 166.7, C | 64.4, CH2 | 3.38, qd (9.2, 7.0), 3.45 qd (9.2, 7.0) | - | - | |
| 10′′′ | - | - | 15.2, CH3 | 1.18, t (7.0) | - | - |
| 3′′′-OMe | 56.0, CH3 | 3.95, s | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-S.; Lee, D.; Seo, Y.-H.; Ryu, S.-M.; Lee, A.-Y.; Moon, B.-C.; Kim, W.-J.; Kang, K.-S.; Lee, J. Chemical Constituents from the Roots of Angelica reflexa That Improve Glucose-Stimulated Insulin Secretion by Regulating Pancreatic β-Cell Metabolism. Pharmaceutics 2023, 15, 1239. https://doi.org/10.3390/pharmaceutics15041239
Kim H-S, Lee D, Seo Y-H, Ryu S-M, Lee A-Y, Moon B-C, Kim W-J, Kang K-S, Lee J. Chemical Constituents from the Roots of Angelica reflexa That Improve Glucose-Stimulated Insulin Secretion by Regulating Pancreatic β-Cell Metabolism. Pharmaceutics. 2023; 15(4):1239. https://doi.org/10.3390/pharmaceutics15041239
Chicago/Turabian StyleKim, Hyo-Seon, Dahae Lee, Young-Hye Seo, Seung-Mok Ryu, A-Yeong Lee, Byeong-Cheol Moon, Wook-Jin Kim, Ki-Sung Kang, and Jun Lee. 2023. "Chemical Constituents from the Roots of Angelica reflexa That Improve Glucose-Stimulated Insulin Secretion by Regulating Pancreatic β-Cell Metabolism" Pharmaceutics 15, no. 4: 1239. https://doi.org/10.3390/pharmaceutics15041239
APA StyleKim, H. -S., Lee, D., Seo, Y. -H., Ryu, S. -M., Lee, A. -Y., Moon, B. -C., Kim, W. -J., Kang, K. -S., & Lee, J. (2023). Chemical Constituents from the Roots of Angelica reflexa That Improve Glucose-Stimulated Insulin Secretion by Regulating Pancreatic β-Cell Metabolism. Pharmaceutics, 15(4), 1239. https://doi.org/10.3390/pharmaceutics15041239