

Activation of Insulin Gene Expression via Transfection of a CRISPR/dCas9a System Using Magnetic Peptide-Imprinted Nanoparticles

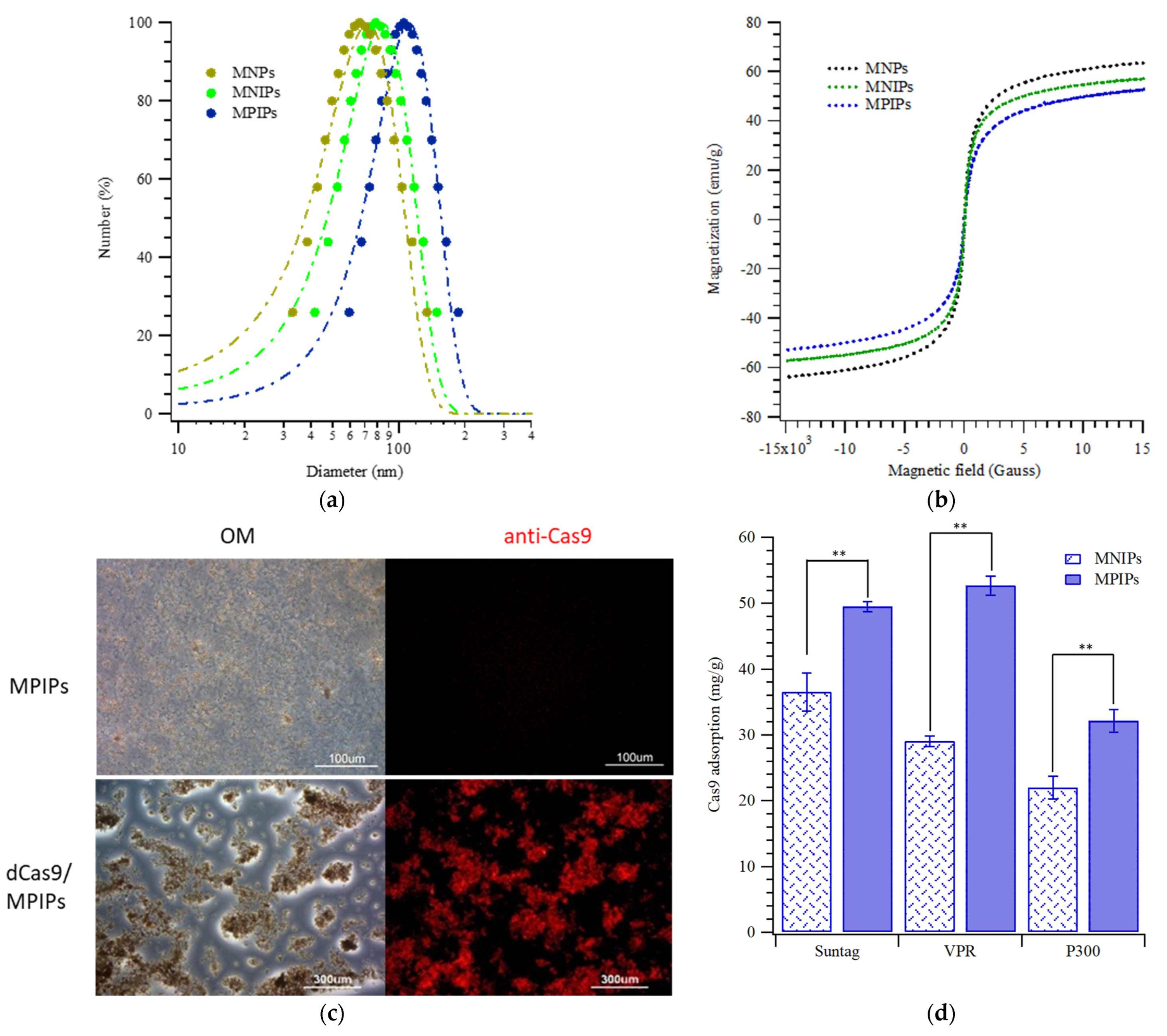{kind=link}
{kind=link}
{kind=link}
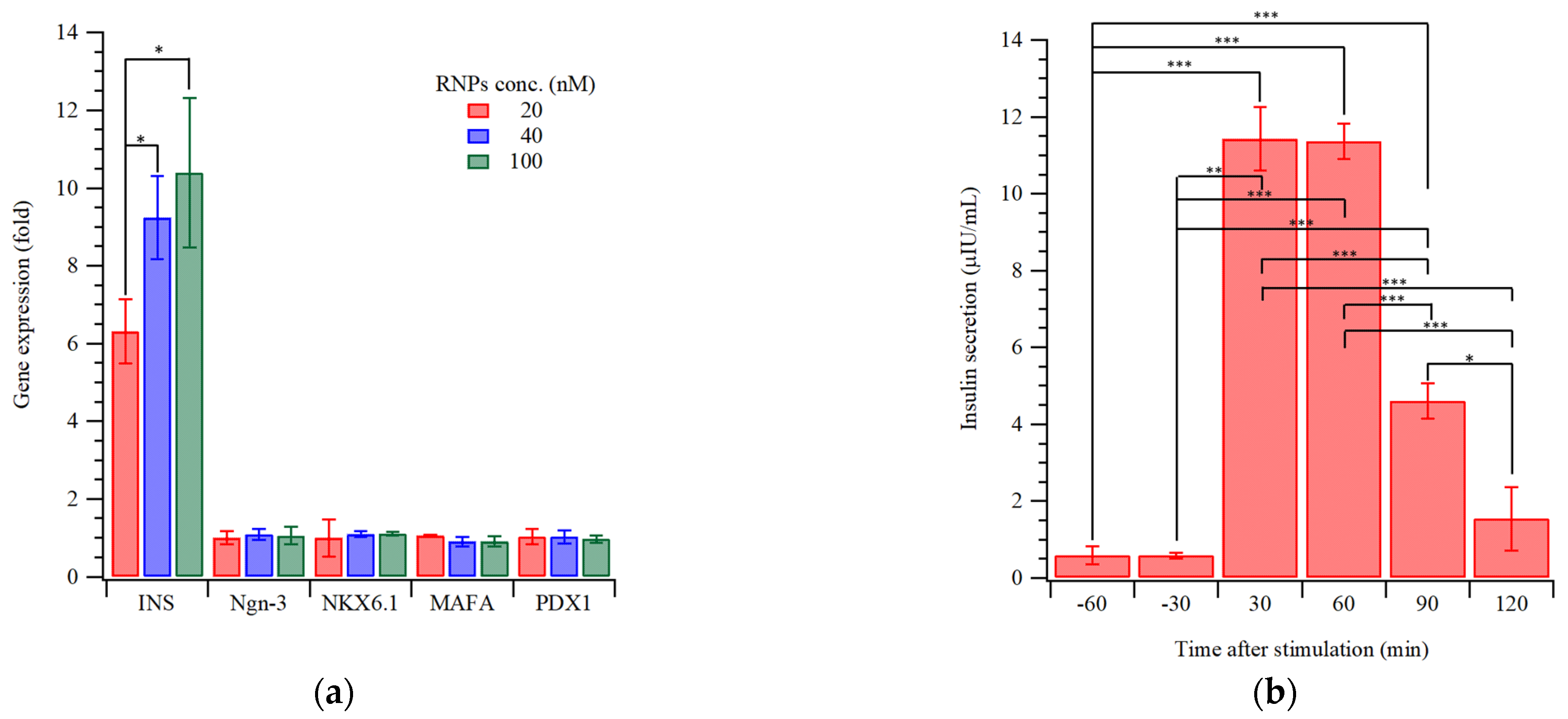{kind=link}
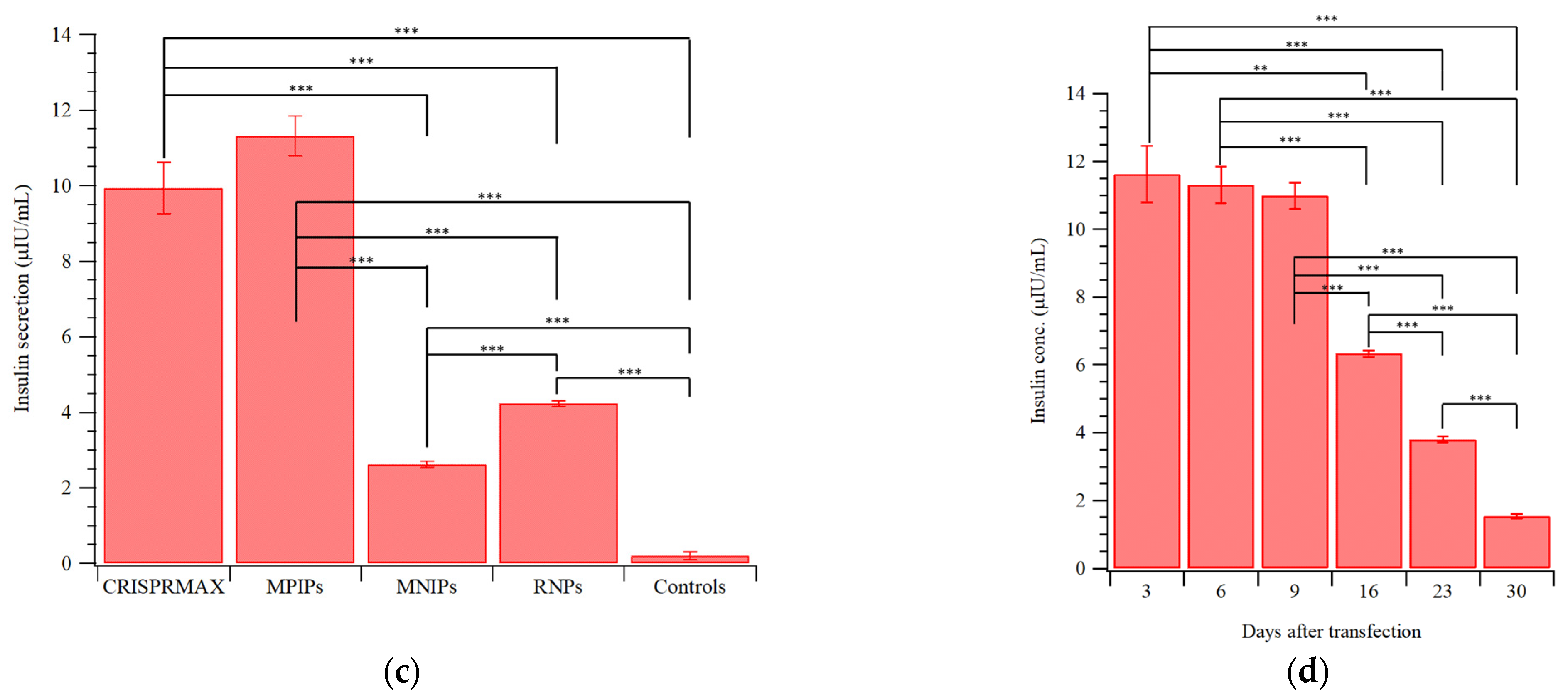{kind=link}
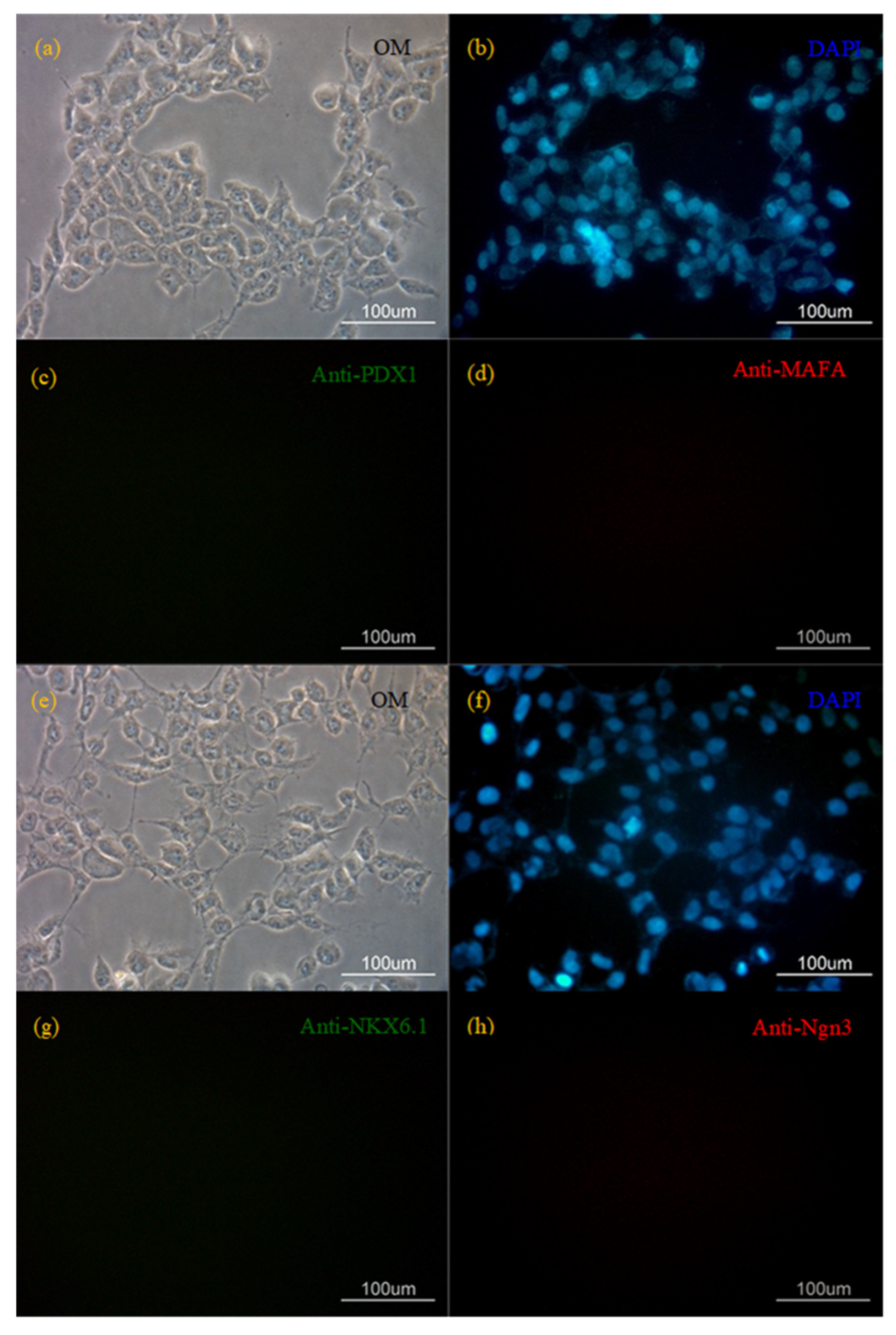{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experiment
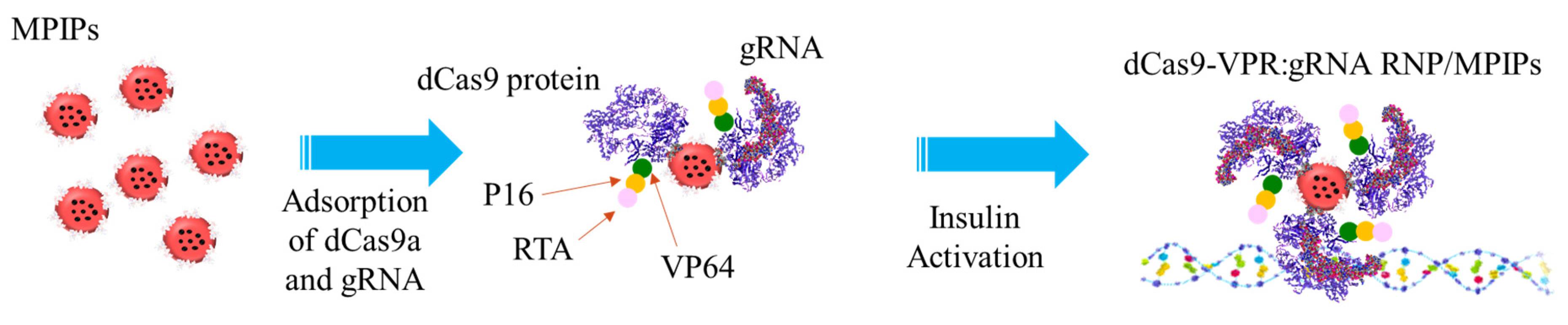
2.1. Delivery of RNP with MPIPs to HEK 293T Cells
2.2. Measurement of Insulin Release with Glucose Stimulation
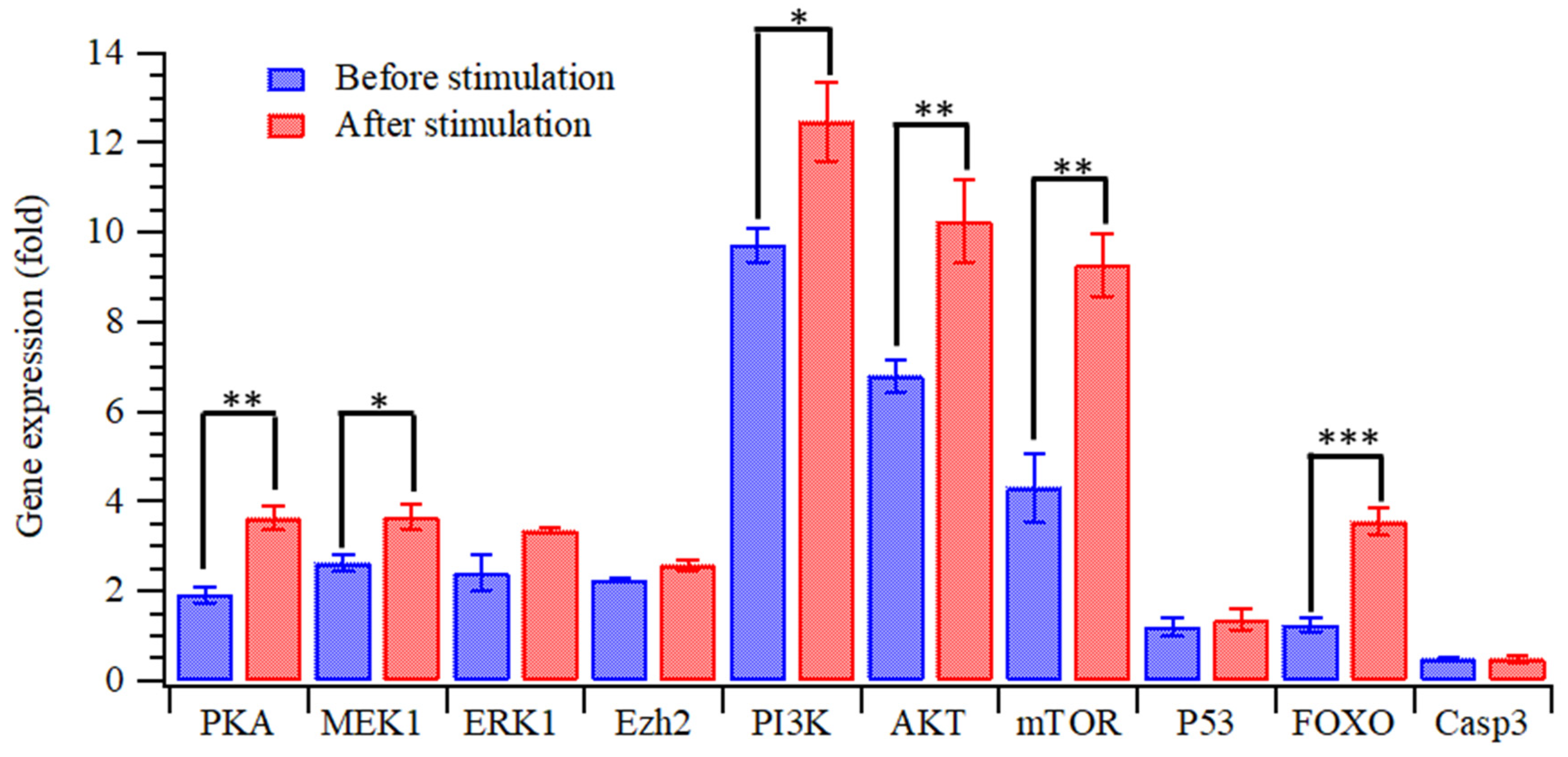
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fu, Z.; Gilbert, E.R.; Liu, D. Regulation of insulin synthesis and secretion and pancreatic Beta-cell dysfunction in diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar] [PubMed]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36 (Suppl. 1), S67–S74. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, M. Insulin resistance and pancreatic beta cell failure. J. Clin. Investig. 2006, 116, 1756–1760. [Google Scholar] [CrossRef] [PubMed]
- Cerf, M.E. Beta cell dysfunction and insulin resistance. Front. Endocrinol. 2013, 4, 37. [Google Scholar] [CrossRef]
- Roep, B.O.; Thomaidou, S.; van Tienhoven, R.; Zaldumbide, A. Type 1 diabetes mellitus as a disease of the β-cell (do not blame the immune system?). Nat. Rev. Endocrinol. 2021, 17, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, F.M.; Rorsman, P. Diabetes Mellitus and the β Cell: The Last Ten Years. Cell 2012, 148, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Alejandro, E.U.; Gregg, B.; Blandino-Rosano, M.; Cras-Méneur, C.; Bernal-Mizrachi, E. Natural history of β-cell adaptation and failure in type 2 diabetes. Mol. Asp. Med. 2015, 42, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Hudish, L.I.; Reusch, J.E.; Sussel, L. β Cell dysfunction during progression of metabolic syndrome to type 2 diabetes. J. Clin. Investig. 2019, 129, 4001–4008. [Google Scholar] [CrossRef]
- Shah, R.B.; Patel, M.; Maahs, D.M.; Shah, V.N. Insulin delivery methods: Past, present and future. Int. J. Pharm. Investig. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Xiong, X.; Chen, M.; Lim, W.A.; Zhao, D.; Qi, L.S. CRISPR/Cas9 for human genome engineering and disease research. Annu. Rev. Genom. Hum. Genet. 2016, 17, 131–154. [Google Scholar] [CrossRef] [PubMed]
- Chavez, A.; Tuttle, M.; Pruitt, B.W.; Ewen-Campen, B.; Chari, R.; Ter-Ovanesyan, D.; Haque, S.J.; Cecchi, R.J.; Kowal, E.J.K.; Buchthal, J.; et al. Comparison of Cas9 activators in multiple species. Nat. Methods 2016, 13, 563. [Google Scholar] [CrossRef] [PubMed]
- Hasanzadeh, A.; Noori, H.; Jahandideh, A.; Haeri Moghaddam, N.; Kamrani Mousavi, S.M.; Nourizadeh, H.; Saeedi, S.; Karimi, M.; Hamblin, M.R. Smart Strategies for Precise Delivery of CRISPR/Cas9 in Genome Editing. ACS Appl. Bio Mater. 2022, 5, 413–437. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, X.; Wu, D.; Xin, H.; Chen, D.; Li, D.; Pan, H.; Zhou, C.; Ping, Y. Delivery of CRISPR/Cas9 Plasmids by Cationic Gold Nanorods: Impact of the Aspect Ratio on Genome Editing and Treatment of Hepatic Fibrosis. Chem. Mater. 2021, 33, 81–91. [Google Scholar] [CrossRef]
- Huang, X.; Zheng, R.; Ding, F.; Yang, J.; Xie, M.; Liu, X.; Li, J.; Feng, J.; Zhu, X.; Zhang, C. Efficient Delivery of mRNA Using Crosslinked Nucleic Acid Nanogel as a Carrier. ACS Mater. Lett. 2020, 2, 1509–1515. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, L.; Liu, H.; Cheng, K. Delivery strategies of the CRISPR-Cas9 gene-editing system for therapeutic applications. J. Control. Release 2017, 266, 17–26. [Google Scholar] [CrossRef]
- Yin, H.; Sun, L.; Pu, Y.; Yu, J.; Feng, W.; Dong, C.; Zhou, B.; Du, D.; Zhang, Y.; Chen, Y.; et al. Ultrasound-Controlled CRISPR/Cas9 System Augments Sonodynamic Therapy of Hepatocellular Carcinoma. ACS Cent. Sci. 2021, 7, 2049–2062. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, X.; Liu, J.; Chen, Q.; Zhao, Y. Convergent Arrangement of sgRNA and Cas9 in CRISPRsome for Transcellular Trafficking. ACS Mater. Lett. 2022, 4, 505–510. [Google Scholar] [CrossRef]
- Perez-Pinera, P.; Kocak, D.D.; Vockley, C.M.; Adler, A.F.; Kabadi, A.M.; Polstein, L.R.; Thakore, P.I.; Glass, K.A.; Ousterout, D.G.; Leong, K.W. RNA-guided gene activation by CRISPR-Cas9–based transcription factors. Nat. Methods 2013, 10, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Thomas, J.L.; Ho, M.-H.; Yuan, C.; Lin, H.-Y. Synthesis of magnetic molecularly imprinted poly (ethylene-co-vinyl alcohol) nanoparticles and their uses in the extraction and sensing of target molecules in urine. ACS Appl. Mater. Interfaces 2010, 2, 1729–1736. [Google Scholar] [CrossRef]
- Ding, S.; Lyu, Z.; Niu, X.; Zhou, Y.; Liu, D.; Falahati, M.; Du, D.; Lin, Y. Integrating ionic liquids with molecular imprinting technology for biorecognition and biosensing: A review. Biosens. Bioelectron. 2020, 149, 111830. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Thomas, J.L.; Chen, Y.-C.; Wang, H.-Y.; Lin, H.-Y. Hydrolysis of Magnetic Amylase-Imprinted Poly(ethylene-co-vinyl alcohol) Composite Nanoparticles. ACS Appl. Mater. Interfaces 2012, 4, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Liu, K.-H.; Thomas, J.L.; Chen, J.-R.; Lin, H.-Y. Immunotherapy of Hepatocellular Carcinoma with Magnetic PD-1 Peptide-Imprinted Polymer Nanocomposite and Natural Killer Cells. Biomolecules 2019, 9, 651. [Google Scholar] [CrossRef]
- Lee, M.-H.; Lin, C.-C.; Thomas, J.L.; Li, J.-A.; Lin, H.-Y. Cellular reprogramming with multigene activation by the delivery of CRISPR/dCas9 ribonucleoproteins via magnetic peptide-imprinted chitosan nanoparticles. Mater. Today Bio 2021, 9, 100091. [Google Scholar] [CrossRef]
- Liu, K.-H.; Lin, H.-Y.; Thomas, J.L.; Shih, Y.-P.; Yang, Z.-Y.; Chen, J.-T.; Lee, M.-H. Synthesis of ginsenoside Rb1-imprinted magnetic polymer nanoparticles for the extraction and cellular delivery of therapeutic ginsenosides. J. Ginseng Res. 2022, 46, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Masoum, S. Molecularly imprinted polymers for capturing and sensing proteins: Current progress and future implications. TrAC Trends Anal. Chem. 2019, 114, 29–47. [Google Scholar] [CrossRef]
- Wilbie, D.; Walther, J.; Mastrobattista, E. Delivery aspects of CRISPR/Cas for in vivo genome editing. Acc. Chem. Res. 2019, 52, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Lino, C.A.; Harper, J.C.; Carney, J.P.; Timlin, J.A. Delivering CRISPR: A review of the challenges and approaches. Drug Deliv. 2018, 25, 1234–1257. [Google Scholar] [CrossRef]
- Ramamoorth, M.; Narvekar, A. Non viral vectors in gene therapy-an overview. J. Clin. Diagn. Res. JCDR 2015, 9, GE01. [Google Scholar] [CrossRef]
- Liu, B.-Y.; He, X.-Y.; Xu, C.; Xu, L.; Ai, S.-L.; Cheng, S.-X.; Zhuo, R.-X. A Dual-Targeting Delivery System for Effective Genome Editing and In Situ Detecting Related Protein Expression in Edited Cells. Biomacromolecules 2018, 19, 2957–2968. [Google Scholar] [CrossRef]
- Zhang, H.; Bahamondez-Canas, T.F.; Zhang, Y.; Leal, J.; Smyth, H.D.C. PEGylated Chitosan for Nonviral Aerosol and Mucosal Delivery of the CRISPR/Cas9 System in Vitro. Mol. Pharm. 2018, 15, 4814–4826. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Lin, C.-C.; Thomas, J.L.; Chan, C.-K.; Lin, H.-Y. Epitope recognition of magnetic peptide-imprinted chitosan composite nanoparticles for the extraction of CRISPR/dCas9a proteins from transfected cells. Nanotechnology 2021, 32, 18LT02. [Google Scholar] [CrossRef] [PubMed]
- Giménez, C.A.; Ielpi, M.; Mutto, A.; Grosembacher, L.; Argibay, P.; Pereyra-Bonnet, F. CRISPR-on system for the activation of the endogenous human INS gene. Gene Ther. 2016, 23, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Thomas, J.L.; Lin, C.-Y.; Li, Y.-C.E.; Lin, H.-Y. Nanoparticle-mediated CRISPR/dCas9a activation of multiple transcription factors to engineer insulin-producing cells. J. Mater. Chem. B 2023, 11, 1866–1870. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Brown, J.; Kanarek, A.; Rajagopal, J.; Melton, D.A. In vivo reprogramming of adult pancreatic exocrine cells to β-cells. Nature 2008, 455, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Phua, K.K.; Leong, K.W.; Nair, S.K. Transfection efficiency and transgene expression kinetics of mRNA delivered in naked and nanoparticle format. J. Control. Release 2013, 166, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Blandino-Rosano, M.; Chen, A.Y.; Scheys, J.O.; Alejandro, E.U.; Gould, A.P.; Taranukha, T.; Elghazi, L.; Cras-Méneur, C.; Bernal-Mizrachi, E. mTORC1 signaling and regulation of pancreatic β-cell mass. Cell Cycle 2012, 11, 1892–1902. [Google Scholar] [CrossRef]
- Müller, T.D.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.J.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef]
- Glauser, D.A.; Schlegel, W. The emerging role of FOXO transcription factors in pancreatic β cells. J. Endocrinol. 2007, 193, 195–207. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-H.; Thomas, J.L.; Lin, C.-Y.; Li, Y.-C.E.; Lin, H.-Y. Activation of Insulin Gene Expression via Transfection of a CRISPR/dCas9a System Using Magnetic Peptide-Imprinted Nanoparticles. Pharmaceutics 2023, 15, 1311. https://doi.org/10.3390/pharmaceutics15041311
Lee M-H, Thomas JL, Lin C-Y, Li Y-CE, Lin H-Y. Activation of Insulin Gene Expression via Transfection of a CRISPR/dCas9a System Using Magnetic Peptide-Imprinted Nanoparticles. Pharmaceutics. 2023; 15(4):1311. https://doi.org/10.3390/pharmaceutics15041311
Chicago/Turabian StyleLee, Mei-Hwa, James L. Thomas, Chien-Yu Lin, Yi-Chen Ethan Li, and Hung-Yin Lin. 2023. "Activation of Insulin Gene Expression via Transfection of a CRISPR/dCas9a System Using Magnetic Peptide-Imprinted Nanoparticles" Pharmaceutics 15, no. 4: 1311. https://doi.org/10.3390/pharmaceutics15041311
APA StyleLee, M. -H., Thomas, J. L., Lin, C. -Y., Li, Y. -C. E., & Lin, H. -Y. (2023). Activation of Insulin Gene Expression via Transfection of a CRISPR/dCas9a System Using Magnetic Peptide-Imprinted Nanoparticles. Pharmaceutics, 15(4), 1311. https://doi.org/10.3390/pharmaceutics15041311