
Anti-Inflammatory Effects of Essential Oils from the Peels of Citrus Cultivars


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Essential Oil Extraction
2.4. Gas Chromatography–Mass Spectrometry Analysis
2.5. Cell Culture
2.6. Cell Cytotoxic Activity
2.7. Determination of NO Production
2.8. Measurement of Proinflammatory Cytokine Production
2.9. Statistical Analysis
3. Results
3.1. Chemical Composition of the Essential Oils
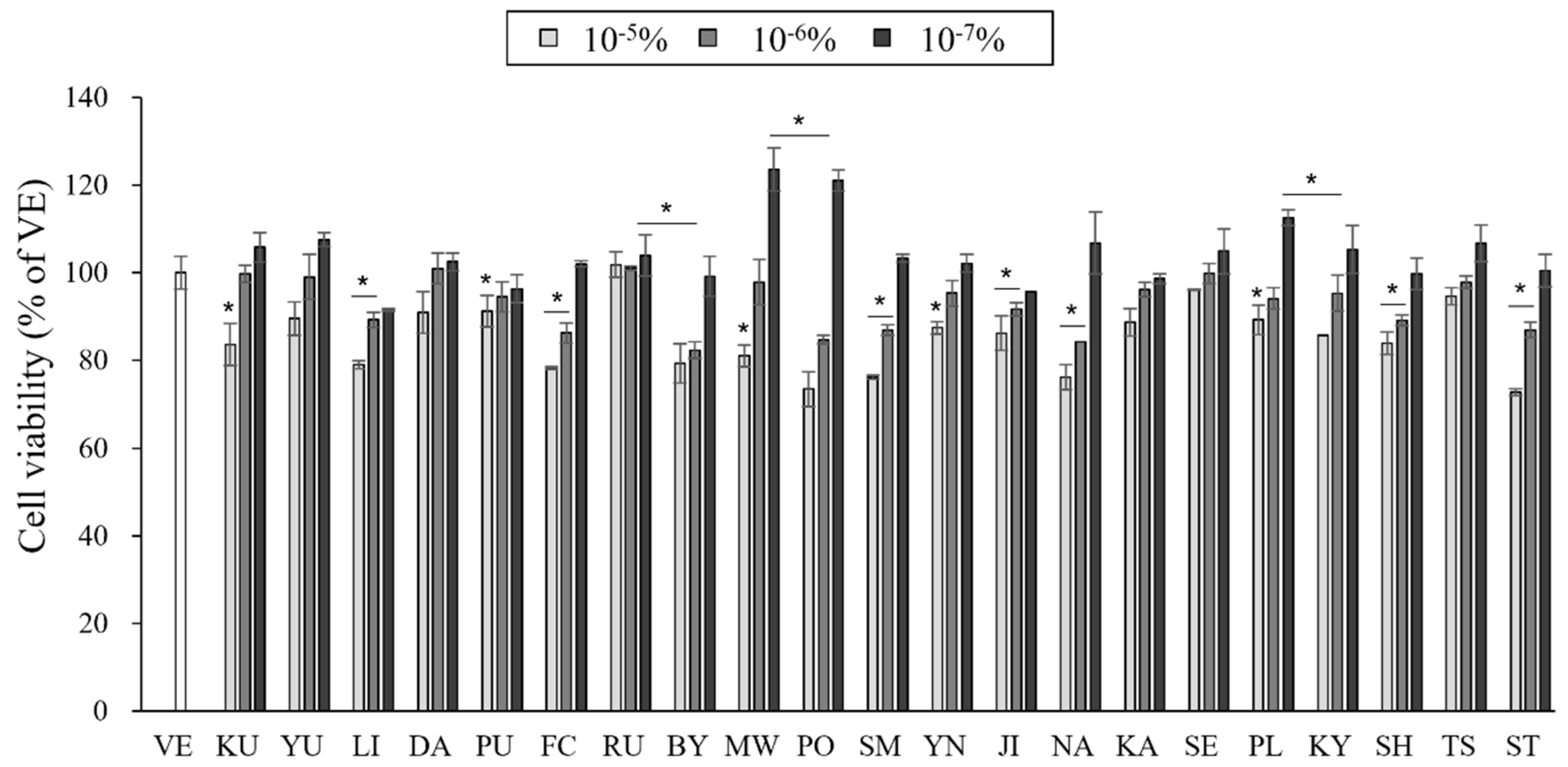
3.2. Effects of the Essential Oils on Cell Cytotoxicity
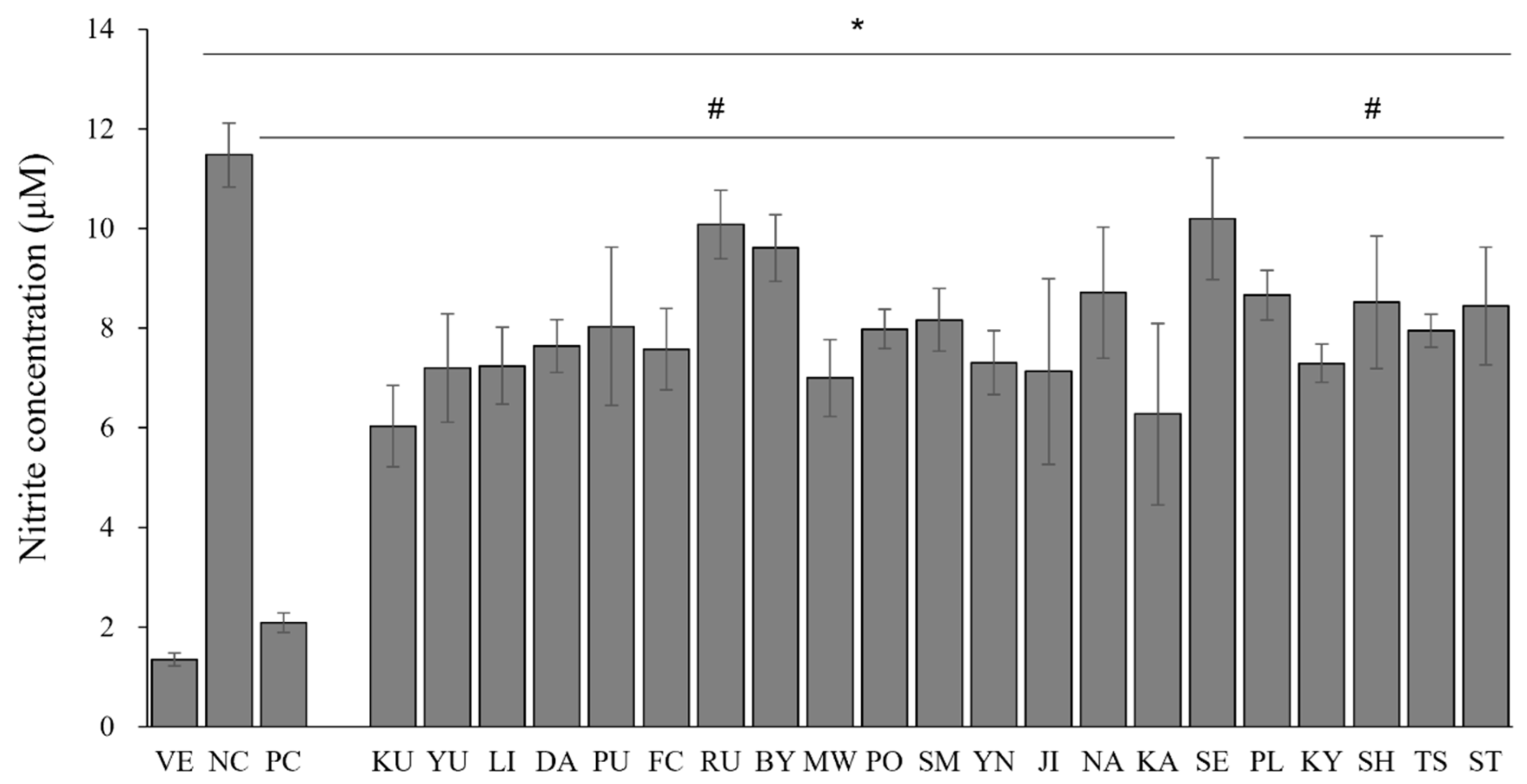
3.3. Effects of the Essential Oils on LPS-Induced NO Production in RAW 264.7 Cells
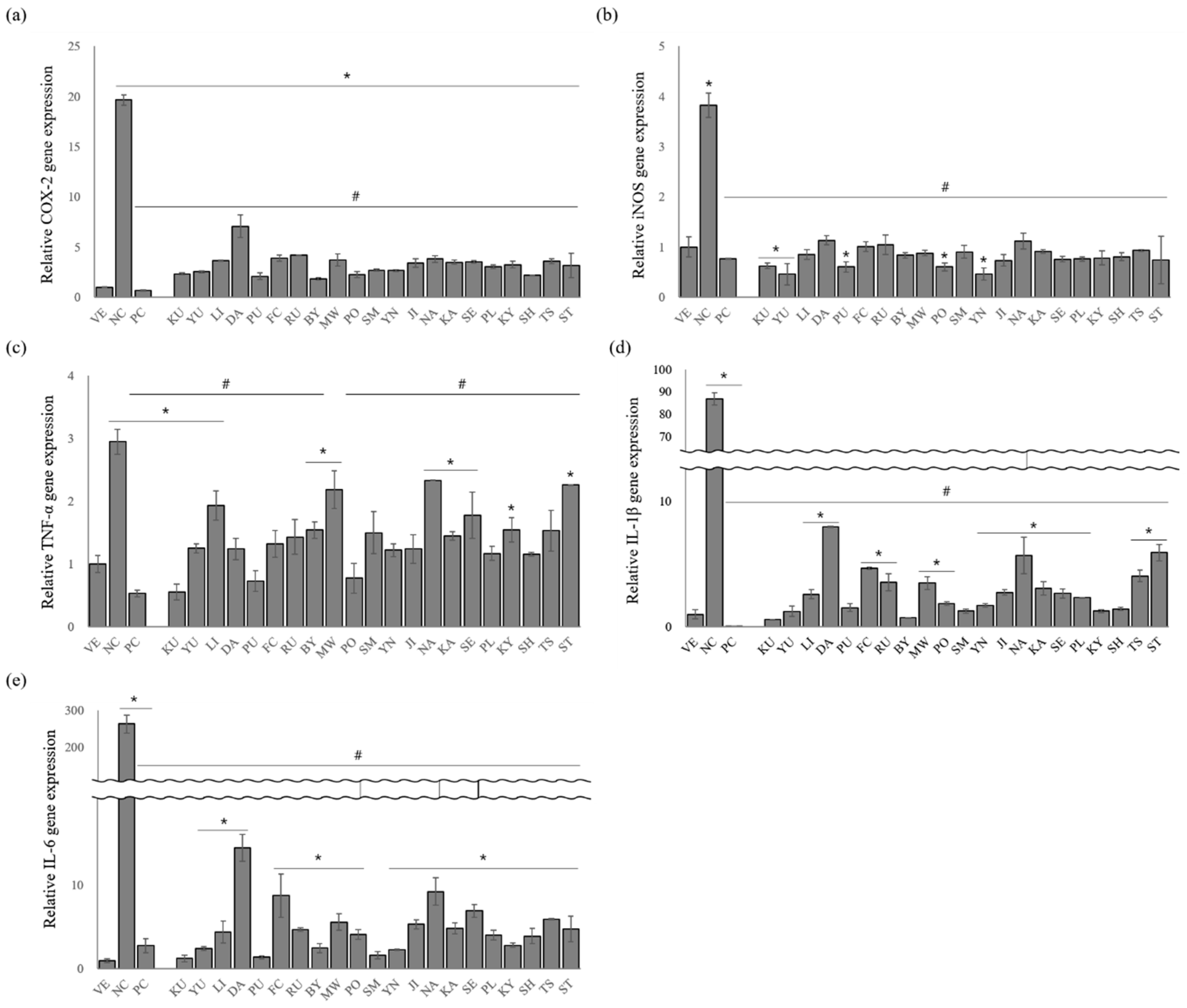
3.4. Effects of Essential Oils on the Gene Expression Levels of Proinflammatory Cytokines
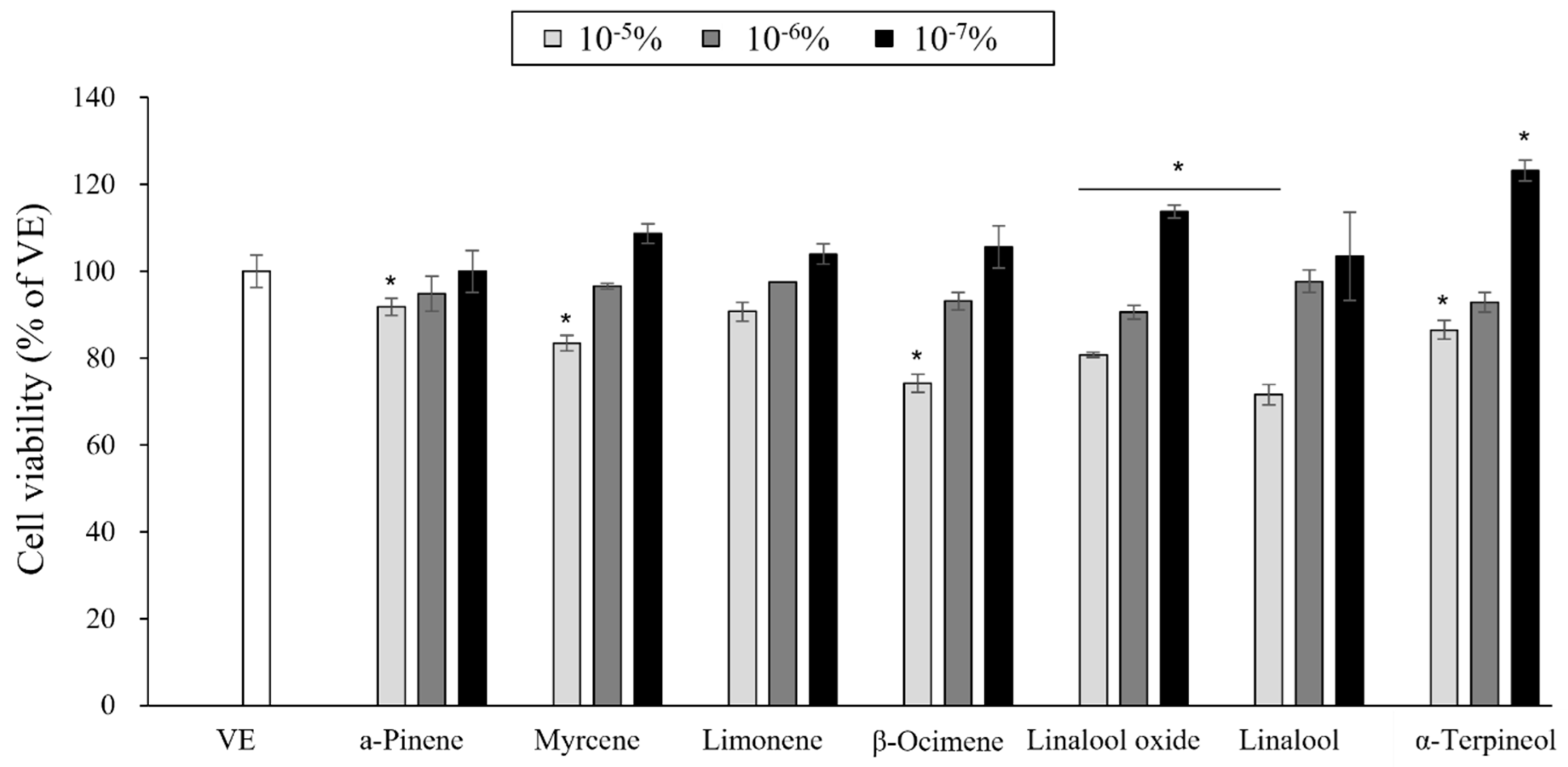
3.5. Cytotoxicity of the Single Compounds
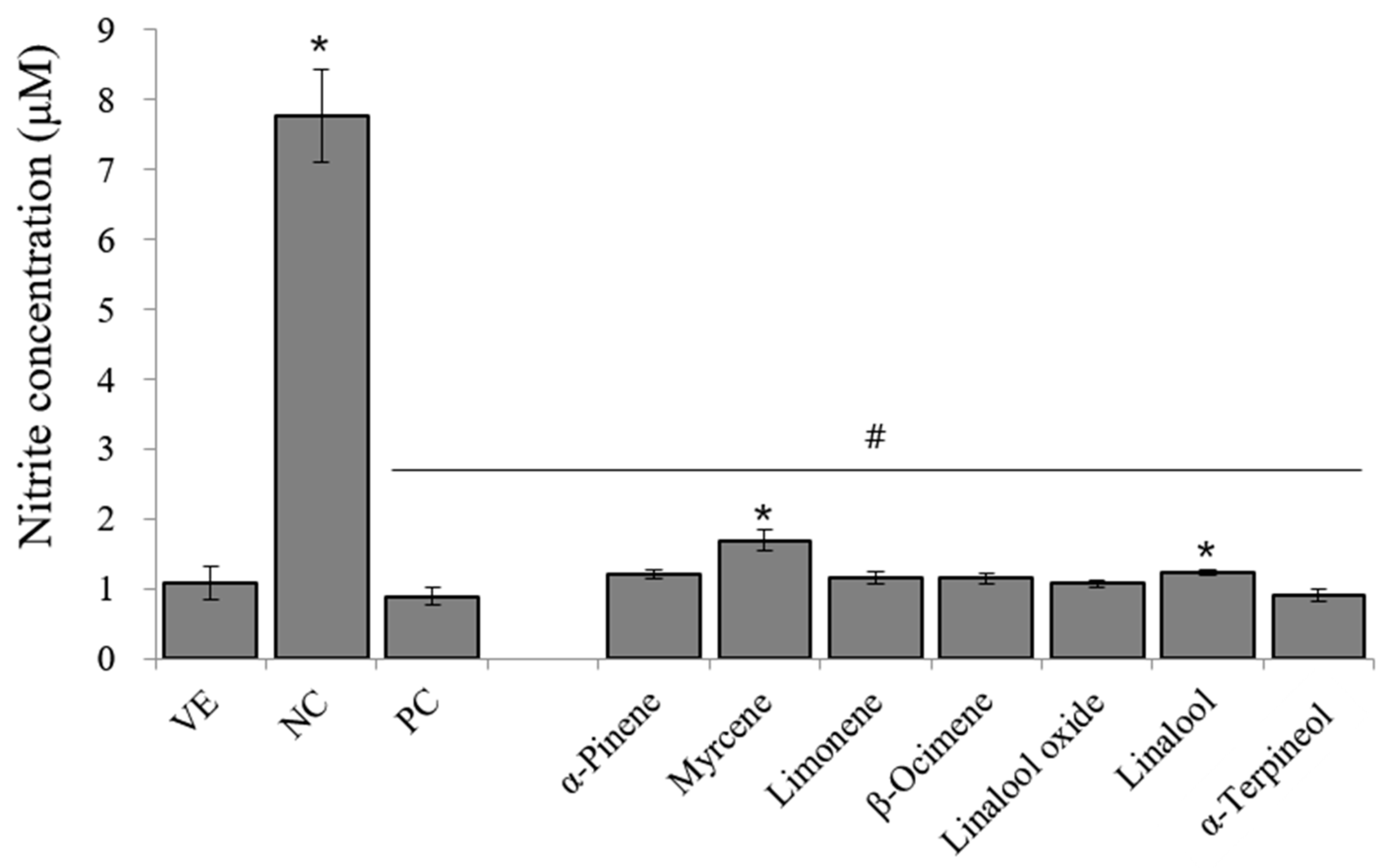
3.6. Effects of Single Compounds on LPS-Induced NO in RAW 264.7 Cells
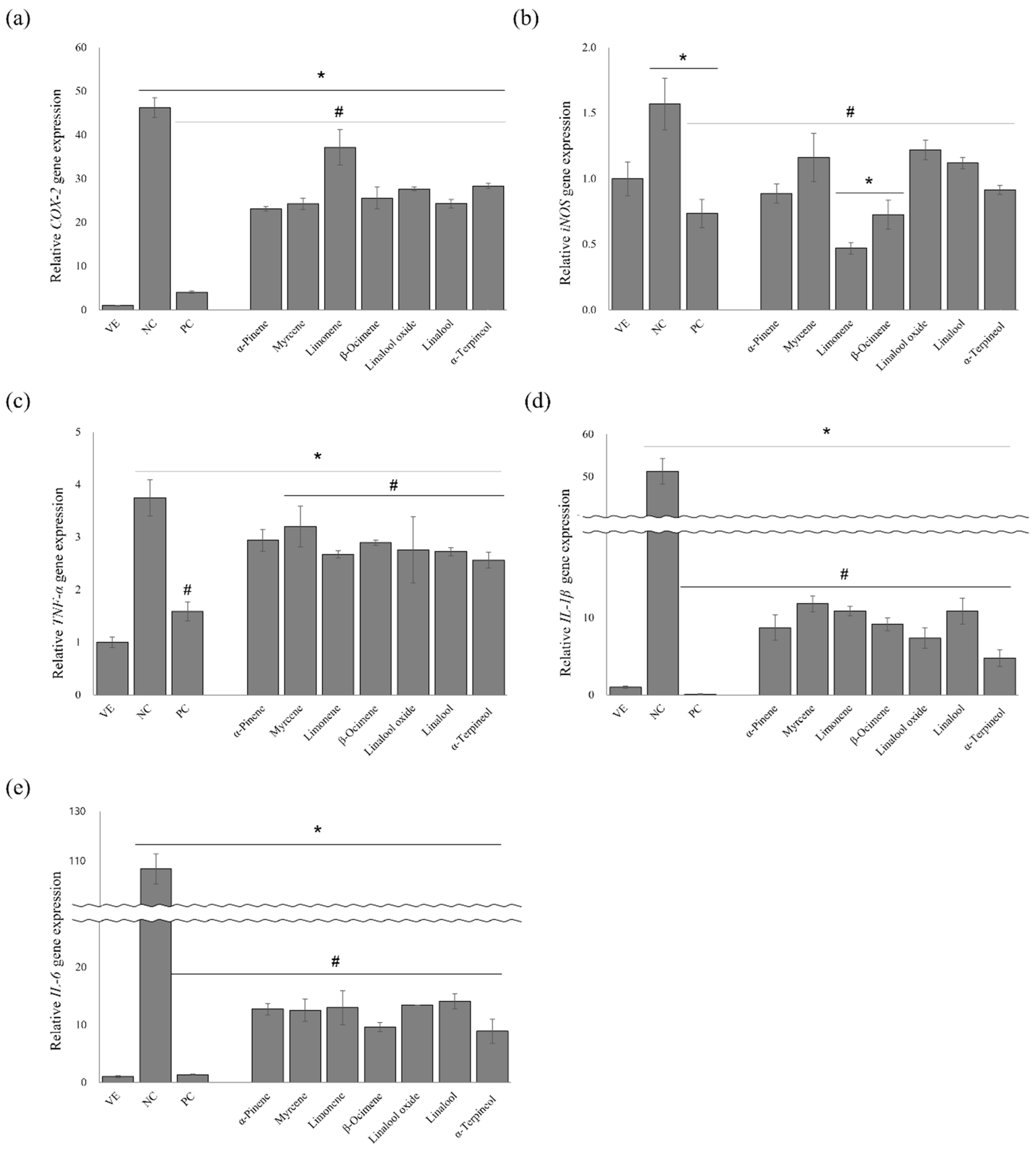
3.7. Effects of the Single Compounds on the Gene Expression Levels of Proinflammatory Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Choi, H.J.; Eun, J.S.; Park, Y.R.; Kim, D.K.; Li, R.; Moon, W.S.; Park, J.M.; Kim, H.S.; Cho, N.P.; Cho, S.D.; et al. Ikarisoside a inhibits inducible nitric oxide synthase in lipopolysaccharide-stimulated RAW 264.7 cells via p38 kinase and nuclear factor-κB signaling pathways. Eur. J. Pharmacol. 2008, 601, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Pierce, G.F. Macrophages: Important physiologic and pathologic sources of polypeptide growth factors. Am. J. Respir. Cell Mol. Biol. 1990, 2, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Sohn, D.S.; Cho, D.Y.; Kim, B.J.; Lim, Y.Y.; Kim, Y.H. Anti-inflammatory and antioxidant effects of Sophora flavescens root extraction in lipopolysaccharide-activated RAW 264.7 cells. Korean J. Med. Mycol. 2010, 15, 39–50. [Google Scholar]
- Ryu, J.H.; Ahn, H.; Kim, J.Y.; Kim, Y.K. Inhibitory activity of plant extracts on nitric oxide synthesis in LPS-activated macrophages. Phytother. Res. 2003, 17, 485–489. [Google Scholar] [CrossRef]
- Mu, M.M.; Chakravortty, D.; Sugiyama, T.; Koide, N.; Takahashi, K.; Mori, I.; Yoshida, T.; Yokochi, T. The nhibitory action of quercetin on lipopolysaccharide-induced nitric oxide production in RAW 264.7 macrophage cells. J. Endotoxin Res. 2001, 7, 431–438. [Google Scholar] [CrossRef]
- Harris, S.G.; Padilla, J.; Koumas, L.; Ray, D.; Phipps, R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002, 23, 144–150. [Google Scholar] [CrossRef]
- Mantelli, F.; Calder, V.L.; Bonini, S. The anti-inflammatory effects of therapies for ocular allergy. J. Ocul. Pharmacol. Ther. 2013, 29, 786–793. [Google Scholar] [CrossRef]
- Stüttgen, G. The present status of anti-inflammatory agents in dermatology. Drugs 1988, 36 (Suppl. 5), 43–48. [Google Scholar] [CrossRef]
- Russo, M.; Suraci, F.; Postorino, S.; Serra, D.; Roccotelli, A.; Agosteo, G.E. Essential oil chemical composition and antifungal effects on Sclerotium cepivorum of Thymus capitatus wild populations from Calabria, southern Italy. Rev. Bras. Farmacogn. 2013, 23, 239–248. [Google Scholar] [CrossRef]
- León-Méndez, G.; Pájaro-Castro, N.; Pájaro-Castro, E.; Torrenegra- Alarcón, M.; Herrera-Barros, A. Essential oils as a source of bioactive molecules. Rev. Colomb. Cienc. Químico-Farm. 2019, 48, 80–93. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-González, G.F.; Phumthum, M. Fundamental chemistry of essential oils and volatile organic compounds, methods of analysis and authentication. Plants 2022, 11, 789. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Shin, M.; Jung, S. Volatile compound analysis and anti-oxidant and anti-inflammatory effects of Oenanthe javanica, Perilla frutescens, and Zanthoxylum piperitum essential oils. Asian J. Beauty Cosmetol. 2017, 15, 355–366. [Google Scholar] [CrossRef]
- Yang, J.; Choi, W.; Jeung, E.; Kim, K.; Park, M. Anti-inflammatory effect of essential oil extracted from Pinus densiflora (Sieb. et Zucc.) wood on RBL-2H3 cells. J. Wood Sci. 2021, 67, 52. [Google Scholar] [CrossRef]
- Yang, J.; Choi, W.S.; Kim, K.J.; Eom, C.D.; Park, M.J. Investigation of active anti-inflammatory constituents of essential oil from Pinus koraiensis (Sieb. et Zucc.) wood in LPS-stimulated RBL-2H3 cells. Biomolecules 2021, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Sun, W.; Ke, C.; Wang, F.; Xue, Y.; Luo, Z.; Wang, X.; Zhang, J.; Zhang, Y. Anti-inflammatory activities of leaf oil from Cinnamomum subavenium in vitro and in vivo. BioMed Res. Int. 2019, 2019, 1823149. [Google Scholar] [CrossRef]
- An, B.S.; Kang, J.H.; Yang, H.; Jung, E.M.; Kang, H.S.; Choi, I.G.; Park, M.J.; Jeung, E.B. Anti-inflammatory effects of essential oils from Chamaecyparis obtusa via the cyclooxygenase-2 pathway in rats. Mol. Med. Rep. 2013, 8, 255–259. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Gaynor, R.B. Therapeutic potential of inhibition of the NK-κB pathway in the treatment of inflammation and cancer. J. Clin. Investig. 2001, 107, 135–142. [Google Scholar] [CrossRef]
- Sharma, K.; Mahato, N.; Lee, Y.R. Extraction, characterization and biological activity of citrus flavonoids. Rev. Chem. Eng. 2018, 35, 265–284. [Google Scholar] [CrossRef]
- Enejoh, O.S.; Ogunyemi, I.O.; Bala, M.S.; Oruene, I.S.; Suleiman, M.M.; Ambali, S.F. Ethnomedical importance of Citrus aurantifolia (Christm) Swingle. J. Pharm. Innov. 2015, 4, 1–6. [Google Scholar]
- Cin, P.; Gezer, C. Citrus fruits as a functional food and the relation with metabolic syndrome. J. Food Health Sci. 2017, 3, 49–58. [Google Scholar] [CrossRef]
- Takahashi, Y.; Inaba, N.; Kuwahara, S.; Kuki, W. Antioxidant effect of Citrus essential oil components on human low-density lipoprotein in vitro. Biosci. Biotechnol. Biochem. 2003, 67, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Cai, M.; Liu, Y.S.; Sun, P.L.; Luo, S.L. Antibacterial activity and mechanisms of essential oil from Citrus medica L. var. sarcodactylis. Molecules 2019, 24, 1577. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Yang, Z.Y.; Fan, G.; Ren, J.N.; Yin, K.J.; Pan, S.Y. Antidepressant-like effect of Citrus sinensis (L.) Osbeck essential oil and its main component limonene on mice. J. Agric. Food Chem. 2019, 67, 13817–13828. [Google Scholar] [CrossRef] [PubMed]
- Luro, F.; Neves, C.G.; Costantino, G.; Silva Gesteira, A.; Paoli, M.; Ollitrault, P.; Tomi, F.; Micheli, F.; Gibernau, M. Effect of environmental conditions on the yield of peel and composition of essential oils from citrus cultivated in Bahia (Brizil) and Corsica (France). Agronomy 2020, 10, 1256. [Google Scholar] [CrossRef]
- Kim, S.; Baik, J.S.; Oh, T.; Yoon, W.; Lee, N.H.; Hyun, C. Biological activities of Korean Citrus obovoides and Citrus natsudaidai essential oils against Acne-inducing bacteria. Biosci. Biotechnol. Biochem. 2008, 72, 2507–2513. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Yang, K.; Kim, S.S.; Park, S.M.; Park, K.J.; Kim, K.S.; Choi, Y.H.; Cho, K.K.; Hyun, C. Chemical composition and anti-inflammation activity of essential oils from Citrus unshiu flower. Nat. Prod. Commun. 2014, 9, 727–730. [Google Scholar] [CrossRef]
- Kim, K.; Ko, Y.; Yang, H.; Ham, Y.; Roh, S.W.; Jeon, Y.; Ahn, G.; Kanf, M.; Yoon, W.; Kim, D.; et al. Anti-inflammatory effect of essential oil and its constituents from fingered citron (Citrus medica L. var. sarcodactylis) through blocking JNK, ERK and NF-κB signaling pathways in LPS-activated RAW 264.7 cells. Food Chem. Toxicol. 2013, 57, 126–131. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.; Jang, S.; Kim, K.; Park, M. Inhibition of melanogenesis by essential oils from the citrus cultivars peels. Int. J. Mol. Sci. 2023, 24, 4207. [Google Scholar] [CrossRef]
- Chan, Y.Y.; Kim, K.H.; Cheah, S.H. Inhibitory effects of Sargassum polycystum on tyrosinase activity and and melanin formation in B16F10 murine melanoma cells. J. Ethnopharmacol. 2011, 137, 1183–1188. [Google Scholar] [CrossRef]
- Chou, S.T.; Peng, H.Y.; Hsu, J.C.; Lin, C.C.; Shih, Y. Achillea millefolium L. essential oil inhibits LPS-induced oxidative stress and nitric oxide production in RAW 264.7 macrophages. Int. J. Mol. Sci. 2013, 14, 12978–12993. [Google Scholar] [CrossRef]
- Caldwell, J.; Gardner, I.; Swales, N. An introduction to drug disposition: The basic principles of absorption, distribution, metabolism, and excretion. Toxicol. Pathol. 1995, 23, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Xaus, J.; Comalada, M.; Valledor, A.F.; Lloberas, J.; López-Soriano, F.; Argilés, J.M.; Bogdan, C.; Celada, A. LPS induces apoptosis in macrophages mostly through the autocrine production of TNF-α. Blood 2000, 95, 3823–3831. [Google Scholar] [CrossRef]
- Tian, Y.; Zhou, S.; Takeda, R.; Okazaki, K.; Sekita, M.; Sakamoto, K. Anti-inflammatory activities of amber extract in lipopolysaccharide-induced RAW 264.7 macrophages. Biomed. Pharmacother. 2021, 141, 111854. [Google Scholar] [CrossRef]
- Almas, I.; Innocent, E.; Machumi, F.; Kisinza, W. Effect of geographical location on yield and chemical composition of essential oils from three Eucalyptus species growing Tanzania. Asian J. Tradit. Med. 2019, 14, 1–12. [Google Scholar]
- Quintans, J.S.S.; Shanmugam, S.; Heimfarth, L.; Araújo, A.A.S.; Almeida, J.R.G.D.S.; Picot, L.; Quintans-Júnior, L.J. Monoterpenes modulating cytokines—A review. Food Chem. Toxicol. 2019, 123, 233–257. [Google Scholar] [CrossRef] [PubMed]
- Silveira e Sά, R.; Andrade, L.; Sousa, D. A review on anti-inflammatory activity of monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Yamazaki, C.; Yamazaki, M. A hydroxyl group of flavonoids affects oral anti-inflammatory activity and inhibition of systemic tumor necrosis factor-alpha production. Biosci. Biotechnol. Biochem. 2004, 68, 119–125. [Google Scholar] [CrossRef]
- Van Vuuren, S.F.; Viljoen, A.M. Antimicrobial activity of limonene enantiomers and 1,8-cineole alone and in combination. Flavour Fragr. J. 2007, 22, 540–544. [Google Scholar] [CrossRef]
- Tserennadmid, R.; Takó, M.; Galgóczy, L.; Papp, T.; Pesti, M.; Vágvölgyi, C.; Almássy, K.; Krisch, J. Anti yeast activities of some essemtial oils in growth medium, fruit juices and milk. Int. J. Food Microbiol. 2011, 144, 480–486. [Google Scholar] [CrossRef]
- Harris, R. Synergism in the essential oil world. Int. J. Aromather. 2002, 12, 179–186. [Google Scholar] [CrossRef]
- Frum, Y.; Viljoen, A.M. In vitro 5-lipoxygenase activity of three indigenous South Africa aromatic plants used in traditional healing and the stereospecific activity of limonene in the 5-lipoxygenase assay. J. Essent. Oil Res. 2006, 18, 85–88. [Google Scholar] [CrossRef]
- Erasto, P.; Viljoen, A.M. Limonene—A review: Biosynthetic, ecological and pharmacological relevance. Nat. Prod. Commun. 2008, 3, 1193–1202. [Google Scholar] [CrossRef]
- Singh, J.; Sharma, V.; Pandey, K.; Ahmed, S.; Kaur, M.; Sidhu, G. Horticultural classification of citrus cultivars. In Citrus, 1st ed.; IntechOpen: London, UK, 2021; p. 1. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Abbreviation | Specimen Information | Extraction Yield (mL/100 g, Dry Basis) |
|---|---|---|---|---|
| 1 | Citrus japonica Thunb. | KU | WTFRC10032742 | 9.52 ± 0.33 |
| 2 | Citrus junos Siebold ex Tanaka | YU | WTFRC10032743 | 6.93 ± 1.04 |
| 3 | Citrus limon (L.) Osbeck ‘Lisbon’ | LI | WTFRC10033803 | 5.05 ± 1.13 |
| 4 | Citrus maxima (Burm.) Merr. | DA | WTFRC10032725 | 6.75 ± 1.41 |
| 5 | Citrus maxima (Burm.) Merr. a | PU | WTFRC10032744 | 2.07 ± 0.60 |
| 6 | Citrus medica L. b | FC | WTFRC10033804 | 10.09 ± 0.01 |
| 7 | Citrus paradisi Macfad. ‘Redblush’ | RU | WTFRC10032741 | 3.70 ± 0.84 |
| 8 | Citrus platymamma hort. ex Tanaka | BY | WTFRC10032726 | 5.93 ± 0.22 |
| 9 | Citrus reticulata Blanco c | MW | WTFRC10032727 | 3.59 ± 1.00 |
| 10 | Citrus reticulata Blanco ‘Ponkan’ | PO | WTFRC10032734 | 16.25 ± 0.01 |
| 11 | Citrus reticulata Blanco d | SM | WTFRC10032740 | 4.85 ± 0.14 |
| 12 | Citrus sinensis (L.) Osbeck ‘Navel’ | YN | WTFRC10032732 | 6.39 ± 0.30 |
| 13 | Citrus sunki (Hayata) Yu. Tanaka | JI | WTFRC10032733 | 6.22 ± 0.69 |
| 14 | Citrus X aurantium L. e | NA | WTFRC10032737 | 2.03 ± 0.36 |
| 15 | Citrus X aurantium L. f | KA | WTFRC10032735 | 10.25 ± 0.40 |
| 16 | Citrus X aurantium L. g | SE | WTFRC10032729 | 10.43 ± 0.01 |
| 17 | Citrus X latifolia (Yu. Tanaka) Yu. Tanaka | PL | WTFRC10032736 | 5.04 ± 0.01 |
| 18 | Citrus unshiu X C. sinensis | KY | WTFRC10032739 | 5.49 ± 0.39 |
| 19 | (Citrus unshiu X C. sinensis) X C. reticulata | SH | WTFRC10032728 | 6.47 ± 0.18 |
| 20 | (Citrus unshiu X C. sinensis) X C. unshiu | TS | WTFRC10032731 | 6.05 ± 1.35 |
| 21 | ((Citrus unshiu X C. sinensis) X C. reticulata) X C. reticulata | ST | WTFRC10032730 | 6.96 ± 0.48 |
| Gene | Primer Sequence (5′–3′) |
|---|---|
| IL-1β | F: CAG GAT GAG GAC ATG AGC ACC R: CTC TGC AGA CTC AAA CTC CAC |
| IL-6 | F: GTA CTC CAG AAG ACC AGA GG R: TGC TGG TGA CAA CCA CGG CC |
| COX-2 | F: CGG ACT GGA TTC TAT GGT GAA A R: CTT GAA GTG GGT CAG GAT GTA G |
| iNOS | F: CCC TTC CGA AGT TTC TGG CAG CAG C R: GGC TGT CAG AGC CTC GTG GCT TTG G |
| TNF-α | F: TCC AGG CGG TGC CTA TGT R: CGA TCA CCC CGA AGT TCA GT |
| β-actin | F: CAG GTC ATC ACT ATT GGC AA R: AGG TCT TTA CGG ATG TCA AC |
| KI a | Compound Name | Area % | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KU | YU | LI | DA | PU | FC | RU | BY | MW | PO | SM | YN | JI | NA | KA | SE | PL | KY | SH | TS | ST | ||
| Monoterpene Hydrocarbons | ||||||||||||||||||||||
| 920 | α-Thujene | - | 0.17 | 0.12 | - | - | 0.64 | - | - | 0.06 | 0.09 | 0.09 | - | - | 0.03 | - | 0.02 | 0.19 | - | - | 0.07 | - |
| 926 | α-Pinene | 0.32 | 1.06 | 1.05 | 0.26 | 0.19 | 2.00 | 0.38 | 0.30 | 0.61 | 0.68 | 0.67 | 0.34 | 0.45 | 0.46 | 0.41 | 0.55 | 1.64 | 0.32 | 0.42 | 0.61 | 0.49 |
| 939 | α-Fenchene | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.02 | - | - | - | - |
| 941 | Camphene | - | - | 0.05 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.09 | - | - | - | - |
| 965 | Sabinene | - | 0.01 | 0.13 | - | - | 0.07 | 0.29 | 0.05 | 0.01 | 0.08 | 0.02 | 0.04 | 0.11 | - | 0.03 | - | 0.21 | 0.03 | 0.16 | 0.20 | 0.63 |
| 970 | β-Pinene | - | 0.50 | 6.03 | - | 0.22 | 1.62 | 0.14 | 0.85 | 0.25 | 0.26 | 0.34 | - | 1.98 | 0.21 | 0.86 | 0.12 | 7.55 | - | 0.04 | 0.27 | 0.36 |
| 986 | β-Myrcene | 1.34 | 1.25 | 0.95 | 21.62 | 28.09 | - | 1.25 | 19.48 | 1.21 | 1.17 | 1.27 | 1.31 | 1.17 | 1.09 | 1.23 | 1.31 | 0.84 | 1.23 | 1.36 | 1.18 | 1.05 |
| 1006 | Cosmene | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.03 |
| 1008 | α-Phellandrene | 0.01 | 0.49 | 0.05 | 0.02 | 0.03 | 0.04 | 0.05 | 0.02 | 0.02 | 0.03 | 0.02 | 0.04 | - | 0.04 | 0.02 | 0.02 | 0.07 | 0.01 | 0.04 | 0.03 | 0.02 |
| 1012 | 3-Carene | - | - | - | - | - | - | - | - | - | - | - | 0.07 | - | - | - | - | - | - | - | - | - |
| 1024 | α-Terpinene | - | 0.39 | 0.34 | - | - | 0.52 | 0.28 | 0.02 | 0.09 | 0.11 | 0.11 | 0.07 | 0.05 | 0.10 | 0.02 | 0.07 | 0.60 | 0.05 | 0.17 | 0.18 | 0.42 |
| 1034 | m-Cymene | - | 0.63 | 1.33 | - | 0.04 | 0.83 | 0.04 | - | 0.27 | 0.13 | 0.39 | 0.01 | - | 0.21 | - | 0.10 | 1.90 | - | - | 1.29 | 0.05 |
| 1041 | D-Limonene | 97.19 | 77.98 | 69.01 | 76.33 | 68.79 | 59.19 | 93.47 | 77.13 | 90.59 | 90.50 | 89.32 | 95.74 | 92.35 | 90.40 | 95.19 | 91.96 | 50.88 | 96.49 | 94.24 | 89.21 | 91.11 |
| 1047 | cis-β-Ocimene | - | - | 0.04 | 0.06 | - | 1.16 | - | 0.13 | - | - | - | - | 0.10 | - | 0.01 | - | 0.04 | - | 0.10 | - | - |
| 1058 | trans-β-Ocimene | 0.04 | 0.31 | 0.10 | 0.40 | 0.20 | 1.76 | 0.44 | 0.52 | 0.07 | 0.06 | 0.10 | 0.04 | 0.38 | 0.19 | 0.10 | 0.32 | 0.10 | 0.07 | 0.44 | 0.20 | 0.14 |
| 1068 | γ-Terpinene | 0.01 | 11.53 | 9.73 | 0.02 | 0.08 | 27.29 | 0.65 | 0.07 | 4.57 | 5.05 | 5.49 | 0.17 | 0.11 | 5.03 | 0.06 | 3.42 | 17.61 | 0.12 | 0.37 | 3.89 | 0.84 |
| 1090 | Terpinolene | 0.04 | 0.70 | 1.01 | 0.14 | 0.21 | 1.25 | 0.31 | 0.06 | 0.26 | 0.26 | 0.33 | 0.09 | 0.11 | 0.37 | 0.19 | 0.24 | 1.92 | 0.06 | 0.12 | 0.27 | 0.23 |
| 1093 | ρ,α-Dimethylstyrene | - | 0.03 | 0.01 | - | - | - | - | - | - | - | 0.01 | - | - | - | - | - | 0.03 | - | - | - | 0.04 |
| Oxygenated Monoterpene | ||||||||||||||||||||||
| 1004 | Octanal | - | - | 0.02 | - | - | - | - | - | - | 0.08 | - | 0.09 | - | 0.04 | - | - | - | 0.10 | 0.04 | 0.20 | - |
| 1042 | 1,8-Cineole | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.25 | - | - | - | - |
| 1078 | Linalool oxide | 0.02 | 0.04 | - | 0.25 | 0.28 | - | 0.24 | - | - | - | 0.01 | - | - | 0.11 | 0.18 | - | - | - | - | 0.02 | - |
| 1099 | Linalool | 0.14 | 1.97 | 0.37 | 0.10 | 0.18 | 0.06 | 0.23 | 0.42 | 0.42 | 0.71 | 0.12 | 0.35 | 0.73 | 0.17 | 0.57 | 0.33 | 0.72 | 0.11 | 0.45 | 0.29 | 0.95 |
| 1104 | Nonanal | - | - | 0.08 | - | - | - | - | - | 0.01 | - | - | - | - | 0.01 | - | - | - | 0.04 | 0.01 | 0.07 | - |
| 1116 | D-Fenchyl alcohol | - | - | 0.10 | - | - | - | - | - | - | - | - | - | 0.01 | - | - | - | 0.19 | - | - | - | - |
| 1119 | trans-ρ-2,8,1-Menthadienol | - | - | - | - | 0.06 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.07 |
| 1122 | (Z)-ρ-2-Menthen-1-ol | - | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.01 | 0.01 | 0.04 |
| 1146 | β-Terpineol | 0.05 | 0.10 | 0.11 | 0.03 | 0.06 | 0.01 | 0.07 | 0.02 | 0.05 | 0.02 | 0.03 | 0.12 | 0.03 | 0.12 | 0.09 | 0.17 | 0.13 | 0.08 | 0.10 | 0.04 | 0.06 |
| 1149 | Citronella | - | - | - | - | - | 0.01 | - | - | - | - | - | - | - | - | 0.01 | - | - | - | 0.03 | - | - |
| 1168 | α-Phellandren-8-ol | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.19 | - | - | - | - |
| 1169 | Borneol | - | - | 0.17 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.16 | - | - | - | - |
| 1179 | Terpinen-4-ol | 0.03 | 0.34 | 1.19 | 0.04 | 0.08 | 0.46 | 1.14 | 0.13 | 0.11 | 0.17 | 0.11 | 0.31 | 0.24 | 0.15 | 0.13 | 0.13 | 1.69 | 0.21 | 0.71 | 0.66 | 1.78 |
| 1186 | ρ-Cymen-8-ol | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.02 | - | - | - | - |
| 1194 | α-Terpineol | 0.28 | 1.01 | 3.28 | 0.19 | 0.37 | 0.66 | 0.57 | 0.28 | 0.37 | 0.26 | 0.26 | 0.58 | 0.49 | 0.77 | 0.58 | 0.96 | 5.85 | 0.45 | 0.51 | 0.41 | 0.51 |
| 1206 | Decanal | - | - | 0.02 | - | - | - | 0.11 | - | 0.07 | 0.08 | 0.01 | 0.10 | - | 0.07 | - | - | 0.08 | 0.10 | 0.32 | 0.31 | 0.20 |
| 1222 | trans-Carveol | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.24 |
| 1228 | Nerol | - | - | 0.08 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1238 | cis-Carveol | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.07 |
| 1243 | (Z)-Citral | - | - | 1.44 | - | 0.08 | 0.26 | - | - | - | - | - | - | - | - | - | - | 1.70 | - | - | - | - |
| 1251 | Carvone | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.10 |
| 1257 | Geraniol | - | - | 0.06 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1273 | (E)-Citral | - | - | 2.17 | - | 0.07 | 0.26 | - | - | - | - | - | - | - | - | - | - | 2.43 | - | - | - | - |
| 1288 | Bornyl acetate | - | - | - | - | - | - | - | - | - | - | - | 0.07 | - | - | - | - | - | - | - | - | - |
| Sesquiterpene Hydrocarbons | ||||||||||||||||||||||
| 1343 | δ-Elemene | 0.06 | 0.07 | - | 0.01 | - | - | - | 0.01 | 0.06 | 0.02 | 0.06 | - | 0.03 | - | 0.01 | - | 0.06 | - | - | - | - |
| 1384 | α-Copaene | 0.01 | 0.01 | - | 0.01 | - | - | 0.03 | - | 0.01 | - | 0.01 | - | 0.04 | 0.01 | 0.01 | 0.01 | - | - | - | 0.01 | - |
| 1395 | β-Elemene | 0.03 | 0.05 | - | 0.01 | - | - | 0.02 | 0.03 | 0.49 | 0.01 | 0.72 | 0.01 | 0.97 | 0.09 | 0.05 | 0.01 | 0.10 | 0.02 | - | 0.14 | 0.06 |
| 1430 | Caryophyllene | - | 0.13 | 0.08 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.26 | - | - | - | - |
| 1437 | γ-Elemene | - | 0.06 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1440 | trans-α-Bergamotene | - | 0.01 | 0.08 | - | 0.02 | 0.13 | - | - | - | - | - | - | - | 0.03 | - | - | 0.33 | - | - | 0.01 | - |
| 1457 | β-Farnesene | - | 0.21 | - | - | - | - | 0.01 | 0.03 | - | - | - | - | - | - | - | - | - | - | - | 0.05 | 0.01 |
| 1467 | Humulene | - | 0.03 | 0.01 | - | - | 0.02 | 0.03 | 0.01 | 0.06 | - | 0.08 | - | 0.11 | 0.01 | 0.01 | 0.01 | 0.05 | - | - | 0.02 | - |
| 1484 | α-Selinene | 0.01 | - | - | - | - | 0.01 | - | - | - | - | - | - | 0.01 | - | - | - | - | - | - | - | - |
| 1488 | α-Muurolene | 0.01 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1491 | Germacrene D | 0.07 | 0.03 | - | 0.11 | - | 0.14 | - | 0.03 | 0.04 | 0.01 | 0.05 | - | 0.03 | 0.01 | 0.01 | - | 0.03 | - | - | - | - |
| 1500 | β-Selinene | - | 0.15 | - | - | - | 0.01 | - | - | - | - | 0.01 | - | 0.02 | - | - | - | 0.02 | - | - | - | - |
| 1502 | Valencene | - | - | 0.01 | - | - | - | - | - | 0.01 | - | 0.01 | 0.33 | - | - | - | 0.05 | - | 0.06 | 0.01 | - | - |
| 1508 | α-Farnesene | - | 0.11 | - | 0.01 | - | 0.03 | - | 0.04 | 0.01 | - | 0.11 | 0.02 | 0.04 | 0.03 | - | - | 0.07 | - | 0.11 | 0.01 | - |
| 1513 | β-Bisabolene | 0.01 | - | 0.10 | - | - | 0.14 | - | 0.06 | 0.08 | - | - | - | - | 0.01 | - | - | 0.53 | - | - | - | - |
| 1528 | δ-Cadinene | 0.03 | 0.08 | - | 0.03 | - | 0.04 | 0.07 | 0.01 | 0.04 | - | 0.05 | 0.01 | 0.09 | 0.02 | 0.02 | 0.02 | - | 0.01 | 0.01 | - | - |
| Oxygenated Sesquiterpene | ||||||||||||||||||||||
| 1352 | α-Terpinyl acetate | - | - | 0.01 | - | - | - | 0.02 | - | - | - | - | - | 0.05 | - | - | - | - | - | - | - | - |
| 1362 | Neryl acetate | - | - | 0.25 | - | - | - | - | - | - | - | - | - | - | 0.06 | - | 0.04 | 0.89 | 0.06 | - | 0.10 | - |
| 1379 | Geranyl acetate | 0.05 | - | 0.24 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.28 | - | - | - | - |
| 1586 | Veridiflorol | - | 0.03 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1641 | β-Eudesmol | - | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1661 | τ-Muurolol | 0.01 | 0.03 | - | - | - | 0.01 | - | - | 0.01 | - | 0.01 | - | 0.01 | - | - | - | - | - | - | - | - |
| 1674 | α-Cadinol | 0.02 | 0.08 | - | 0.01 | - | 0.02 | - | - | 0.02 | - | 0.01 | - | 0.02 | - | - | - | - | - | - | - | - |
| Monoterpene hydrocarbons | 98.38 | 98.95 | 95.05 | 89.97 | 98.85 | 97.84 | 96.37 | 97.29 | 98.63 | 98.01 | 98.44 | 98.16 | 97.93 | 96.82 | 98.15 | 98.12 | 98.13 | 83.69 | 98.38 | 97.47 | 97.41 | |
| Oxygenated monoterpenes | 1.10 | 0.52 | 3.48 | 9.10 | 0.61 | 1.18 | 1.71 | 2.36 | 0.85 | 1.04 | 1.33 | 0.54 | 1.62 | 1.51 | 1.45 | 1.57 | 1.60 | 13.43 | 1.10 | 2.18 | 2.01 | |
| Sesquiterpene hydrocarbons | 0.09 | 0.23 | 0.96 | 0.29 | 0.18 | 0.02 | 0.53 | 0.16 | 0.22 | 0.81 | 0.04 | 1.11 | 0.37 | 1.35 | 0.21 | 0.11 | 0.10 | 1.46 | 0.09 | 0.13 | 0.25 | |
| Oxygenated sesquiterpenes | 0.06 | 0.08 | 0.16 | 0.49 | 0.01 | - | 0.03 | 0.02 | - | 0.03 | - | 0.02 | - | 0.08 | 0.06 | - | 0.04 | 1.17 | 0.06 | - | 0.10 | |
| Unknown compounds | 0.35 | 0.21 | 0.37 | 0.14 | 0.37 | 1.00 | 0.26 | 0.15 | 0.30 | 0.15 | 0.17 | 0.20 | 0.11 | 0.25 | 0.08 | 0.21 | 0.13 | 0.22 | 0.35 | 0.23 | 0.21 | |
| Total | 100 | 100 | 100 | 100 | 100 | 99 | 99 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Lee, S.-Y.; Jang, S.-K.; Kim, K.-J.; Park, M.-J. Anti-Inflammatory Effects of Essential Oils from the Peels of Citrus Cultivars. Pharmaceutics 2023, 15, 1595. https://doi.org/10.3390/pharmaceutics15061595
Yang J, Lee S-Y, Jang S-K, Kim K-J, Park M-J. Anti-Inflammatory Effects of Essential Oils from the Peels of Citrus Cultivars. Pharmaceutics. 2023; 15(6):1595. https://doi.org/10.3390/pharmaceutics15061595
Chicago/Turabian StyleYang, Jiyoon, Su-Yeon Lee, Soo-Kyeong Jang, Ki-Joong Kim, and Mi-Jin Park. 2023. "Anti-Inflammatory Effects of Essential Oils from the Peels of Citrus Cultivars" Pharmaceutics 15, no. 6: 1595. https://doi.org/10.3390/pharmaceutics15061595
APA StyleYang, J., Lee, S. -Y., Jang, S. -K., Kim, K. -J., & Park, M. -J. (2023). Anti-Inflammatory Effects of Essential Oils from the Peels of Citrus Cultivars. Pharmaceutics, 15(6), 1595. https://doi.org/10.3390/pharmaceutics15061595