
Therapeutic Potential and Predictive Pharmaceutical Modeling of Stilbenes in Cannabis sativa





Abstract
:1. Introduction
2. Canniprene
2.1. Anti-Inflammatory Effects
2.2. Anti-Cancer Effects
2.3. Antiviral Effects
3. Cannabistilbene
Anti-Inflammatory Effects
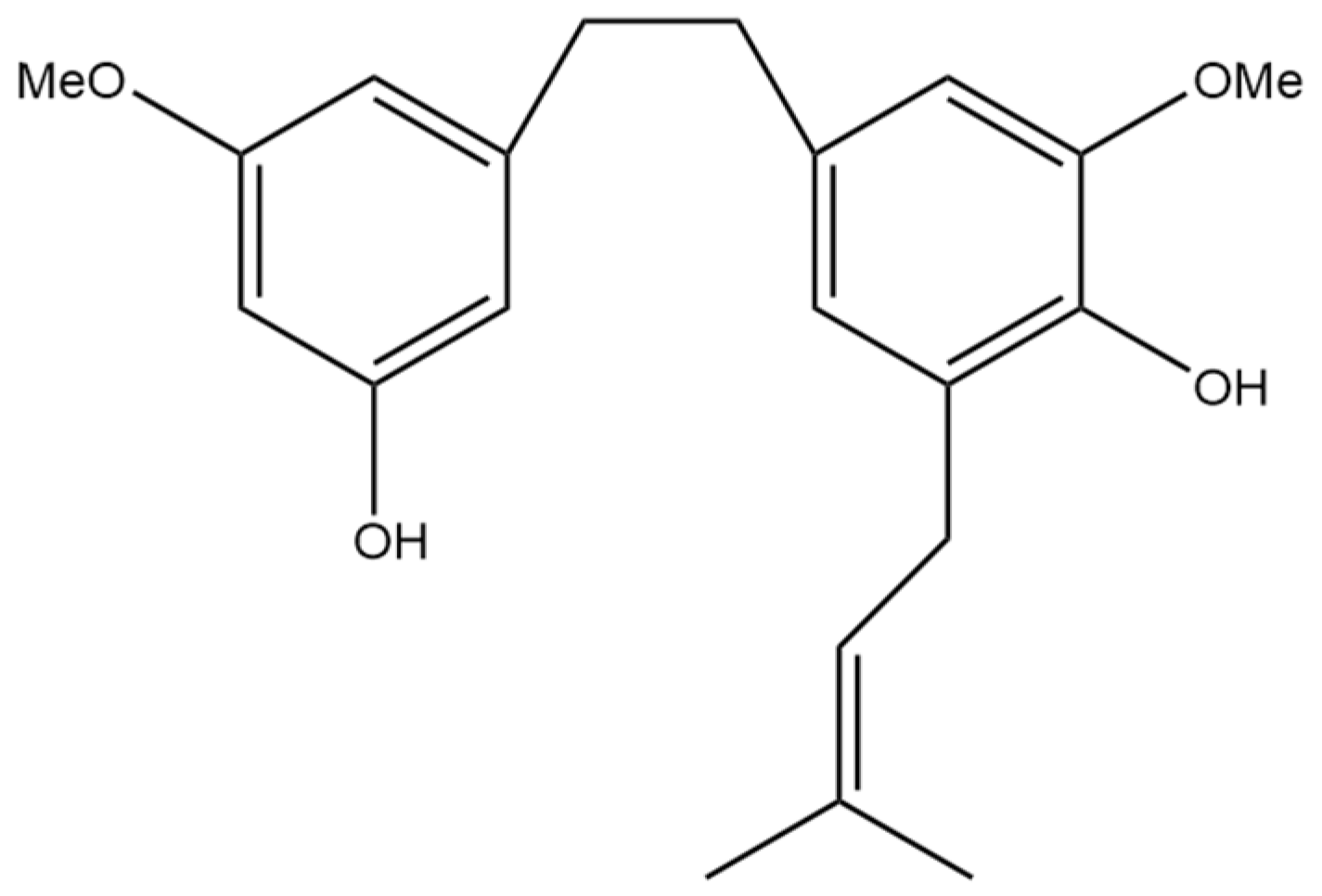
4. 3,4′-Dihydroxy-5,3′-dimethoxy-5′-isoprenylbibenzyl
Anti-Inflammatory Effects
5. HM1, HM2, and HM3
5.1. Anti-Cancer Effects
5.2. Cardioprotective Effects
6. α,α′-Dihydro-3,4′,5-trihydroxy-4,5′-diisopentenylstilbene
6.1. Anti-Cancer Effects
6.2. Antioxidant Effects
7. Dihydroresveratrol
7.1. Anti-Inflammatory Effects
7.2. Anti-Cancer Effects
7.3. Antioxidant Effects
7.4. Anti-Diabetic Effects
8. 3,4′-Dihydroxy-5-methoxy Bibenzyl
8.1. Anti-Inflammatory Effects
8.2. Estrogenic Effects
9. Gigantol
9.1. Anti-Inflammatory Effects
9.2. Anti-Cancer Effects
9.3. Antioxidant Effects
9.4. Spasmolytic Effects
9.5. Androgenic Effects
9.6. Antiparasitic Effects
10. Combretastatin B-2
Anti-Cancer Effects
11. 3-O-Methylbatatasin
12. In Silico Modeling
12.1. Methods
12.2. Results
13. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rock, E.M.; Parker, L.A. Constituents of Cannabis Sativa. In Cannabinoids and Neuropsychiatric Disorders; Murillo-Rodriguez, E., Pandi-Perumal, S.R., Monti, J.M., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2021; pp. 1–13. ISBN 978-3-030-57369-0. [Google Scholar]
- White, C.M. A Review of Human Studies Assessing Cannabidiol’s (CBD) Therapeutic Actions and Potential. J. Clin. Pharmacol. 2019, 59, 923–934. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Rasul Suleria, H.A.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A Comprehensive Review of the Health Perspectives of Resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef] [PubMed]
- Roupe, K.A.; Remsberg, C.M.; Yanez, J.A.; Davies, N.M. Pharmacometrics of Stilbenes: Seguing Towards the Clinic. Curr. Clin. Pharmacol. 2006, 1, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Flores-Sanchez, I.J.; Verpoorte, R. Secondary Metabolism in Cannabis. Phytochem. Rev. 2008, 7, 615–639. [Google Scholar] [CrossRef]
- Crombie, L.; Mary, W.; Crombie, L. Dihydrostilbenes of Thailand Cannabis. Tetrahedron Lett. 1978, 19, 4711–4714. [Google Scholar] [CrossRef]
- Crombie, L.; Crombie, W.M.L.; Jamieson, S.V. Extractives of Thailand Cannabis: Synthesis of Canniprene and Isolation of New Geranylated and Prenylated Chrysoeriols. Tetrahedron Lett. 1980, 21, 3607–3610. [Google Scholar] [CrossRef]
- Elsohly, H.N.; Turner, C.E. Constituents of Cannabis sativa L. XXII: Isolation of Spiro-Indan and Dihydrostilbene Compounds from a Panamanian Variant Grown in Mississippi, United States of America. Bull. Narc. 1982, 34, 51–56. [Google Scholar]
- Allegrone, G.; Pollastro, F.; Magagnini, G.; Taglialatela-Scafati, O.; Seegers, J.; Koeberle, A.; Werz, O.; Appendino, G. The Bibenzyl Canniprene Inhibits the Production of Pro-Inflammatory Eicosanoids and Selectively Accumulates in Some Cannabis sativa Strains. J. Nat. Prod. 2017, 80, 731–734. [Google Scholar] [CrossRef]
- Werz, O.; Seegers, J.; Schaible, A.M.; Weinigel, C.; Barz, D.; Koeberle, A.; Allegrone, G.; Pollastro, F.; Zampieri, L.; Grassi, G.; et al. Cannflavins from Hemp Sprouts, a Novel Cannabinoid-Free Hemp Food Product, Target Microsomal Prostaglandin E2 Synthase-1 and 5-Lipoxygenase. PharmaNutrition 2014, 2, 53–60. [Google Scholar] [CrossRef]
- Rubin, P.; Mollison, K.W. Pharmacotherapy of Diseases Mediated by 5-Lipoxygenase Pathway Eicosanoids. Prostaglandins Other Lipid Mediat. 2007, 83, 188–197. [Google Scholar] [CrossRef]
- Koeberle, A.; Laufer, S.A.; Werz, O. Design and Development of Microsomal Prostaglandin E2 Synthase-1 Inhibitors: Challenges and Future Directions. J. Med. Chem. 2016, 59, 5970–5986. [Google Scholar] [CrossRef]
- Romero Salamanca, J.M.; Castañeda Castellanos, B.A. Modelo de Interacción Molecular de Metabolitos Presentes en la Especie Cannabis satival Con la Enzima Ciclooxigebasa-1 (COX1) y la Enzima Ciclooxigensa-2 (COX2) de Homo Sapines; Universidad de Ciencias Aplicadas Y Ambientales: Bogota, Colombia, 2019. [Google Scholar]
- Warner, T.D.; Mitchell, J.A. Cyclooxygenases: New Forms, New Inhibitors, and Lessons from the Clinic. FASEB J. 2004, 18, 790–804. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.; Liu, Q.; Hou, P.; Li, F.; Guo, S.; Song, W.; Zhang, H.; Liu, X.; Zhang, S.; Zhang, J.; et al. Stilbenoids and Cannabinoids from the Leaves of Cannabis sativa f. Sativa with Potential Reverse Cholesterol Transport Activity. Food Funct. 2018, 9, 6608–6617. [Google Scholar] [CrossRef]
- Khattab, A.R.; Teleb, M. In Silico Discovery of Non-Psychoactive Scaffolds in Cannabis Halting SARS-CoV-2 Host Entry and Replication Machinery. Future Virol. 2022, 17, 367–386. [Google Scholar] [CrossRef] [PubMed]
- ElSohly, H.N.; Ma, G.-E.; Turner, C.E.; ElSohly, M.A. Constituents of Cannabis Sativa, XXV. Isolation of Two New Dihydrostilbenes from a Panamanian Variant. J. Nat. Prod. 1984, 47, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, S.; Akhtar, T.; Casaretto, J.; Perrin, C.; Gelder, K.V. Polypeptides for Use in the Synthesis of Bioactive Phenolic Compounds. Available online: https://patents.google.com/patent/WO2022109736A1/en (accessed on 3 November 2022).
- Boddington, K.F.; Soubeyrand, E.; Van Gelder, K.; Casaretto, J.A.; Perrin, C.; Forrester, T.J.B.; Parry, C.; Al-Abdul-Wahid, M.S.; Jentsch, N.G.; Magolan, J.; et al. Bibenzyl Synthesis in Cannabis sativa L. Plant J. 2022, 109, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Pollastro, F.; Minassi, A.; Fresu, L.G. Cannabis Phenolics and Their Bioactivities. Curr. Med. Chem. 2018, 25, 1160–1185. [Google Scholar] [CrossRef]
- Kettenes-van den Bosch, J.J.; Salemink, C.A. Cannabis XIX. Oxygenated 1,2-Diphenylethanes from Marihuana. Recl. Trav. Chim. Pays-Bas 1978, 97, 221–222. [Google Scholar] [CrossRef]
- Zhao, G.-J.; Yin, K.; Fu, Y.; Tang, C.-K. The Interaction of ApoA-I and ABCA1 Triggers Signal Transduction Pathways to Mediate Efflux of Cellular Lipids. Mol. Med. 2011, 18, 149–158. [Google Scholar] [CrossRef]
- Vergeer, M.; Holleboom, A.G.; Kastelein, J.J.P.; Kuivenhoven, J.A. The HDL Hypothesis: Does High-Density Lipoprotein Protect from Atherosclerosis? J. Lipid Res. 2010, 51, 2058–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biondi, D.M.; Rocco, C.; Ruberto, G. New Dihydrostilbene Derivatives from the Leaves of Glycyrrhiza Glabra and Evaluation of Their Antioxidant Activity. J. Nat. Prod. 2003, 66, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.-S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.; Kulling, S.E. In Vivo and in Vitro Metabolism of Trans-Resveratrol by Human Gut Microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef] [Green Version]
- El-Feraly, F.S. Isolation, Characterization, and Synthesis of 3,5,4′-Trihydroxybibenzyl from Cannabis Sativa. J. Nat. Prod. 1984, 47, 89–92. [Google Scholar] [CrossRef]
- Juan, M.E.; Alfaras, I.; Planas, J.M. Determination of Dihydroresveratrol in Rat Plasma by HPLC. J. Agric. Food Chem. 2010, 58, 7472–7475. [Google Scholar] [CrossRef]
- Radko, Y.; Christensen, K.B.; Christensen, L.P. Semi-Preparative Isolation of Dihydroresveratrol-3-O-β-d-Glucuronide and Four Resveratrol Conjugates from Human Urine after Oral Intake of a Resveratrol-Containing Dietary Supplement. J. Chromatogr. B 2013, 930, 54–61. [Google Scholar] [CrossRef]
- Poór, M.; Kaci, H.; Bodnárová, S.; Mohos, V.; Fliszár-Nyúl, E.; Kunsági-Máté, S.; Özvegy-Laczka, C.; Lemli, B. Interactions of Resveratrol and Its Metabolites (Resveratrol-3-Sulfate, Resveratrol-3-Glucuronide, and Dihydroresveratrol) with Serum Albumin, Cytochrome P450 Enzymes, and OATP Transporters. Biomed. Pharmacother. 2022, 151, 113136. [Google Scholar] [CrossRef]
- Ji, Q.-G.; Ma, M.-H.; Hu, X.-M.; Zhang, Y.-J.; Xu, X.-H.; Nian, H. Detection and Structural Characterization of the Metabolites of Dihydroresveratrol in Rats by Liquid Chromatography Coupled to High-Resolution Tandem Mass Spectrometry. Rapid Commun. Mass Spectrom. 2021, 35, e8991. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.; Liu, X.; Zhao, C.; Yin, J.; Li, X.; Zhang, X.; Wang, J.; Wang, S. Distinctive Anti-Inflammatory Effects of Resveratrol, Dihydroresveratrol, and 3-(4-Hydroxyphenyl)-Propionic Acid on DSS-Induced Colitis in Pseudo-Germ-Free Mice. Food Chem. 2023, 400, 133904. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The Role of Interleukin-1 in General Pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef] [Green Version]
- Fei, Y.; Zhang, S.; Han, S.; Qiu, B.; Lu, Y.; Huang, W.; Li, F.; Chen, D.; Berglund, B.; Xiao, H.; et al. The Role of Dihydroresveratrol in Enhancing the Synergistic Effect of Ligilactobacillus Salivarius Li01 and Resveratrol in Ameliorating Colitis in Mice. Research 2022, 2022, 9863845. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Han, Y.; Wu, X.; Cao, X.; Gao, Z.; Sun, Y.; Wang, M.; Xiao, H. Gut Microbiota-Derived Resveratrol Metabolites, Dihydroresveratrol and Lunularin, Significantly Contribute to the Biological Activities of Resveratrol. Front. Nutr. 2022, 9, 912591. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Gálvez, M.Á.; Espín, J.C.; González-Sarrías, A. Physiological Relevance of the Antiproliferative and Estrogenic Effects of Dietary Polyphenol Aglycones versus Their Phase-II Metabolites on Breast Cancer Cells: A Call of Caution. J. Agric. Food Chem. 2018, 66, 8547–8555. [Google Scholar] [CrossRef] [PubMed]
- Gakh, A.A.; Anisimova, N.Y.; Kiselevsky, M.V.; Sadovnikov, S.V.; Stankov, I.N.; Yudin, M.V.; Rufanov, K.A.; Krasavin, M.Y.; Sosnov, A.V. Dihydro-Resveratrol—A Potent Dietary Polyphenol. Bioorg. Med. Chem. Lett. 2010, 20, 6149–6151. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Bastida, J.A.; Ávila-Gálvez, M.Á.; Espín, J.C.; González-Sarrías, A. Conjugated Physiological Resveratrol Metabolites Induce Senescence in Breast Cancer Cells: Role of P53/P21 and P16/Rb Pathways, and ABC Transporters. Mol. Nutr. Food Res. 2019, 63, 1900629. [Google Scholar] [CrossRef] [PubMed]
- Faragher, R.G.A.; Burton, D.G.A.; Majecha, P.; Fong, N.S.Y.; Davis, T.; Sheerin, A.; Ostler, E.L. Resveratrol, but Not Dihydroresveratrol, Induces Premature Senescence in Primary Human Fibroblasts. Age 2011, 33, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, N.Y.; Kiselevsky, M.V.; Sosnov, A.V.; Sadovnikov, S.V.; Stankov, I.N.; Gakh, A.A. Trans-, Cis-, and Dihydro-Resveratrol: A Comparative Study. Chem. Cent. J. 2011, 5, 88. [Google Scholar] [CrossRef] [Green Version]
- Tsang, S.W.; Guan, Y.-F.; Wang, J.; Bian, Z.-X.; Zhang, H.-J. Inhibition of Pancreatic Oxidative Damage by Stilbene Derivative Dihydro-Resveratrol: Implication for Treatment of Acute Pancreatitis. Sci. Rep. 2016, 6, 22859. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Cai, X.; Zeng, Q.; Ren, X.; Kong, Q. Exploring the Mechanism of Stilbenes to Quench Singlet Oxygen Based on the Key Structures of Resveratrol and Its Analogues. Food Chem. 2023, 403, 134350. [Google Scholar] [CrossRef]
- Lin, Z.-S.; Ku, C.F.; Guan, Y.-F.; Xiao, H.-T.; Shi, X.-K.; Wang, H.-Q.; Bian, Z.-X.; Tsang, S.W.; Zhang, H.-J. Dihydro-Resveratrol Ameliorates Lung Injury in Rats with Cerulein-Induced Acute Pancreatitis. Phytother. Res. 2016, 30, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Calvo, R.; Girona, J.; Rodríguez, M.; Samino, S.; Barroso, E.; de Gonzalo-Calvo, D.; Guaita-Esteruelas, S.; Heras, M.; van der Meer, R.W.; Lamb, H.J.; et al. Fatty Acid Binding Protein 4 (FABP4) as a Potential Biomarker Reflecting Myocardial Lipid Storage in Type 2 Diabetes. Metabolism 2019, 96, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; González-Sarrías, A.; Larrosa, M.; Vallejo, F.; Pallarés, F.J.; Lucas, R.; Morales, J.C.; Tomás-Barberán, F.A.; García-Conesa, M.-T.; et al. Effects of Long-Term Consumption of Low Doses of Resveratrol on Diet-Induced Mild Hypercholesterolemia in Pigs: A Transcriptomic Approach to Disease Prevention. J. Nutr. Biochem. 2012, 23, 829–837. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.; Liu, X.; Yin, J.; Li, X.; Zhang, X.; Xing, X.; Wang, J.; Wang, S. Differential Protective Effect of Resveratrol and Its Microbial Metabolites on Intestinal Barrier Dysfunction Is Mediated by the AMPK Pathway. J. Agric. Food Chem. 2022, 70, 11301–11313. [Google Scholar] [CrossRef]
- Sorini, C.; Cosorich, I.; Lo Conte, M.; De Giorgi, L.; Facciotti, F.; Lucianò, R.; Rocchi, M.; Ferrarese, R.; Sanvito, F.; Canducci, F.; et al. Loss of Gut Barrier Integrity Triggers Activation of Islet-Reactive T Cells and Autoimmune Diabetes. Proc. Natl. Acad. Sci. USA 2019, 116, 15140–15149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Huang, Y.; Sun, J.; Zhang, T.; Liu, T.; Ke, B.; Shi, X.; Li, H.; Zhang, G.; Ye, Z.; et al. Protective Effects of Calorie Restriction on Insulin Resistance and Islet Function in STZ-Induced Type 2 Diabetes Rats. Nutr. Metab. 2021, 18, 48. [Google Scholar] [CrossRef]
- Pallauf, K.; Chin, D.; Günther, I.; Birringer, M.; Lüersen, K.; Schultheiß, G.; Vieten, S.; Krauß, J.; Bracher, F.; Danylec, N.; et al. Resveratrol, Lunularin and Dihydroresveratrol Do Not Act as Caloric Restriction Mimetics When Administered Intraperitoneally in Mice. Sci. Rep. 2019, 9, 4445. [Google Scholar] [CrossRef] [Green Version]
- Günther, I.; Rimbach, G.; Mack, C.I.; Weinert, C.H.; Danylec, N.; Lüersen, K.; Birringer, M.; Bracher, F.; Soukup, S.T.; Kulling, S.E.; et al. The Putative Caloric Restriction Mimetic Resveratrol Has Moderate Impact on Insulin Sensitivity, Body Composition, and the Metabolome in Mice. Mol. Nutr. Food Res. 2020, 64, 1901116. [Google Scholar] [CrossRef]
- Richard, J.; Lingvay, I. Hepatic Steatosis and Type 2 Diabetes: Current and Future Treatment Considerations. Expert Rev. Cardiovasc. Ther. 2011, 9, 321–328. [Google Scholar] [CrossRef]
- Trepiana, J.; Krisa, S.; Renouf, E.; Portillo, M.P. Resveratrol Metabolites Are Able to Reduce Steatosis in Cultured Hepatocytes. Pharmaceuticals 2020, 13, 285. [Google Scholar] [CrossRef]
- Timmers, S.; de Ligt, M.; Phielix, E.; van de Weijer, T.; Hansen, J.; Moonen-Kornips, E.; Schaart, G.; Kunz, I.; Hesselink, M.K.C.; Schrauwen-Hinderling, V.B.; et al. Resveratrol as Add-on Therapy in Subjects with Well-Controlled Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Care 2016, 39, 2211–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.-Q.; Jiang, H.; Li, Q.-M.; Zha, X.-Q.; Luo, J.-P. [Study on chemical constituents of Dendrobium huoshanense stems and their anti-inflammatory activity]. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Medica 2020, 45, 3452–3458. [Google Scholar] [CrossRef]
- Wirth, P.W.; Murphy, J.C.; El-Feraly, F.S.; Turner, C.E. Constituents of Cannabis sativa L. XXI: Estrogenic Activity of a Non-Cannabinoid Constituent. Experientia 1981, 37, 1181–1182. [Google Scholar] [CrossRef] [PubMed]
- Teoh, E.S. Medicinal Orchids of Asia; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24272-9. [Google Scholar]
- Charoenrungruang, S.; Chanvorachote, P.; Sritularak, B.; Pongrakhananon, V. Gigantol, a Bibenzyl from Dendrobium Draconis, Inhibits the Migratory Behavior of Non-Small Cell Lung Cancer Cells. J. Nat. Prod. 2014, 77, 1359–1366. [Google Scholar] [CrossRef]
- Déciga-Campos, M.; Palacios-Espinosa, J.F.; Reyes-Ramírez, A.; Mata, R. Antinociceptive and Anti-Inflammatory Effects of Compounds Isolated from Scaphyglottis Livida and Maxillaria Densa. J. Ethnopharmacol. 2007, 114, 161–168. [Google Scholar] [CrossRef]
- Won, J.-H.; Kim, J.-Y.; Yun, K.-J.; Lee, J.-H.; Back, N.-I.; Chung, H.-G.; Chung, S.A.; Jeong, T.-S.; Choi, M.-S.; Lee, K.-T. Gigantol Isolated from the Whole Plants of Cymbidium Goeringii Inhibits the LPS-Induced INOS and COX-2 Expression via NF-ΚB Inactivation in RAW 264.7 Macrophages Cells. Planta Med. 2006, 72, 1181–1187. [Google Scholar] [CrossRef]
- Simmler, C.; Antheaume, C.; Lobstein, A. Antioxidant Biomarkers from Vanda Coerulea Stems Reduce Irradiated HaCaT PGE-2 Production as a Result of COX-2 Inhibition. PLoS ONE 2010, 5, e13713. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Romero, Y.; Rojas, J.-I.; Castillo, R.; Rojas, A.; Mata, R. Spasmolytic Effects, Mode of Action, and Structure−Activity Relationships of Stilbenoids from Nidema boothii. J. Nat. Prod. 2004, 67, 160–167. [Google Scholar] [CrossRef]
- Huang, J.; Liu, C.; Duan, S.; Lin, J.; Luo, Y.; Tao, S.; Xing, S.; Zhang, X.; Du, H.; Wang, H.; et al. Gigantol Inhibits Proliferation and Enhances DDP-Induced Apoptosis in Breast-Cancer Cells by Downregulating the PI3K/Akt/MTOR Signaling Pathway. Life Sci. 2021, 274, 119354. [Google Scholar] [CrossRef]
- Bhummaphan, N.; Chanvorachote, P. Gigantol Suppresses Cancer Stem Cell-Like Phenotypes in Lung Cancer Cells. Evid. Based Complement. Altern. Med. 2015, 2015, e836564. [Google Scholar] [CrossRef] [Green Version]
- Losuwannarak, N.; Maiuthed, A.; Kitkumthorn, N.; Leelahavanichkul, A.; Roytrakul, S.; Chanvorachote, P. Gigantol Targets Cancer Stem Cells and Destabilizes Tumors via the Suppression of the PI3K/AKT and JAK/STAT Pathways in Ectopic Lung Cancer Xenografts. Cancers 2019, 11, 2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losuwannarak, N.; Roytrakul, S.; Chanvorachote, P. Gigantol Targets MYC for Ubiquitin-Proteasomal Degradation and Suppresses Lung Cancer Cell Growth. Cancer Genom. Proteom. 2020, 17, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Aksorn, N.; Losuwannarak, N.; Tungsukruthai, S.; Roytrakul, S.; Chanvorachote, P. Analysis of the Protein–Protein Interaction Network Identifying c-Met as a Target of Gigantol in the Suppression of Lung Cancer Metastasis. Cancer Genom. Proteom. 2021, 18, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, H.; Yin, D.; Xue, X.; Chen, X.; Li, X.; Li, J.; Yi, Y. Effect of Gigantol on the Proliferation of Hepatocellular Carcinoma Cells Tested by a Network-Based Pharmacological Approach and Experiments. Front. Biosci.-Landmark 2022, 27, 25. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Sun, Y.; Hu, X.; Liu, L. Gigantol Inhibits Proliferation and Enhanced Oxidative Stress-Mediated Apoptosis through Modulating of Wnt/β-Catenin Signaling Pathway in HeLa Cells. J. Biochem. Mol. Toxicol. 2022, 36, e22944. [Google Scholar] [CrossRef]
- Ahammed, S.; Afrin, R.; Uddin, N.; Al-Amin, Y.; Hasan, K.; Haque, U.; Islam, K.M.M.; Alam, A.H.M.K.; Tanaka, T.; Sadik, G. Acetylcholinesterase Inhibitory and Antioxidant Activity of the Compounds Isolated from Vanda Roxburghii. Adv. Pharmacol. Pharm. Sci. 2021, 2021, 5569054. [Google Scholar] [CrossRef]
- Annaházi, A.; Róka, R.; Rosztóczy, A.; Wittmann, T. Role of Antispasmodics in the Treatment of Irritable Bowel Syndrome. World J. Gastroenterol. WJG 2014, 20, 6031–6043. [Google Scholar] [CrossRef]
- Basque, A.; Nguyen, H.T.; Touaibia, M.; Martin, L.J. Gigantol Improves Cholesterol Metabolism and Progesterone Biosynthesis in MA-10 Leydig Cells. Curr. Issues Mol. Biol. 2021, 44, 73–93. [Google Scholar] [CrossRef]
- Rahman, F.; Tabrez, S.; Ali, R.; Akand, S.K.; Zahid, M.; Alaidarous, M.A.; Alsaweed, M.; Alshehri, B.M.; Banawas, S.; Bin Dukhyil, A.A.; et al. Virtual Screening of Natural Compounds for Potential Inhibitors of Sterol C-24 Methyltransferase of Leishmania Donovani to Overcome Leishmaniasis. J. Cell. Biochem. 2021, 122, 1216–1228. [Google Scholar] [CrossRef]
- Pettit, G.R.; Singh, S.B. Isolation, Structure, and Synthesis of Combretastatin A-2, A-3, and B-2. Can. J. Chem. 1987, 65, 2390–2396. [Google Scholar] [CrossRef]
- Fyhrquist, P.; Salih, E.Y.A.; Helenius, S.; Laakso, I.; Julkunen-Tiitto, R. HPLC-DAD and UHPLC/QTOF-MS Analysis of Polyphenols in Extracts of the African Species Combretum padoides, C. zeyheri and C. psidioides Related to Their Antimycobacterial Activity. Antibiotics 2020, 9, 459. [Google Scholar] [CrossRef] [PubMed]
- Marti, G.; Schnee, S.; Andrey, Y.; Simoes-Pires, C.; Carrupt, P.-A.; Wolfender, J.-L.; Gindro, K. Study of Leaf Metabolome Modifications Induced by UV-C Radiations in Representative Vitis, Cissus and Cannabis Species by LC-MS Based Metabolomics and Antioxidant Assays. Molecules 2014, 19, 14004–14021. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Romero, Y.; Acevedo, L.; de Los Ángeles Sánchez, M.; Shier, W.T.; Abbas, H.K.; Mata, R. Phytotoxic Activity of Bibenzyl Derivatives from the Orchid Epidendrum rigidum. J. Agric. Food Chem. 2005, 53, 6276–6280. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Morikawa, T.; Xie, H.; Yoshikawa, M. Antiallergic Phenanthrenes and Stilbenes from the Tubers of Gymnadenia Conopsea. Planta Med. 2004, 70, 847–855. [Google Scholar] [CrossRef]
- Zhang, C.; Ning, D.; Pan, J.; Chen, C.; Gao, C.; Ding, Z.; Jiang, F.; Li, M. Anti-Inflammatory Effect Fraction of Bletilla Striata and Its Protective Effect on LPS-Induced Acute Lung Injury. Mediat. Inflamm. 2021, 2021, 6684120. [Google Scholar] [CrossRef]
- Amaral Silva, D.; Pate, D.W.; Clark, R.D.; Davies, N.M.; El-Kadi, A.O.S.; Löbenberg, R. Phytocannabinoid Drug-Drug Interactions and Their Clinical Implications. Pharmacol. Ther. 2020, 215, 107621. [Google Scholar] [CrossRef]
- Mao, Q.; Unadkat, J.D. Role of the Breast Cancer Resistance Protein (BCRP/ABCG2) in Drug Transport—An Update. AAPS J. 2014, 17, 65–82. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Lei, T.; Wang, Z.; Shen, C.; Cao, D.; Hou, T. ADMET Evaluation in Drug Discovery. 20. Prediction of Breast Cancer Resistance Protein Inhibition through Machine Learning. J. Cheminform. 2020, 12, 16. [Google Scholar] [CrossRef] [Green Version]
- Mbhele, N.; Chimukangara, B.; Gordon, M. HIV-1 Integrase Strand Transfer Inhibitors: A Review of Current Drugs, Recent Advances and Drug Resistance. Int. J. Antimicrob. Agents 2021, 57, 106343. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stilbene | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CYP SUBSTRATE | ||||||||||||||
| CYP1A2 | 91 | 79 | 79 | 79 | 79 | 91 | 49 | 51 | 79 | 91 | 91 | 91 | 91 | 91 |
| CYP2A6 | −69 | −66 | −72 | −67 | 63 | −72 | −86 | −86 | −98 | −98 | 60 | 55 | 44 | 82 |
| CYP2B6 | −89 | −67 | −92 | −89 | −89 | −92 | −89 | −89 | −86 | −92 | −73 | −92 | −89 | 42 |
| CYP2C8 | −86 | −83 | −92 | −92 | −86 | −83 | −80 | −86 | −89 | −80 | −86 | −92 | −89 | −89 |
| CYP2C9 | 39 | 45 | −67 | −66 | 34 | −67 | 37 | 33 | −65 | −85 | 45 | −66 | −66 | 50 |
| CYP2C19 | 82 | 82 | 82 | 82 | 82 | 54 | 62 | 63 | 62 | 40 | 82 | 82 | 82 | 82 |
| CYP2D6 | −54 | 56 | −60 | −60 | 55 | −63 | −61 | −61 | −64 | −95 | −55 | −56 | −57 | 52 |
| CYP2E1 | −71 | −69 | 63 | 63 | −73 | −71 | −87 | −82 | −87 | 40 | 74 | 56 | 49 | 82 |
| CYP3A4 | 73 | −54 | −42 | −38 | 70 | −62 | 72 | −65 | −48 | −84 | −84 | −48 | −51 | −51 |
| CYP INHIBITOR | ||||||||||||||
| CYP1A2 | −63 | −79 | −61 | −57 | −57 | 62 | −61 | −57 | −97 | −97 | −97 | −47 | 66 | −76 |
| CYP2C9 | 53 | 53 | 47 | 43 | 47 | 34 | 53 | 45 | 47 | 36 | 39 | 35 | 34 | 47 |
| CYP2C19 | −98 | 18 | −99 | −98 | −99 | −99 | −99 | −98 | 20 | −84 | −87 | −98 | −99 | 18 |
| CYP2D6 | 51 | 51 | −72 | −76 | 51 | 44 | 70 | 70 | 55 | −95 | −60 | −70 | −84 | −57 |
| CYP3A4 | 59 | −76 | 51 | 46 | 55 | 51 | 80 | 80 | −65 | −81 | −90 | −64 | 51 | −78 |
| Stilbene | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| UGT SUBSTRATE | ||||||||||||||
| UGT1A1 | 63 | 53 | 56 | 56 | 63 | −45 | 56 | −41 | −66 | −50 | 58 | 58 | 63 | 63 |
| UGT1A3 | 90 | 90 | 51 | 51 | 90 | 97 | 97 | 97 | 97 | 83 | 64 | 64 | 73 | −48 |
| UGT1A4 | −99 | −92 | −99 | −99 | −99 | −99 | −99 | −99 | −95 | −99 | −95 | −99 | −99 | −86 |
| UGT1A6 | −97 | −97 | −97 | −97 | −91 | −91 | −97 | −91 | −83 | 48 | −62 | −83 | −85 | −80 |
| UGT1A8 | 75 | 62 | −66 | −63 | 83 | 98 | 67 | 81 | 93 | 55 | −46 | 72 | 83 | −96 |
| UGT1A9 | 97 | 97 | 97 | 97 | 97 | 97 | 76 | 97 | 97 | 97 | 97 | 97 | 97 | 97 |
| UGT1A10 | −65 | −66 | −77 | −83 | −83 | 53 | −61 | −56 | 53 | 50 | −56 | −61 | −70 | −90 |
| UGT2B7 | 68 | −60 | 68 | 72 | 72 | 93 | 85 | 93 | −82 | −55 | −42 | 85 | 93 | −44 |
| UGT2B15 | 49 | 61 | 51 | 51 | 50 | 49 | 46 | 46 | 49 | 50 | 81 | 58 | 51 | 61 |
| Stilbene | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Substrate | 62 | 64 | 79 | 79 | 62 | 79 | −53 |
| Inhibitor | 64 | 67 | −51 | −51 | 62 | 58 | 64 |
| Stilbene | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| Substrate | 60 | 59 | 79 | 75 | 79 | 79 | 70 |
| Inhibitor | 67 | 81 | −81 | −49 | −47 | −44 | 52 |
| Stilbene | Fa (%) | Fb (%) | Cmin (ng/mL) | Cmax (ng/mL) | Tmax (h) | AUC (ng-h/mL) | AUCinf (ng-h/mL) | CL (L/h) | CLp (L/h) | THalf (h) | Vd (L) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 88.6 | 49.13 | 3.42 | 5.28 | 13.5 | 92.27 | 166.45 | 29.52 | 29.52 | 14.9 | 634.34 |
| 2 | 99.55 | 55.34 | 3.24 | 6.79 | 5.51 | 109.95 | 185.3 | 29.86 | 29.16 | 16.55 | 713.22 |
| 3 | 99.99 | 71.23 | 2.1 | 40.61 | 2.24 | 339.19 | 354.11 | 20.11 | 20.1 | 4.95 | 143.68 |
| 4 | 99.99 | 69.1 | 1.05 | 44.69 | 2.12 | 314.15 | 320.06 | 21.59 | 21.58 | 3.94 | 122.58 |
| 5 | 85.07 | 44.91 | 3.1 | 4.49 | 14.71 | 79.34 | 143.54 | 31.29 | 31.29 | 14.27 | 644.03 |
| 6 | 96.57 | 48.67 | 2.6 | 6.06 | 8.97 | 96.61 | 141.31 | 34.44 | 34.45 | 11.82 | 587.55 |
| 7 | 66.53 | 25.88 | 1.62 | 1.69 | 19.72 | 29.8 | 97.06 | 26.66 | 39.83 | 22.32 | 858.52 |
| 8 | 51.18 | 12.25 | 0.68 | 0.69 | 20.49 | 12.33 | 53.76 | 22.78 | 49.5 | 27.32 | 898.04 |
| 9 | 98.08 | 45.23 | 2.31 | 4.18 | 7.91 | 70.32 | 124.22 | 36.41 | 34.25 | 17.33 | 910.28 |
| 10 | 100 | 78.71 | 2.97 | 51.28 | 1.73 | 450.04 | 472.66 | 16.65 | 16.65 | 5.22 | 125.3 |
| 11 | 99.99 | 83.29 | 10.68 | 29.92 | 2.05 | 451.62 | 674.32 | 12.35 | 12.33 | 14.25 | 253.93 |
| 12 | 99.99 | 76.17 | 4.88 | 35.63 | 2.56 | 386.09 | 437.16 | 17.42 | 17.39 | 7.26 | 182.56 |
| 13 | 99.99 | 67.54 | 1.97 | 31.14 | 2.62 | 275.16 | 289.86 | 23.3 | 23.27 | 5.24 | 176.22 |
| 14 | 99.98 | 78.29 | 8.13 | 16.05 | 3.17 | 271.65 | 513.56 | 15.25 | 15.14 | 20.47 | 450.24 |
| Stilbene | Permeability through Human Skin (cm/s × 107) | Effective Human Jejunal Permeability (cm/s × 104) | Blood–Brain Barrier Penetration (Confidence %) | Lipinski’s Rule of 5 Score |
|---|---|---|---|---|
| 1 | 13.784 | 8.011 | High (84%) | 0 |
| 2 | 32.755 | 5.229 | High (81%) | 0 |
| 3 | 2.534 | 7.132 | High (82%) | 0 |
| 4 | 2.385 | 7.172 | High (82%) | 0 |
| 5 | 15.118 | 7.866 | High (82%) | 0 |
| 6 | 16.865 | 7.38 | Low (39%) | 0 |
| 7 | 31.767 | 10.02 | High (70%) | 1 |
| 8 | 47.737 | 8.562 | Low (70%) | 1 |
| 9 | 100.123 | 4.272 | Low (45%) | 1 |
| 10 | 4.824 | 3.951 | Low (48%) | 0 |
| 11 | 4.097 | 5.539 | High (84%) | 0 |
| 12 | 2.807 | 7.246 | High (86%) | 0 |
| 13 | 2.699 | 10.255 | High (89%) | 0 |
| 14 | 10.247 | 7.402 | High (96%) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Croinin, C.; Garcia Guerra, A.; Doschak, M.R.; Löbenberg, R.; Davies, N.M. Therapeutic Potential and Predictive Pharmaceutical Modeling of Stilbenes in Cannabis sativa. Pharmaceutics 2023, 15, 1941. https://doi.org/10.3390/pharmaceutics15071941
O’Croinin C, Garcia Guerra A, Doschak MR, Löbenberg R, Davies NM. Therapeutic Potential and Predictive Pharmaceutical Modeling of Stilbenes in Cannabis sativa. Pharmaceutics. 2023; 15(7):1941. https://doi.org/10.3390/pharmaceutics15071941
Chicago/Turabian StyleO’Croinin, Conor, Andres Garcia Guerra, Michael R. Doschak, Raimar Löbenberg, and Neal M. Davies. 2023. "Therapeutic Potential and Predictive Pharmaceutical Modeling of Stilbenes in Cannabis sativa" Pharmaceutics 15, no. 7: 1941. https://doi.org/10.3390/pharmaceutics15071941
APA StyleO’Croinin, C., Garcia Guerra, A., Doschak, M. R., Löbenberg, R., & Davies, N. M. (2023). Therapeutic Potential and Predictive Pharmaceutical Modeling of Stilbenes in Cannabis sativa. Pharmaceutics, 15(7), 1941. https://doi.org/10.3390/pharmaceutics15071941