

Testing S. sonnei GMMA with and without Aluminium Salt-Based Adjuvants in Animal Models
 ,
,  , , , ,
, , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Generation of Mutants
2.2. GMMA Production and Characterisation
2.3. GMMA Formulation
2.4. Animal Studies
2.5. Ethics and 3R Statement
2.6. Assessment of Anti-S. sonnei LPS and Anti-GMMA Protein Immune Responses in Mice and Rabbits
2.7. Assessment of Serum Bactericidal Activity against Shigella
2.8. Statistical Analysis
3. Results
3.1. GMMA Formulations
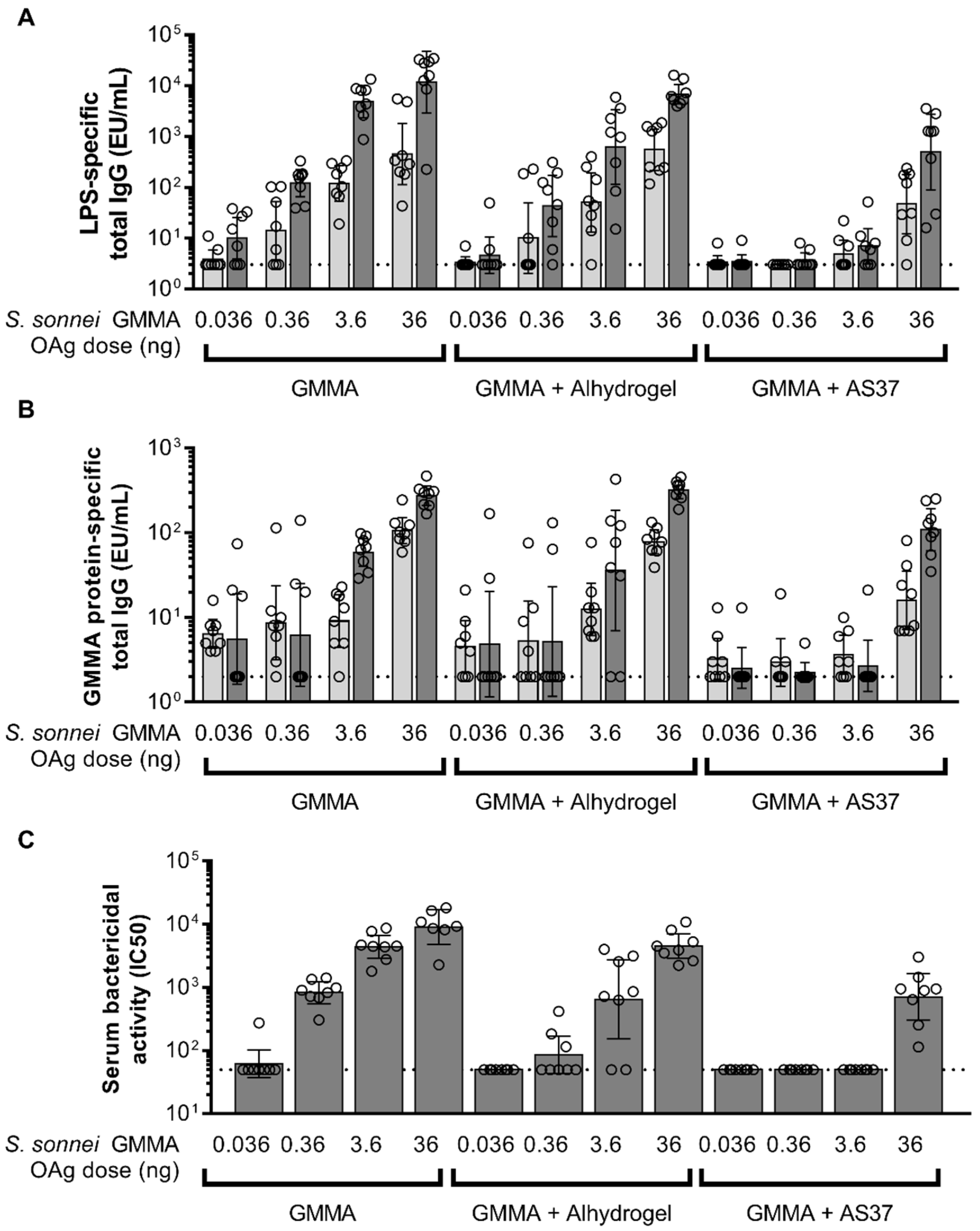
3.2. Comparing GMMA Formulations in Mice
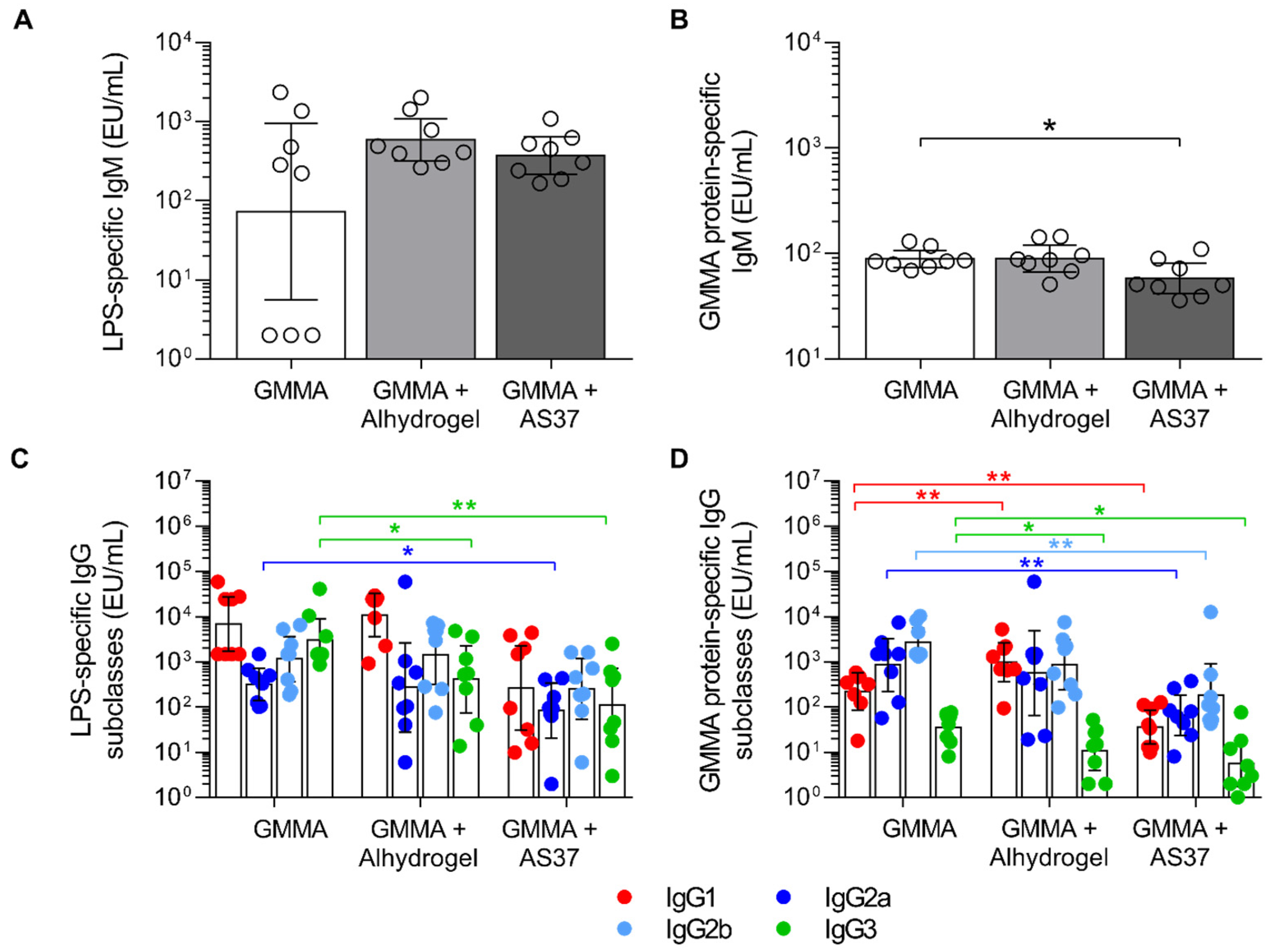
3.3. Comparing GMMA Formulations in Rabbits
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Troeger, C.; Blacker, B.F.; Khalil, I.A.; Rao, P.C.; Cao, S.; Zimsen, S.R.M.; Albertson, S.B.; Stanaway, J.D.; Deshpande, A.; Abebe, Z.; et al. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.; Panchalingham, S.; Sow, S.O.; Sur, D.; Zaidi, A.K.M.; Faruque, A.S.G.; et al. The incidence, aetiology, and adverse clinical consequences of less severe diarrhoeal episodes among infants and children residing in low-income and middle-income countries: A 12-month case-control study as a follow-on to the Global Enteric Multicenter Study (GEMS). Lancet Glob. Health 2019, 7, e568–e584. [Google Scholar] [CrossRef]
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G.; et al. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990–2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar]
- Cohen, D.; Meron-Sudai, S.; Bialik, A.; Asato, V.; Goren, S.; Ariel-Cohen, O.; Reizis, A.; Hochberg, A.; Ashkenazi, S. Serum IgG antibodies to Shigella lipopolysaccharide antigens—A correlate of protection against shigellosis. Hum. Vaccines Immunother. 2019, 15, 1401–1408. [Google Scholar] [CrossRef]
- Cohen, D.; Block, C.; Green, M.S.; Lowell, G.; Ofek, I. Immunoglobulin M, A, and G antibody response to lipopolysaccharide O antigen in symptomatic and asymptomatic Shigella infections. J. Clin. Microbiol. 1989, 27, 162–167. [Google Scholar] [CrossRef]
- Simon, J.K.; Maciel, M.; Weld, E.D.; Wahid, R.; Pasetti, M.F.; Picking, W.L.; Kotloff, K.L.; Levine, M.M.; Sztein, M.B. Antigen-specific IgA B memory cell responses to Shigella antigens elicited in volunteers immunized with live attenuated Shigella flexneri 2a oral vaccine candidates. Clin. Immunol. 2011, 139, 185–192. [Google Scholar] [CrossRef]
- Ndungo, E.; Randall, A.; Hazen, T.H.; Kania, D.A.; Trappl-Kimmons, K.; Liang, X.; Barry, E.M.; Kotloff, K.L.; Chakraborty, S.; Mani, S.; et al. A Novel Shigella Proteome Microarray Discriminates Targets of Human Antibody Reactivity following Oral Vaccination and Experimental Challenge. mSphere 2018, 3. [Google Scholar] [CrossRef]
- Walker, R.; Kaminski, R.W.; Porter, C.; Choy, R.K.M.; White, J.A.; Fleckenstein, J.M.; Cassels, F.; Bourgeois, L. Vaccines for Protecting Infants from Bacterial Causes of Diarrheal Disease. Microorganisms 2021, 9, 1382. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Alaimo, C. The Ongoing Journey of a Shigella Bioconjugate Vaccine. Vaccines 2022, 10, 212. [Google Scholar] [CrossRef]
- Phalipon, A.; Mulard, L.A. Toward a Multivalent Synthetic Oligosaccharide-Based Conjugate Vaccine against Shigella: State-of-the-Art for a Monovalent Prototype and Challenges. Vaccines 2022, 10, 403. [Google Scholar] [CrossRef]
- Turbyfill, K.R.; Clarkson, K.A.; Oaks, E.V.; Kaminski, R.W. From Concept to Clinical Product: A Brief History of the Novel Shigella Invaplex Vaccine’s Refinement and Evolution. Vaccines 2022, 10, 548. [Google Scholar] [CrossRef] [PubMed]
- Rossi, O.; Citiulo, F.; Giannelli, C.; Cappelletti, E.; Gasperini, G.; Mancini, F.; Acquaviva, A.; Raso, M.M.; Sollai, L.; Alfini, R.; et al. A next-generation GMMA-based vaccine candidate to fight shigellosis. NPJ Vaccines 2023, 8, 130. [Google Scholar] [CrossRef]
- Mancini, F.; Rossi, O.; Necchi, F.; Micoli, F. OMV Vaccines and the Role of TLR Agonists in Immune Response. Int. J. Mol. Sci. 2020, 21, 4416. [Google Scholar] [CrossRef] [PubMed]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2019, 14, e1800376. [Google Scholar] [CrossRef] [PubMed]
- Launay, O.; Lewis, D.J.M.; Anemona, A.; Loulergue, P.; Leahy, J.; Sciré, A.S.; Maugard, A.; Marchetti, E.; Zancan, S.; Huo, Z.; et al. Safety Profile and Immunologic Responses of a Novel Vaccine Against Shigella sonnei Administered Intramuscularly, Intradermally and Intranasally: Results From Two Parallel Randomized Phase 1 Clinical Studies in Healthy Adult Volunteers in Europe. EBioMedicine 2017, 22, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Obiero, C.W.; Ndiaye, A.G.W.; Sciré, A.S.; Kaunyangi, B.M.; Marchetti, E.; Gone, A.M.; Schütte, L.D.; Riccucci, D.; Auerbach, J.; Saul, A.; et al. A Phase 2a Randomized Study to Evaluate the Safety and Immunogenicity of the 1790GAHB Generalized Modules for Membrane Antigen Vaccine against Shigella sonnei Administered Intramuscularly to Adults from a Shigellosis-Endemic Country. Front. Immunol. 2017, 8, 1884. [Google Scholar] [CrossRef] [PubMed]
- Launay, O.; Ndiaye, A.G.W.; Conti, V.; Loulergue, P.; Sciré, A.S.; Landre, A.M.; Ferruzzi, P.; Nedjaai, N.; Schütte, L.D.; Auerbach, J.; et al. Booster Vaccination With GVGH Shigella sonnei 1790GAHB GMMA Vaccine Compared to Single Vaccination in Unvaccinated Healthy European Adults: Results From a Phase 1 Clinical Trial. Front. Immunol. 2019, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Kool, M.; Fierens, K.; Lambrecht, B.N. Alum adjuvant: Some of the tricks of the oldest adjuvant. J. Med. Biol. 2012, 61, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Rost, B.E.; Relyveld, E.; Siber, G.R. Adjuvant properties of aluminum and calcium compounds. Pharm. Biotechnol. 1995, 6, 229–248. [Google Scholar] [CrossRef] [PubMed]
- Warren, H.S.; Chedid, L.A. Future prospects for vaccine adjuvants. Crit. Rev. Immunol. 1988, 8, 83–101. [Google Scholar]
- Piccioli, D.; Alfini, R.; Monaci, V.; Arato, V.; Carducci, M.; Aruta, M.G.; Rossi, O.; Necchi, F.; Anemona, A.; Bartolini, E.; et al. Antigen presentation by Follicular Dendritic cells to cognate B cells is pivotal for Generalised Modules for membrane antigens (GMMA) immunogenicity. Vaccine 2022, 40, 6305–6314. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.Y.; Singh, M.; Miller, A.T.; De Gregorio, E.; Doro, F.; D’Oro, U.; Skibinski, D.A.; Mbow, M.L.; Bufali, S.; Herman, A.E.; et al. Rational design of small molecules as vaccine adjuvants. Sci. Transl. Med. 2014, 6, 263ra160. [Google Scholar] [CrossRef]
- Kasturi, S.P.; Skountzou, I.; Albrecht, R.A.; Koutsonanos, D.; Hua, T.; Nakaya, H.I.; Ravindran, R.; Stewart, S.; Alam, M.; Kwissa, M.; et al. Programming the magnitude and persistence of antibody responses with innate immunity. Nature 2011, 470, 543–547. [Google Scholar] [CrossRef]
- Short, K.K.; Miller, S.M.; Walsh, L.; Cybulski, V.; Bazin, H.; Evans, J.T.; Burkhart, D. Co-encapsulation of synthetic lipidated TLR4 and TLR7/8 agonists in the liposomal bilayer results in a rapid, synergistic en-hancement of vaccine-mediated humoral immunity. J. Control. Release 2019, 315, 186–196. [Google Scholar] [CrossRef]
- Napolitani, G.; Rinaldi, A.; Bertoni, F.; Sallusto, F.; Lanzavecchia, A. Selected Toll-like receptor agonist combinations synergistically trigger a T helper type 1-polarizing program in dendritic cells. Nat. Immunol. 2005, 6, 769. [Google Scholar] [CrossRef]
- Surendran, N.; Simmons, A.; Pichichero, M.E. TLR agonist combinations that stimulate Th type I polarizing responses from human neonates. Innate Immun. 2018, 24, 240–251. [Google Scholar] [CrossRef]
- Micoli, F.; Alfini, R.; Giannelli, C. Methods for Assessment of OMV/GMMA Quality and Stability. Methods Mol. Biol. 2022, 2414, 227–279. [Google Scholar] [CrossRef]
- Buonsanti, C.; Balocchi, C.; Harfouche, C.; Corrente, F.; Galli Stampino, L.; Mancini, F.; Tontini, M.; Malyala, P.; Bufali, S.; Baudner, B.; et al. Novel adjuvant Alum-TLR7 significantly potentiates immune response to glycoconjugate vaccines. Sci. Rep. 2016, 6, 29063. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Gasperini, G.; Rossi, O.; Aruta, M.G.; Raso, M.M.; Alfini, R.; Biagini, M.; Necchi, F.; Micoli, F. Dissecting the contribution of O-Antigen and proteins to the immunogenicity of Shigella sonnei generalized modules for membrane antigens (GMMA). Sci. Rep. 2021, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Boland, G.; Beran, J.; Lievens, M.; Sasadeusz, J.; Dentico, P.; Nothdurft, H.; Zuckerman, J.N.; Genton, B.; Steffen, R.; Loutan, L.; et al. Safety and immunogenicity profile of an experimental hepatitis B vaccine adjuvanted with AS04. Vaccine 2004, 23, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Wille-Reece, U.; Flynn, B.J.; Loré, K.; Koup, R.A.; Kedl, R.M.; Mattapallil, J.J.; Weiss, W.R.; Roederer, M.; Seder, R.A. HIV Gag protein conjugated to a Toll-like receptor 7/8 agonist improves the magnitude and quality of Th1 and CD8+ T cell responses in nonhuman primates. Proc. Natl. Acad. Sci. USA 2005, 102, 15190–15194. [Google Scholar] [CrossRef]
- Kim, O.Y.; Hong, B.S.; Park, K.S.; Yoon, Y.J.; Choi, S.J.; Lee, W.H.; Roh, T.Y.; Lötvall, J.; Kim, Y.K.; Gho, Y.S. Immunization with Escherichia coli outer membrane vesicles protects bacteria-induced lethality via Th1 and Th17 cell responses. . J. Immunol. 2013, 190, 4092–4102. [Google Scholar] [CrossRef]
- Lee, B.-J.; Kwon, H.-i.; Kim, E.-H.; Park, S.-J.; Lee, S.-H.; Choi, Y.K.; Kim, S.-H. Assessment of mOMV adjuvant efficacy in the pathogenic H1N1 influenza virus vaccine. Clin. Exp. Vaccine Re.s 2014, 3, 194–201. [Google Scholar] [CrossRef]
- Piccioli, D.; Buricchi, F.; Bacconi, M.; Bechi, N.; Galli, B.; Ferlicca, F.; Luzzi, E.; Cartocci, E.; Marchi, S.; Romagnoli, G.; et al. Enhanced Systemic Humoral Immune Response Induced in Mice by Generalized Modules for Membrane Antigens (GMMA) Is Associated with Affinity Maturation and Isotype Switching. Vaccines 2023, 11, 1219. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Rossi, O.; Conti, V.; Launay, O.; Sciré, A.S.; Aruta, M.G.; Nakakana, U.N.; Marchetti, E.; Rappuoli, R.; Saul, A.; et al. Antibodies Elicited by the Shigella sonnei GMMA Vaccine in Adults Trigger Complement-Mediated Serum Bactericidal Activity: Results From a Phase 1 Dose Escalation Trial Followed by a Booster Extension. Front. Immunol. 2021, 12, 671325. [Google Scholar] [CrossRef] [PubMed]
- Kaaijk, P.; van der Ark, A.A.; van Amerongen, G.; van den Dobbelsteen, G.P. Nonclinical vaccine safety evaluation: Advantages of continuous temperature monitoring using abdominally implanted data loggers. J. Appl. Toxicol. 2013, 33, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.; Myhill, S.; Palmieri, E.; Necchi, F.; Rijpkema, S.; Micoli, F.; Nordgren, I.K.; Rossi, O.; Vipond, C. Development of a Monocyte Activation Test as an Alternative to the Rabbit Pyrogen Test for Mono- and Multi-Component Shigella GMMA-Based Vaccines. Microorganisms 2021, 9, 1375. [Google Scholar] [CrossRef] [PubMed]
- Rossi, O.; Citiulo, F.; Mancini, F. Outer membrane vesicles: Moving within the intricate labyrinth of assays that can predict risks of reactogenicity in humans. Hum. Vaccines Immunother. 2021, 17, 601–613. [Google Scholar] [CrossRef]
- Keiser, P.B.; Gibbs, B.T.; Coster, T.S.; Moran, E.E.; Stoddard, M.B.; Labrie, J.E., 3rd; Schmiel, D.H.; Pinto, V.; Chen, P.; Zollinger, W.D. A phase 1 study of a group B meningococcal native outer membrane vesicle vaccine made from a strain with deleted lpxL2 and synX and stable expression of opcA. Vaccine 2010, 28, 6970–6976. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pearson Correlation Coefficients vs. Bactericidal Activity (and p-Values of the Correlation) | |
|---|---|
| IgG | 0.923 (0.000) |
| IgM | −0.363 (0.089) |
| IgG2a | 0.532 (0.009) |
| IgG1 | 0.629 (0.001) |
| IgG2b | 0.505 (0.014) |
| IgG3 | 0.714 (0.000) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancini, F.; Caradonna, V.; Alfini, R.; Aruta, M.G.; Vitali, C.G.; Gasperini, G.; Piccioli, D.; Berlanda Scorza, F.; Rossi, O.; Micoli, F. Testing S. sonnei GMMA with and without Aluminium Salt-Based Adjuvants in Animal Models. Pharmaceutics 2024, 16, 568. https://doi.org/10.3390/pharmaceutics16040568
Mancini F, Caradonna V, Alfini R, Aruta MG, Vitali CG, Gasperini G, Piccioli D, Berlanda Scorza F, Rossi O, Micoli F. Testing S. sonnei GMMA with and without Aluminium Salt-Based Adjuvants in Animal Models. Pharmaceutics. 2024; 16(4):568. https://doi.org/10.3390/pharmaceutics16040568
Chicago/Turabian StyleMancini, Francesca, Valentina Caradonna, Renzo Alfini, Maria Grazia Aruta, Claudia Giorgina Vitali, Gianmarco Gasperini, Diego Piccioli, Francesco Berlanda Scorza, Omar Rossi, and Francesca Micoli. 2024. "Testing S. sonnei GMMA with and without Aluminium Salt-Based Adjuvants in Animal Models" Pharmaceutics 16, no. 4: 568. https://doi.org/10.3390/pharmaceutics16040568
APA StyleMancini, F., Caradonna, V., Alfini, R., Aruta, M. G., Vitali, C. G., Gasperini, G., Piccioli, D., Berlanda Scorza, F., Rossi, O., & Micoli, F. (2024). Testing S. sonnei GMMA with and without Aluminium Salt-Based Adjuvants in Animal Models. Pharmaceutics, 16(4), 568. https://doi.org/10.3390/pharmaceutics16040568