
Pre-Clinical Assessment of Roflumilast Therapy in a Thoracic Model of Spinal Cord Injury

 , , , , ,
, , , , , 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
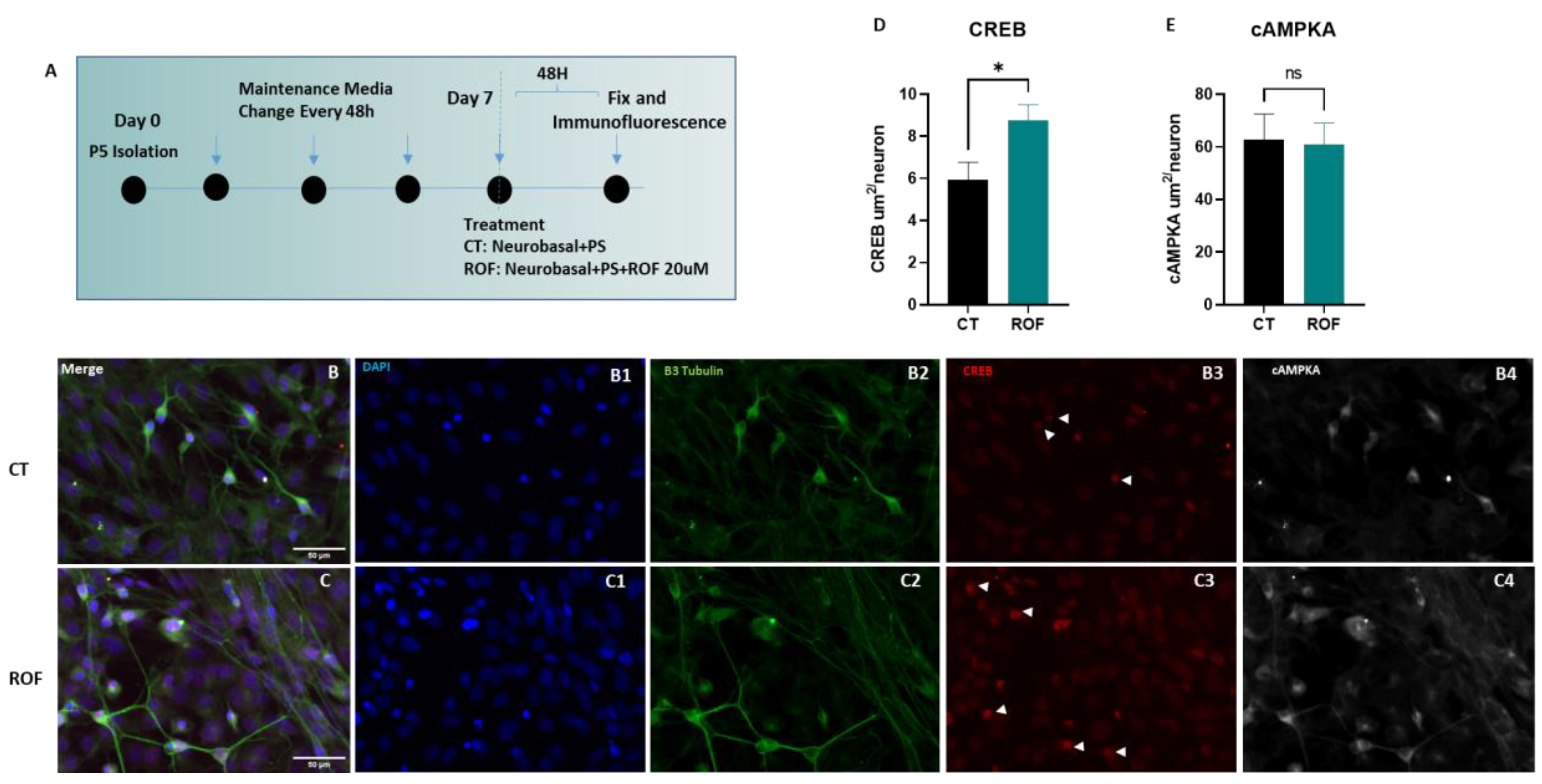
2.1. Spinal Cord Cultures
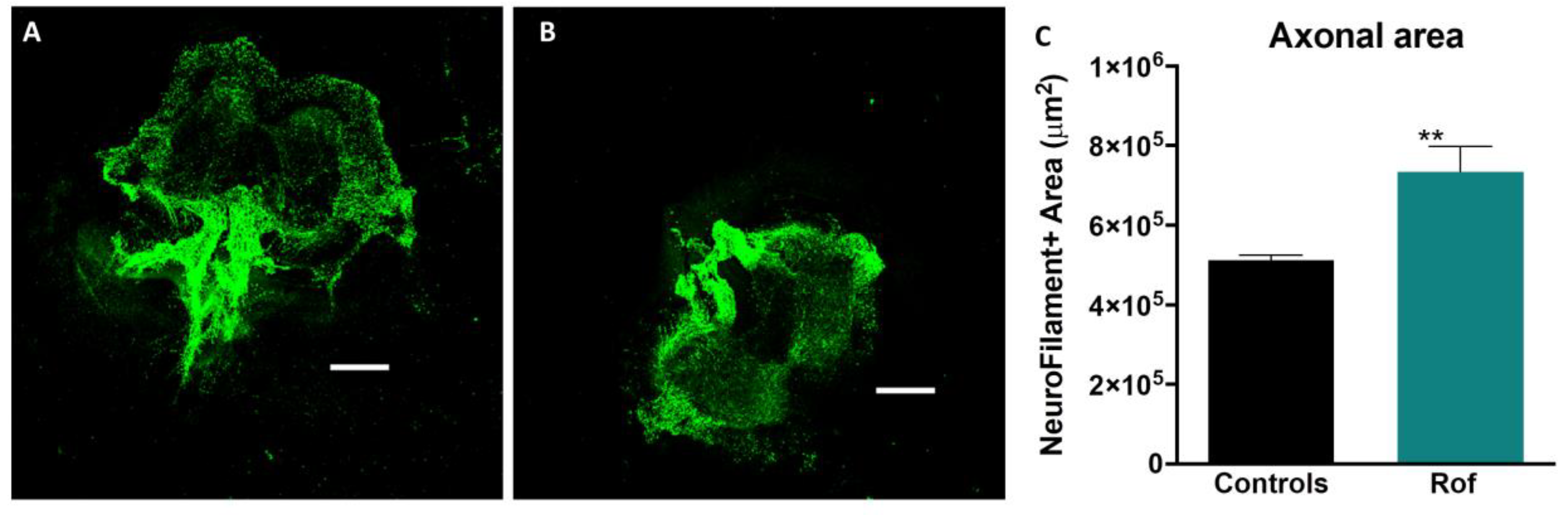
2.2. Organotypic Cultures
2.3. ELISA Assay
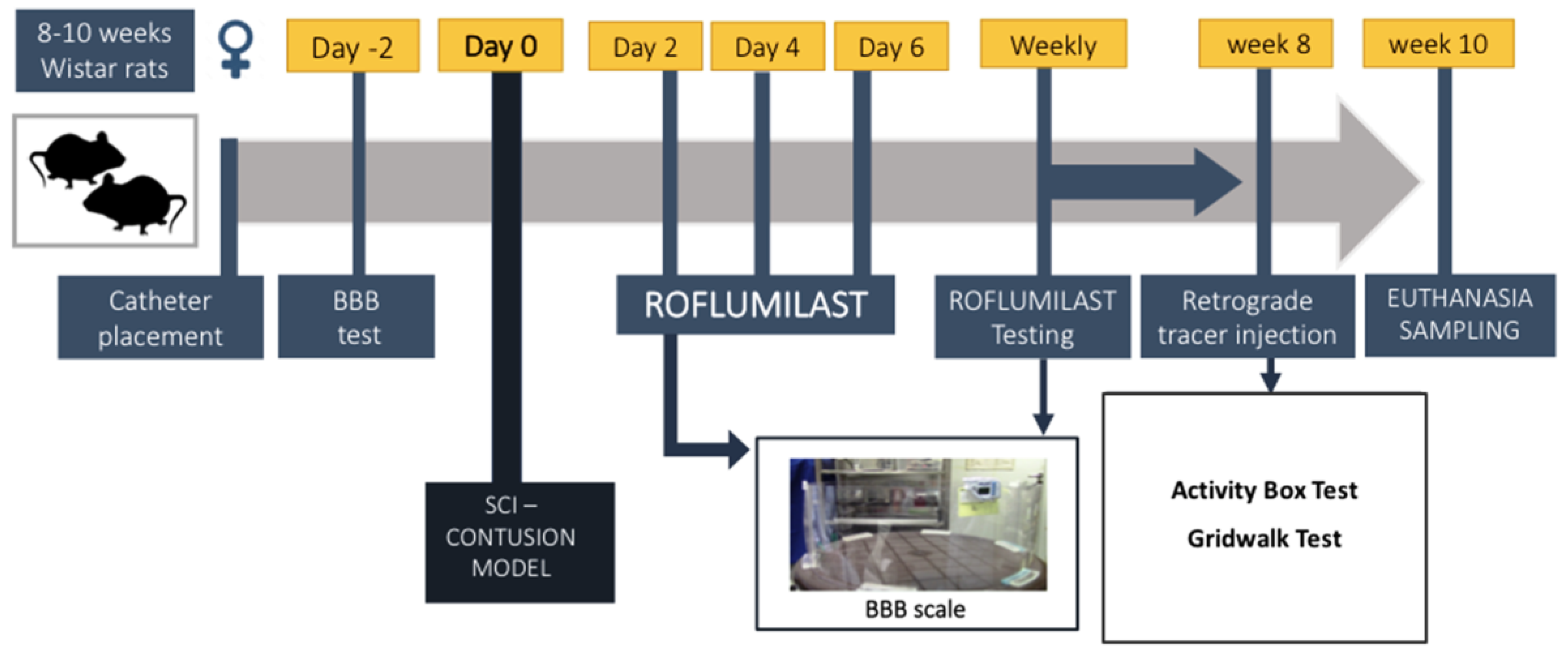
2.4. In Vivo Study Design
2.5. Spinal Cord Injury Model and Treatment
2.6. Behavioral Assessment
2.7. Fluoro-Gold–Retrograde Axonal Tracer
2.8. Serum Cytokine and VEGF Mediator Analysis
2.9. Immunohistochemistry Protocol
2.10. Immunofluorescence Analysis
2.11. Statistical Analysis
3. Results
3.1. Roflumilast Supports Survival and Increases Nuclear CREB Labeling in Spinal Cord Neurons In Vitro
3.2. Roflumilast Administration Promotes Axonal Growth In Vitro
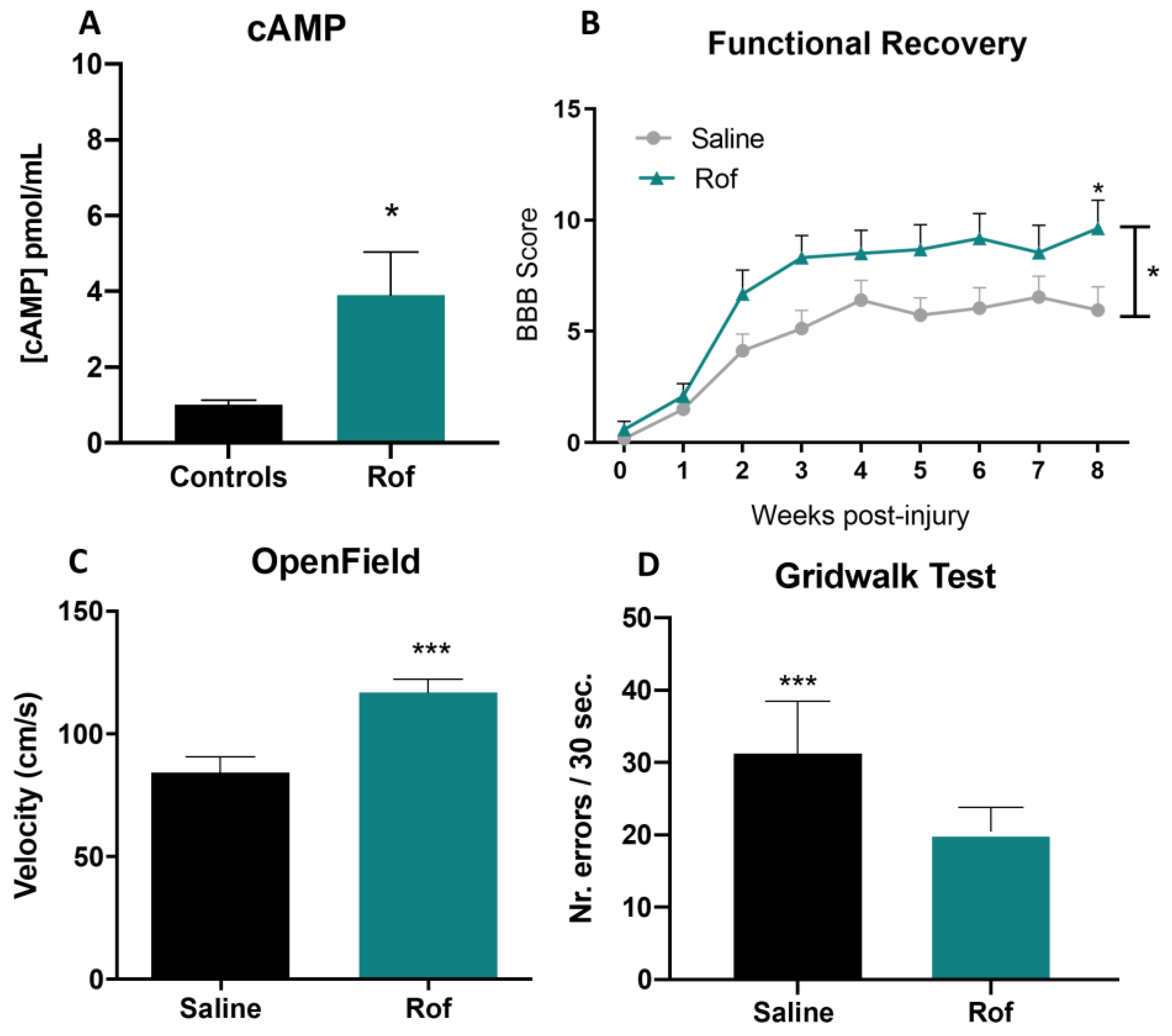
3.3. Roflumilast Treatment Increases Cyclic AMP Concentration In Vivo
3.4. Roflumilast Treatment Promotes Functional Recovery in a Thoracic Contusion Injury Model
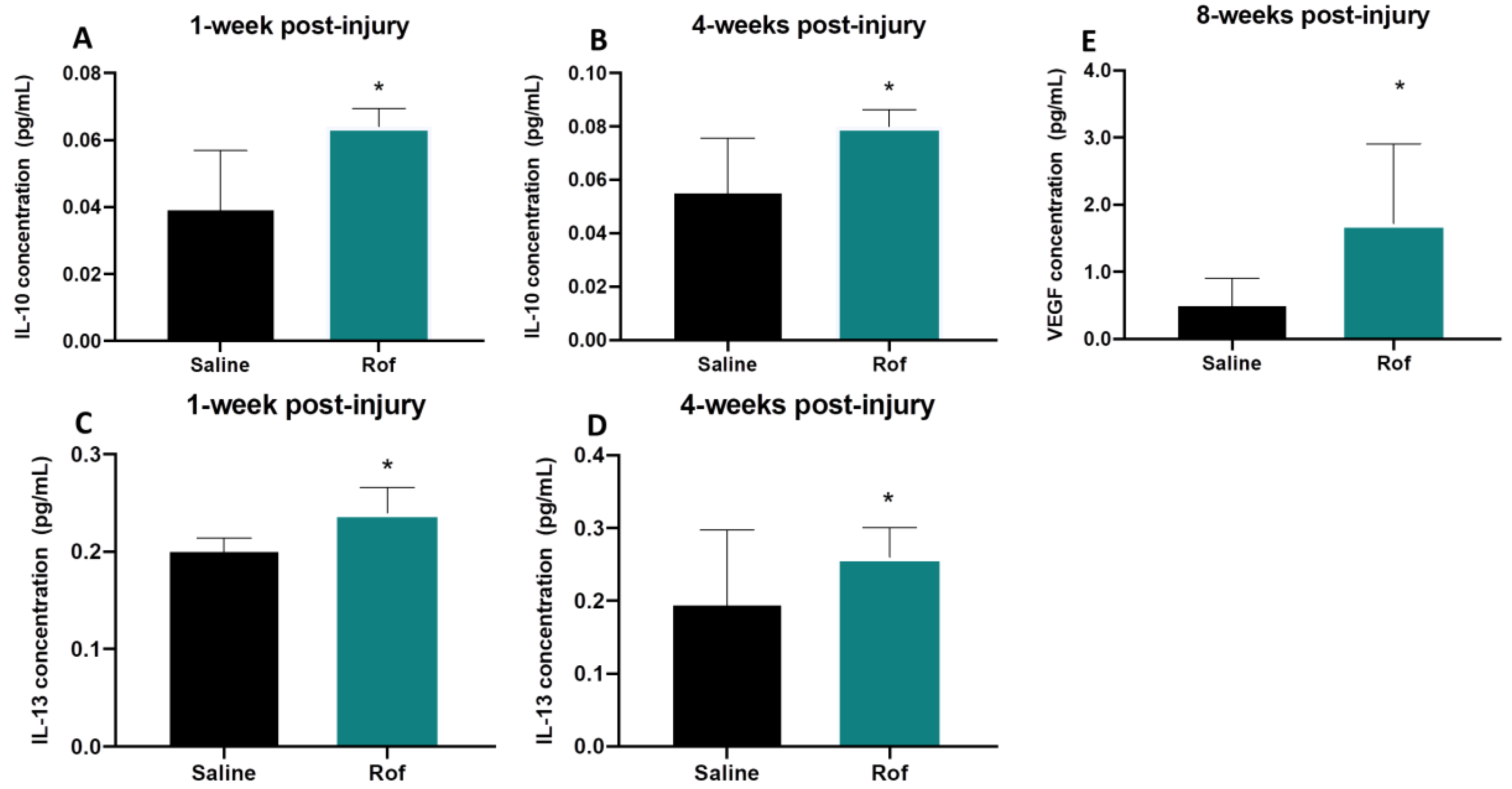
3.5. Roflumilast Treatment Induces an Anti-Inflammatory Environment
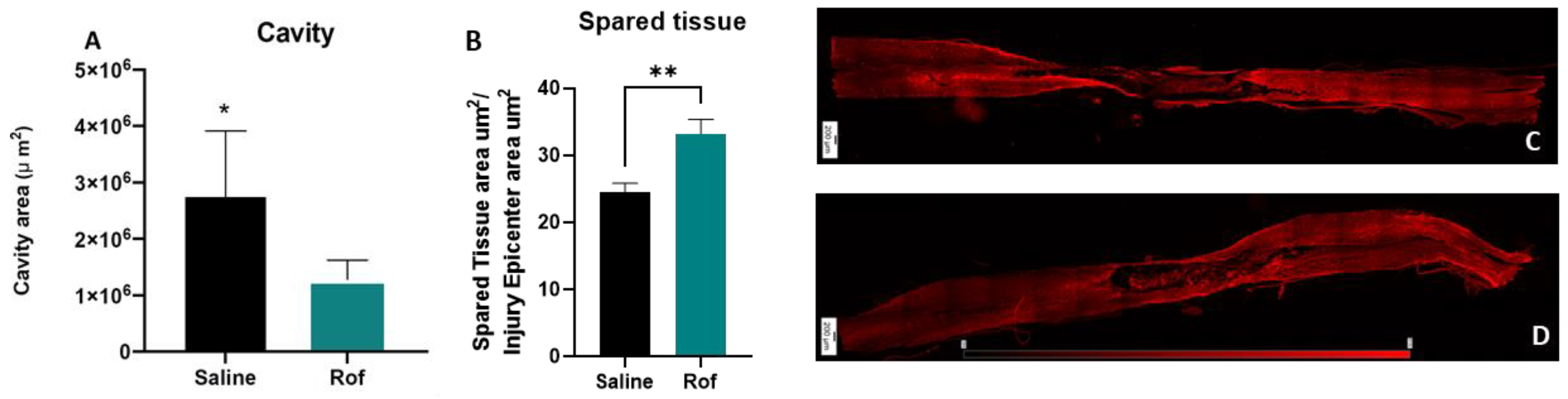
3.6. Roflumilast Treatment Reduces Cavity Size, Increases the Spared Tissue and Reduces Microglia Reactivity
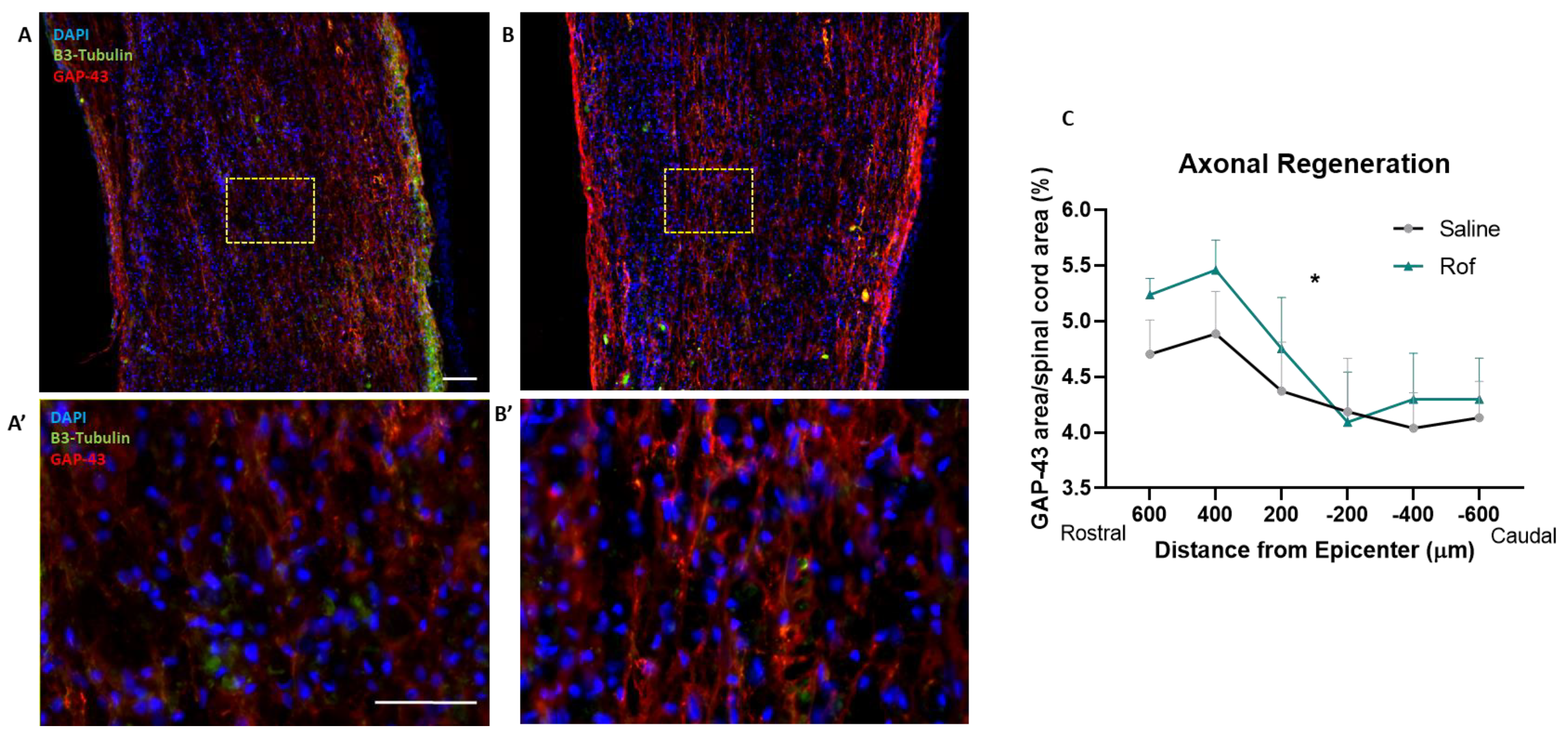
3.7. Roflumilast Treatment Increases Neuronal Regeneration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monteiro, S.; Pinho, A.G.; Macieira, M.; Serre-Miranda, C.; Cibrão, J.R.; Lima, R.; Soares-Cunha, C.; Vasconcelos, N.L.; Lentilhas-Graça, J.; Duarte-Silva, S.; et al. Splenic sympathetic signaling contributes to acute neutrophil infiltration of the injured spinal cord. J. Neuroinflamm. 2020, 17, 282. [Google Scholar] [CrossRef]
- Catapano, J.S.; Hawryluk, G.W.J.; Whetstone, W.; Saigal, R.; Ferguson, A.; Talbott, J.; Bresnahan, J.; Dhall, S.; Pan, J.; Beattie, M.; et al. Higher Mean Arterial Pressure Values Correlate with Neurological Improvement in Patients With Initially Complete Spinal Cord Injuries. World Neurosurg. 2016, 96, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Park, Y.J.; Song, S.Y.; Hwang, S.C.; Kim, K.T.; Kim, D.H. The importance of early surgical decompression for acute traumatic spinal cord injury. CiOS Clin. Orthop. Surg. 2018, 10, 448–454. [Google Scholar] [CrossRef]
- Canseco, J.A.; Karamian, B.A.; Bowles, D.R.; Markowitz, M.P.; DiMaria, S.L.; Semenza, N.C.; Leibensperger, M.R.; Smith, M.L.; Vaccaro, A.R. Updated Review: The Steroid Controversy for Management of Spinal Cord Injury. World Neurosurg. 2021, 150, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From basics to clinical: A comprehensive review on spinal cord injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Wei, Z.; Yao, X.; Shi, G.; Cheng, X.; Zhou, X.; Zhou, H.; Ning, G.; Kong, X.; Feng, S. Microenvironment Imbalance of Spinal Cord Injury. Cell Transplant. 2018, 27, 853–866. [Google Scholar] [CrossRef]
- Caroni, P.; Schwab, M.E. Two membrane protein fractions from rat central myelin with inhibitory properties for neurite growth and fibroblast spreading. J. Cell Biol. 1988, 106, 1281–1288. [Google Scholar] [CrossRef]
- Yamashita, T.; Higuchi, H.; Tohyama, M. The p75 receptor transduces the signal from myelin-associated glycoprotein to Rho. J. Cell Biol. 2002, 157, 565–570. [Google Scholar] [CrossRef]
- Dergham, P.; Ellezam, B.; Essagian, C.; Avedissian, H.; Lubell, W.D.; McKerracher, L. Rho Signaling Pathway Targeted to Promote Spinal Cord Repair. J. Neurosci. 2002, 22, 6570–6577. [Google Scholar] [CrossRef]
- Cai, D.; Qiu, J.; Cao, Z.; McAtee, M.; Bregman, B.S.; Filbin, M.T. Neuronal Cyclic AMP Controls the Developmental Loss in Ability of Axons to Regenerate. J. Neurosci. 2001, 21, 4731–4739. [Google Scholar] [CrossRef]
- Hannila, S.S.; Filbin, M.T. The role of cyclic AMP signaling in promoting axonal regeneration after spinal cord injury. Exp. Neurol. 2008, 209, 321–332. [Google Scholar] [CrossRef]
- Pearse, D.D.; Pereira, F.C.; Marcillo, A.E.; Bates, M.L.; Berrocal, Y.A.; Filbin, M.T.; Bunge, M.B. cAMP and Schwann cells promote axonal growth and functional recovery after spinal cord injury. Nat. Med. 2004, 10, 610–616. [Google Scholar] [CrossRef]
- Whitaker, C.M.; Beaumont, E.; Wells, M.J.; Magnuson, D.S.K.; Hetman, M.; Onifer, S.M. Rolipram attenuates acute oligodendrocyte death in the adult rat ventrolateral funiculus following contusive cervical spinal cord injury. Neurosci. Lett. 2008, 438, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, E.; Whitaker, C.M.; Burke, D.A.; Hetman, M.; Onifer, S.M. Effects of Rolipram on Adult Rat Oligodendrocytes and Functional Recovery after Contusive Cervical Spinal Cord Injury. Neuroscience 2009, 163, 985–990. [Google Scholar] [CrossRef]
- Heckman, P.R.A.; Wouters, C.; Prickaerts, J. Phosphodiesterase Inhibitors as a Target for Cognition Enhancement in Aging and Alzheimer’s Disease: A Translational Overview. Curr. Pharm. Des. 2014, 21, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Fu, C.X.; Yang, X.Y.; Cui, C.; Yang, S.; Kuang, Y.; Guo, C.X.; Hu, P.; Pei, Q.; Yang, G.P. Pharmacokinetics of single- and multiple-dose roflumilast: An open-label, three-way crossover study in healthy Chinese volunteers. Drug Des. Devel. Ther. 2018, 12, 4047–4057. [Google Scholar] [CrossRef] [PubMed]
- Garnock-Jones, K.P. Roflumilast: A Review in COPD. Drugs 2015, 75, 1645–1656. [Google Scholar] [CrossRef]
- Lima, R.; Monteiro, S.; Lopes, J.P.; Barradas, P.; Vasconcelos, N.L.; Gomes, E.D.; Assunção-Silva, R.C.; Teixeira, F.G.; Morais, M.; Sousa, N.; et al. Systemic Interleukin-4 Administration after Spinal Cord Injury Modulates Inflammation and Promotes Neuroprotection. Pharmaceuticals 2017, 10, 83. [Google Scholar] [CrossRef]
- Vasconcelos, N.L.; Gomes, E.D.; Oliveira, E.P.; Silva, C.J.; Lima, R.; Sousa, N.; Salgado, A.J.; Silva, N.A. Combining neuroprotective agents: Effect of riluzole and magnesium in a rat model of thoracic spinal cord injury. Spine J. 2016, 16, 1015–1024. [Google Scholar] [CrossRef]
- Lima, R.; Gomes, E.D.; Cibrão, J.R.; Rocha, L.A.; Assunção-Silva, R.C.; Rodrigues, C.S.; Neves-Carvalho, A.; Monteiro, S.; Salgado, A.J.; Silva, N.A. Levetiracetam treatment leads to functional recovery after thoracic or cervical injuries of the spinal cord. npj Regen. Med. 2021, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.A.; Sousa, R.A.; Fraga, J.S.; Fontes, M.; Leite-Almeida, H.; Cerqueira, R.; Almeida, A.; Sousa, N.; Reis, R.L.; Salgado, A. J Benefits of spine stabilization with biodegradable scaffolds in spinal cord injured rats. Tissue Eng. Part C Methods 2013, 19, 101–108. [Google Scholar] [CrossRef]
- Gomes, E.D.; Mendes, S.S.; Assunção-Silva, R.C.; Teixeira, F.G.; Pires, A.O.; Anjo, S.I.; Manadas, B.; Leite-Almeida, H.; Gimble, J.M.; Sousa, N.; et al. Co-Transplantation of Adipose Tissue-Derived Stromal Cells and Olfactory Ensheathing Cells for Spinal Cord Injury Repair. Stem Cells 2018, 36, 696–708. [Google Scholar] [CrossRef] [PubMed]
- Metz, G.A.S.; Merkler, D.; Dietz, V.; Schwab, M.E.; Fouad, K. Efficient testing of motor function in spinal cord injured rats. Brain Res. 2000, 883, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Havton, L.A. Retrogradely Transported Fluorogold Accumulates in Lysosomes of Neurons and is Detectable Ultrastructurally using Post-embedding Immuno-gold Methods. J. Neurosci. 2009, 184, 42–47. [Google Scholar] [CrossRef]
- Lima, R.; Monteiro, S.; Gomes, E.D.; Vasconcelos, N.L.; Assunção-Silva, R.; Morais, M.; Salgado, A.J.; Silva, N.A. Citalopram Administration Does Not Promote Function or Histological Recovery after Spinal Cord Injury. Int. J. Mol. Sci. 2020, 21, 5062. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.P.; Cafferty, W.B.J.; Budel, S.O.; Strittmatter, S.M. Extracellular regulators of axonal growth in the adult central nervous system. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1593–1610. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.P.; Fournier, A.; GrandPré, T.; Strittmatter, S.M. Myelin-associated glycoprotein as a functional ligand for the Nogo-66 receptor. Science 2002, 297, 1190–1193. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.E. Nogo and axon regeneration. Curr. Opin. Neurobiol. 2004, 14, 118–124. [Google Scholar] [CrossRef]
- Filbin, M.T. Myelin-associated inhibitors of axonal regeneration in the adult mammalian. CNS. Nat. Rev. Neurosci. 2003, 4, 703–713. [Google Scholar] [CrossRef]
- Cai, D.; Deng, K.; Mellado, W.; Lee, J.; Ratan, R.R.; Filbin, M.T. Arginase I and polyamines act downstream from cyclic AMP in overcoming inhibition of axonal growth MAG and myelin in vitro. Neuron 2002, 35, 711–719. [Google Scholar] [CrossRef]
- Bielekova, B.; Richert, N.; Howard, T.; Packer, A.N.; Blevins, G.; Ohayon, J.; McFarland, H.F.; Stürzebecher, C.S.; Martin, R. Treatment with the phosphodiesterase type-4 inhibitor rolipram fails to inhibit blood-brain barrier disruption in multiple sclerosis. Mult. Scler. J. 2009, 15, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Cortijo, J.; Iranzo, A.; Milara, X.; Mata, M.; Cerdá-Nicolás, M.; Ruiz-Saurí, A.; Tenor, H.; Hatzelmann, A.; Morcillo, E.J. Roflumilast, a phosphodiesterase 4 inhibitor, alleviates bleomycin-induced lung injury. Br. J. Pharmacol. 2009, 156, 534. [Google Scholar] [CrossRef] [PubMed]
- Nikulina, E.; Tidwell, J.L.; Dai, H.N.; Bregman, B.S.; Filbin, M.T. The phosphodiesterase inhibitor rolipram delivered after a spinal cord lesion promotes axonal regeneration and functional recovery. Proc. Natl. Acad. Sci. USA 2004, 101, 8786–8790. [Google Scholar] [CrossRef]
- Qiu, J.; Cai, D.; Dai, H.; McAtee, M.; Hoffman, P.N.; Bregman, B.S.; Filbin, M.T. Spinal axon regeneration induced by elevation of cyclic AMP. Neuron 2002, 34, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.; Bradke, F.; Tessier-Lavigne, M.; Basbaum, A.I. Regeneration of sensory axons within the injured spinal cord induced by intraganglionic cAMP elevation. Neuron 2002, 34, 885–893. [Google Scholar] [CrossRef]
- Gordon, T. The role of neurotrophic factors in nerve regeneration. Neurosurg. Focus 2009, 26, E3. [Google Scholar] [CrossRef]
- Cui, Q.; So, K.F. Involvement of cAMP in neuronal survival and axonal regeneration. Anat. Sci. Int. 2004, 79, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Siddiq, M.M.; Hannila, S.S. Looking downstream: The role of cyclic AMP-regulated genes in axonal regeneration. Front. Mol. Neurosci. 2015, 8, 26. [Google Scholar] [CrossRef]
- Batty, N.J.; Fenrich, K.K.; Fouad, K. The role of cAMP and its downstream targets in neurite growth in the adult nervous system. Neurosci. Lett. 2016, 652, 56–63. [Google Scholar] [CrossRef]
- Zhou, G.; Wang, Z.; Han, S.; Chen, X.; Li, Z.; Hu, X.; Li, Y.; Gao, J. Multifaceted Roles of cAMP Signaling in the Repair Process of Spinal Cord Injury and Related Combination Treatments. Front. Mol. Neurosci. 2022, 15, 808510. [Google Scholar] [CrossRef]
- Fabbri, L.M.; Beghé, B.; Yasothan, U.; Kirkpatrick, P. Roflumilast. Nat. Rev. Drug Discov. 2010, 9, 761–762. [Google Scholar] [CrossRef] [PubMed]
- Zebda, R.; Paller, A.S. Phosphodiesterase 4 inhibitors. J. Am. Acad. Dermatol. 2018, 78, S43–S52. [Google Scholar] [CrossRef] [PubMed]
- Pagès, L.; Gavaldà, A.; Lehner, M.d. PDE4 inhibitors: A review of current developments (2005–2009). Expert Opin. Ther. Pat. 2009, 19, 1501–1519. [Google Scholar] [CrossRef]
- Li, H.; Zuo, J.; Tang, W. Phosphodiesterase-4 inhibitors for the treatment of inflammatory diseases. Front. Pharmacol. 2018, 9, 1048. [Google Scholar] [CrossRef] [PubMed]
- Brustein, E.; Rossignol, S. Recovery of locomotion after ventral and ventrolateral spinal lesions in the cat. I. Deficits and adaptive mechanisms. J. Neurophysiol. 1998, 80, 1245–1267. [Google Scholar] [CrossRef] [PubMed]
- Z’Graggen, W.J.; Metz, G.A.S.; Kartje, G.L.; Thallmair, M.; Schwab, M.E. Functional Recovery and Enhanced Corticofugal Plasticity after Unilateral Pyramidal Tract Lesion and Blockade of Myelin-Associated Neurite Growth Inhibitors in Adult Rats. J. Neurosci. 1998, 18, 4744–4757. [Google Scholar] [CrossRef] [PubMed]
- Muir, G.D.; Whishaw, I.Q. Red nucleus lesions impair overground locomotion in rats: A kinetic analysis. Eur. J. Neurosci. 2000, 12, 1113–1122. [Google Scholar] [CrossRef]
- Rabe, K.F.; Watz, H.; Baraldo, S.; Pedersen, F.; Biondini, D.; Bagul, N.; Hanauer, G.; Göhring, U.M.; Purkayastha, D.; Román, J.; et al. Anti-inflammatory effects of roflumilast in chronic obstructive pulmonary disease (ROBERT): A 16-week, randomized, placebo-controlled trial. Lancet Respir. Med. 2018, 6, 827–836. [Google Scholar] [CrossRef]
- Salvator, H.; Buenestado, A.; Brollo, M.; Naline, E.; Victoni, T.; Longchamp, E.; Tenor, H.; Grassin-Delyle, S.; Devillier, P. Clinical Relevance of the Anti-inflammatory Effects of Roflumilast on Human Bronchus: Potentiation by a Long-Acting Beta-2-Agonist. Front. Pharmacol. 2020, 11, 1981. [Google Scholar] [CrossRef]
- Martinez, F.O.; Helming, L.; Gordon, S. Alternative Activation of Macrophages: An Immunologic Functional Perspective. Annu. Rev. Immunol. 2009, 27, 451–483. [Google Scholar] [CrossRef]
- Gensel, J.C.; Zhang, B. Macrophage activation and its role in repair and pathology after spinal cord injury. Brain Res. 2015, 1619, 1–11. [Google Scholar] [CrossRef]
- Serezani, C.H.; Ballinger, M.N.; Aronoff, D.M.; Peters-Golden, M. Cyclic AMP: Master Regulator of Innate Immune Cell Function. Am. J. Respir. Cell Mol. Biol. 2008, 39, 127. [Google Scholar] [CrossRef] [PubMed]
- Moradi, K.; Golbakhsh, M.; Haghighi, F.; Afshari, K.; Nikbakhsh, R.; Khavandi, M.M.; Faghani, S.; Badripour, A.; Etemadi, A.; Ashraf-Ganjouei, A.; et al. Inhibition of phosphodiesterase IV enzyme improves locomotor and sensory complications of spinal cord injury via altering microglial activity: Introduction of Roflumilast as an alternative therapy. Int. Immunopharmacol. 2020, 86, 106743. [Google Scholar] [CrossRef]
- Thompson, C.D.; Zurko, J.C.; Hanna, B.F.; Hellenbrand, D.J.; Hanna, A. The Therapeutic Role of Interleukin-10 after Spinal Cord Injury. J. Neurotrauma 2013, 30, 1311–1324. [Google Scholar] [CrossRef] [PubMed]
- Florentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’garra, A. IL-10 inhibits cytokine production by activated macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar] [CrossRef]
- Fuchs, A.C.; Granowitz, E.V.; Shapiro, L.; Vannier, E.; Lonnemann, G.; Angel, J.B.; Kennedy, J.S.; Rabson, A.R.; Radwanski, E.; Affrime, M.B.; et al. Clinical, hematologic, and immunologic effects of interleukin-10 in humans. J. Clin. Immunol. 1996, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Balasingam, V.; Yong, V.W. Attenuation of Astroglial Reactivity by Interleukin-10. J. Neurosci. 1996, 16, 2945–2955. [Google Scholar] [CrossRef]
- Xin, J.; Wainwright, D.A.; Mesnard, N.A.; Serpe, C.J.; Sanders, V.M.; Jones, K.J. IL-10 within the CNS is necessary for CD4+ T cells to mediate neuroprotection. Brain. Behav. Immun. 2011, 25, 820–829. [Google Scholar] [CrossRef]
- Zhou, Z.; Peng, X.; Insolera, R.; Fink, D.J.; Mata, M. IL-10 promotes neuronal survival following spinal cord injury. Exp. Neurol. 2009, 220, 183–190. [Google Scholar] [CrossRef]
- Kolosowska, N.; Keuters, M.H.; Wojciechowski, S.; Keksa-Goldsteine, V.; Laine, M.; Malm, T.; Goldsteins, G.; Koistinaho, J.; Dhungana, H. Peripheral Administration of IL-13 Induces Anti-inflammatory Microglial/Macrophage Responses and Provides Neuroprotection in Ischemic Stroke. Neurotherapeutics 2019, 16, 1304–1319. [Google Scholar] [CrossRef]
- Moro, K.; Yamada, T.; Tanabe, M.; Takeuchi, T.; Ikawa, T.; Kawamoto, H.; Furusawa, J.I.; Ohtani, M.; Fujii, H.; Koyasu, S. Innate production of TH2 cytokines by adipose tissue-associated c-Kit+Sca-1+ lymphoid cells. Nature 2009, 463, 540–544. [Google Scholar] [CrossRef]
- Neill, D.R.; Wong, S.H.; Bellosi, A.; Flynn, R.J.; Daly, M.; Langford, T.K.A.; Bucks, C.; Kane, C.M.; Fallon, P.G.; Pannell, R.; et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 2010, 464, 1367. [Google Scholar] [CrossRef] [PubMed]
- Saenz, S.A.; Siracusa, M.C.; Perrigoue, J.G.; Spencer, S.P.; Urban, J.F.; Tocker, J.E.; Budelsky, A.L.; Kleinschek, M.A.; Kastelein, R.A.; Kambayashi, T.; et al. IL-25 elicits a multi-potent progenitor cell population that promotes Th2 cytokine responses. Nature 2010, 464, 1362. [Google Scholar] [CrossRef] [PubMed]
- Francos-Quijorna, I.; Amo-Aparicio, J.; Martinez-Muriana, A.; López-Vales, R. IL-4 drives microglia and macrophages toward a phenotype conducive for tissue repair and functional recovery after spinal cord injury. Glia 2016, 64, 2079–2092. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Xu, Y.; Pearse, D.D. Cyclic AMP is a key regulator of M1 to M2a phenotypic conversion of microglia in the presence of Th2 cytokines. J. Neuroinflamm. 2016, 13, 9. [Google Scholar] [CrossRef]
- Monteiro, S.; Salgado, A.J.; Silva, N.A. Immunomodulation as a neuroprotective strategy after spinal cord injury. Neural Regen. Res. 2018, 13, 423. [Google Scholar] [PubMed]
- Jin, K.; Zhu, Y.; Sun, Y.; Mao, X.O.; Xie, L.; Greenberg, D.A. Vascular endothelial growth factor (VEGF) stimulates neurogenesis in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11946. [Google Scholar] [CrossRef]
- Mackenzie, F.; Ruhrberg, C. Diverse roles for VEGF-A in the nervous system. Development 2012, 139, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Khaibullina, A.A.; Rosenstein, J.M.; Krum, J.M. Vascular endothelial growth factor promotes neurite maturation in primary CNS neuronal cultures. Dev. Brain Res. 2004, 148, 59–68. [Google Scholar] [CrossRef]
- Liu, Y.; Figley, S.; Spratt, S.K.; Lee, G.; Ando, D.; Surosky, R.; Fehlings, M.G. An engineered transcription factor which activates VEGF-A enhances recovery after spinal cord injury. Neurobiol. Dis. 2010, 37, 384–393. [Google Scholar] [CrossRef]
- Rong, W.; Wang, J.; Liu, X.; Jiang, L.; Wei, F.; Hu, X.; Han, X.; Liu, Z. Naringin treatment improves functional recovery by increasing BDNF and VEGF expression, inhibiting neuronal apoptosis after spinal cord injury. Neurochem. Res. 2012, 37, 1615–1623. [Google Scholar] [CrossRef]
- Bhuiyan, M.I.H.; Kim, J.C.; Hwang, S.N.; Lee, M.Y.; Kim, S.Y. Ischemic tolerance is associated with VEGF-C and VEGFR-3 signaling in the mouse hippocampus. Neuroscience 2015, 290, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Guaiquil, V.H.; Pan, Z.; Karagianni, N.; Fukuoka, S.; Alegre, G.; Rosenblatt, M.I. Vegf-B selectively regenerates injured peripheral neurons and restores sensory and trophic functions. Proc. Natl. Acad. Sci. USA 2014, 111, 17272–17277. [Google Scholar] [CrossRef] [PubMed]
- Han, M.-K.; Kim, M.; Bae, S.Y.; Kang, L.; Han, S.Y.; Lee, Y.-S.; Rha, J.H.; Kim, S.U.; Roh, J.-K. VEGF protects human cerebral hybrid neurons from in vitro ischemia. NeuroReport 2004, 15, 847–850. [Google Scholar] [CrossRef]
- Wen, Z.H.; Huang, S.Y.; Kuo, H.M.; Chen, C.T.; Chen, N.F.; Chen, W.F.; Tsui, K.H.; Liu, H.T.; Sung, C.S. Fumagillin attenuates spinal angiogenesis, neuroinflammation, and pain in neuropathic rats after chronic constriction injury. Biomedicines 2021, 9, 1187. [Google Scholar] [CrossRef] [PubMed]
- Yick, L.W.; So, K.F.; Cheung, P.T.; Wu, W.T. Lithium chloride reinforces the regeneration-promoting effect of chondroitinase ABC on rubrospinal neurons after spinal cord injury. J. Neurotrauma 2004, 21, 932–943. [Google Scholar] [CrossRef]
- McKillop, W.M.; York, E.M.; Rubinger, L.; Liu, T.; Ossowski, N.M.; Xu, K.; Hryciw, T.; Brown, A. Conditional Sox9 ablation improves locomotor recovery after spinal cord injury by increasing reactive sprouting. Exp. Neurol 2016, 283, 1–15. [Google Scholar] [CrossRef]
- Jakeman, L.B.; Chen, Y.; Lucin, K.M.; McTigue, D.M. Mice lacking L1 cell adhesion molecule have deficits in locomotion and exhibit enhanced corticospinal tract sprouting following mild contusion injury to the spinal cord. Eur. J. Neurosci. 2006, 23, 1997–2011. [Google Scholar] [CrossRef]
- Baptiste, D.C.; Austin, J.W.; Zhao, W.; Nahirny, A.; Sugita, S.; Fehlings, M.G. Systemic polyethylene glycol promotes neurological recovery and tissue sparing in rats after cervical spinal cord injury. J. Neuropathol. Exp. Neurol. 2009, 68, 661–676. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, C.S.; Lima, R.; Cibrão, J.R.; Gomes, E.D.; Fernandes, L.S.; Pinho, T.S.; Silva, D.; Campos, J.; Salgado, A.J.; Silva, N.A. Pre-Clinical Assessment of Roflumilast Therapy in a Thoracic Model of Spinal Cord Injury. Pharmaceutics 2023, 15, 1556. https://doi.org/10.3390/pharmaceutics15051556
Sousa CS, Lima R, Cibrão JR, Gomes ED, Fernandes LS, Pinho TS, Silva D, Campos J, Salgado AJ, Silva NA. Pre-Clinical Assessment of Roflumilast Therapy in a Thoracic Model of Spinal Cord Injury. Pharmaceutics. 2023; 15(5):1556. https://doi.org/10.3390/pharmaceutics15051556
Chicago/Turabian StyleSousa, Carla S., Rui Lima, Jorge R. Cibrão, Eduardo D. Gomes, Luís S. Fernandes, Tiffany S. Pinho, Deolinda Silva, Jonas Campos, António J. Salgado, and Nuno A. Silva. 2023. "Pre-Clinical Assessment of Roflumilast Therapy in a Thoracic Model of Spinal Cord Injury" Pharmaceutics 15, no. 5: 1556. https://doi.org/10.3390/pharmaceutics15051556
APA StyleSousa, C. S., Lima, R., Cibrão, J. R., Gomes, E. D., Fernandes, L. S., Pinho, T. S., Silva, D., Campos, J., Salgado, A. J., & Silva, N. A. (2023). Pre-Clinical Assessment of Roflumilast Therapy in a Thoracic Model of Spinal Cord Injury. Pharmaceutics, 15(5), 1556. https://doi.org/10.3390/pharmaceutics15051556