
Sleep and Chronobiology as a Key to Understand Cluster Headache


 , , ,
, , ,  ,
, 
{kind=link}
Abstract
:1. Introduction
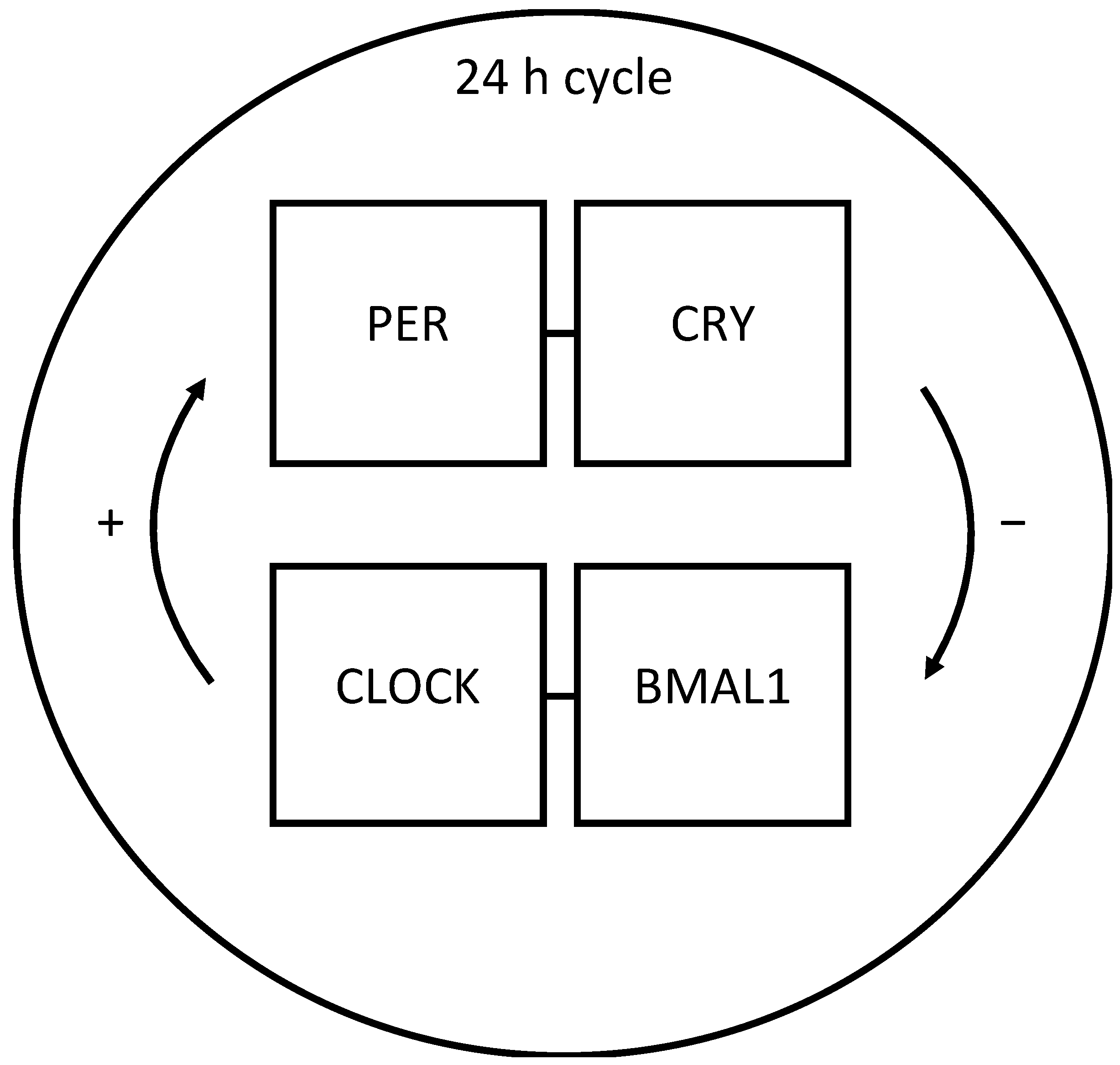
2. Chronobiology in Cluster Headache
2.1. Diurnal Rhythmicity
2.2. Seasonality
2.3. Molecular Mechanisms
3. Sleep Disorders and CH
3.1. REM Sleep
3.2. Chronotypes and Sleep Quality
3.3. Sleep Breathing Disorders
3.4. The Role of Hypothalamus
4. Melatonin and Autonomic Dysfunction
5. Possible (Sleep Related) Therapeutic Implications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Olesen, J. Headache Classification Committee of the International Headache Society (IHS) The International Classification of Headache Disorders, 3rd Edition. Cephalalgia 2018, 38, 1–211. [Google Scholar] [CrossRef]
- May, A.; Schwedt, T.J.; Magis, D.; Pozo-Rosich, P.; Evers, S.; Wang, S.J. Cluster Headache. Nat. Rev. Dis. Prim. 2018, 4, 18006. [Google Scholar] [CrossRef] [PubMed]
- Fischera, M.; Marziniak, M.; Gralow, I.; Evers, S. The Incidence and Prevalence of Cluster Headache: A Meta-Analysis of Population-Based Studies. Cephalalgia 2008, 28, 614–618. [Google Scholar] [CrossRef]
- Wei, D.Y.-T.; Yuan Ong, J.J.; Goadsby, P.J. Cluster Headache: Epidemiology, Pathophysiology, Clinical Features, and Diagnosis. Ann. Indian Acad. Neurol. 2018, 21 (Suppl. S1), S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.S.; Lund, N.; Snoer, A.; Jensen, R.H.; Barloese, M. The Economic and Personal Burden of Cluster Headache: A Controlled Cross-Sectional Study. J. Headache Pain 2022, 23, 58. [Google Scholar] [CrossRef] [PubMed]
- Barloese, M.; Lund, N.; Petersen, A.; Rasmussen, M.; Jennum, P.; Jensen, R. Sleep and Chronobiology in Cluster Headache. Cephalalgia 2015, 35, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Rozen, T.D.; Fishman, R.S. Cluster Headache in the United States of America: Demographics, Clinical Characteristics, Triggers, Suicidality, and Personal Burden. Headache 2012, 52, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Dibner, C.; Schibler, U.; Albrecht, U. The Mammalian Circadian Timing System: Organization and Coordination of Central and Peripheral Clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [Green Version]
- Russell, D. Cluster Headache: Severity and Temporal Profiles of Attacks and Patient Activity Prior to and during Attacks. Cephalalgia 1981, 1, 209–216. [Google Scholar] [CrossRef]
- Manzoni, G.C.; Terzano, M.G.; Moretti, G.; Cocchi, M. Clinical Observations on 76 Cluster Headache Cases. Eur. Neurol. 1981, 20, 88–94. [Google Scholar] [CrossRef]
- Manzoni, G.C.; Terzano, M.G.; Bono, G.; Micieli, G.; Martucci, N.; Nappi, G. Cluster Headache—Clinical Findings in 180 Patients. Cephalalgia 1983, 3, 21–30. [Google Scholar] [CrossRef]
- Leone, M.; D’Amico, D.; Moschiano, F.; Fraschini, F.; Bussone, G. Melatonin versus Placebo in the Prophylaxis of Cluster Headache: A Double-Blind Pilot Study with Parallel Groups. Cephalalgia 1996, 16, 494–496. [Google Scholar] [CrossRef]
- Gaul, C.; Christmann, N.; Schröder, D.; Weber, R.; Shanib, H.; Diener, H.C.; Holle, D. Differences in Clinical Characteristics and Frequency of Accompanying Migraine Features in Episodic and Chronic Cluster Headache. Cephalalgia 2012, 32, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Lund, N.; Barloese, M.; Petersen, A.; Haddock, B.; Jensen, R. Chronobiology Differs between Men and Women with Cluster Headache, Clinical Phenotype Does Not. Neurology 2017, 88, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Fourier, C.; Ran, C.; Steinberg, A.; Sjöstrand, C.; Waldenlind, E.; Belin, A.C. Sex Differences in Clinical Features, Treatment, and Lifestyle Factors in Patients With Cluster Headache. Neurology 2022. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Cho, S.-J.; Park, J.W.; Chu, M.K.; Moon, H.-S.; Chung, P.-W.; Chung, J.-M.; Sohn, J.-H.; Kim, B.-K.; Kim, B.-S.; et al. Temporal Changes of Circadian Rhythmicity in Cluster Headache. Cephalalgia 2020, 40, 278–287. [Google Scholar] [CrossRef]
- Barloese, M.; Haddock, B.; Lund, N.T.; Petersen, A.; Jensen, R. Chronorisk in Cluster Headache: A Tool for Individualised Therapy? Cephalalgia 2018, 38, 2058–2067. [Google Scholar] [CrossRef]
- Hagedorn, A.; Snoer, A.; Jensen, R.; Haddock, B.; Barloese, M. The Spectrum of Cluster Headache: A Case Report of 4600 Attacks. Cephalalgia 2019, 39, 1134–1142. [Google Scholar] [CrossRef]
- Ekbom, K. A Clinical Comparison of Cluster Headache and Migraine. Acta Neurol. Scand. 1970, Suppl. S41, 1+. [Google Scholar]
- Kudrow, L. The Cyclic Relationship of Natural Illumination to Cluster Period Frequency. Cephalalgia 1987, 7 (Suppl. S6), 76–78. [Google Scholar] [CrossRef]
- Ofte, H.K.; Berg, D.H.; Bekkelund, S.I.; Alstadhaug, K.B. Insomnia and Periodicity of Headache in an Arctic Cluster Headache Population. Headache 2013, 53, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.C.; Chou, K.H.; Fuh, J.L.; Lee, P.L.; Lirng, J.F.; Lin, Y.Y.; Lin, C.P.; Wang, S.J. Altered Hypothalamic Functional Connectivity in Cluster Headache: A Longitudinal Resting-State Functional MRI Study. J. Neurol. Neurosurg. Psychiatry 2015, 86, 437–445. [Google Scholar] [CrossRef]
- Barloese, M.C.J.; Beske, R.P.; Petersen, A.S.; Haddock, B.; Lund, N.; Jensen, R.H. Episodic and Chronic Cluster Headache: Differences in Family History, Traumatic Head Injury, and Chronorisk. Headache 2020, 60, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Fourier, C.; Ran, C.; Zinnegger, M.; Johansson, A.-S.; Sjöstrand, C.; Waldenlind, E.; Steinberg, A.; Belin, A.C. A Genetic CLOCK Variant Associated with Cluster Headache Causing Increased MRNA Levels. Cephalalgia 2018, 38, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Bargiello, T.A.; Young, M.W. Molecular Genetics of a Biological Clock in Drosophila. Proc. Natl. Acad. Sci. USA 1984, 81, 2142–2146. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zwiebel, L.J.; Hinton, D.; Benzer, S.; Hall, J.C.; Rosbash, M. The Period Gene Encodes a Predominantly Nuclear Protein in Adult Drosophila. J. Neurosci. Off. J. Soc. Neurosci. 1992, 12, 2735–2744. [Google Scholar] [CrossRef] [Green Version]
- Rosbash, M. A 50-Year Personal Journey: Location, Gene Expression, and Circadian Rhythms. Cold Spring Harb. Perspect. Biol. 2017, 9, a032516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Rothenfluh-Hilfiker, A.; Hunter-Ensor, M.; Chen, Y.; Myers, M.P.; Young, M.W. Rhythmic Expression of Timeless: A Basis for Promoting Circadian Cycles in Period Gene Autoregulation. Science 1995, 270, 808–810. [Google Scholar] [CrossRef]
- Price, J.L.; Blau, J.; Rothenfluh, A.; Abodeely, M.; Kloss, B.; Young, M.W. Double-Time Is a Novel Drosophila Clock Gene That Regulates PERIOD Protein Accumulation. Cell 1998, 94, 83–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darlington, T.K.; Wager-Smith, K.; Ceriani, M.F.; Staknis, D.; Gekakis, N.; Steeves, T.D.; Weitz, C.J.; Takahashi, J.S.; Kay, S.A. Closing the Circadian Loop: CLOCK-Induced Transcription of Its Own Inhibitors per and Tim. Science 1998, 280, 1599–1603. [Google Scholar] [CrossRef]
- Costa, M.; Squassina, A.; Piras, I.S.; Pisanu, C.; Congiu, D.; Niola, P.; Angius, A.; Chillotti, C.; Ardau, R.; Severino, G.; et al. Preliminary Transcriptome Analysis in Lymphoblasts from Cluster Headache and Bipolar Disorder Patients Implicates Dysregulation of Circadian and Serotonergic Genes. J. Mol. Neurosci. 2015, 56, 688–695. [Google Scholar] [CrossRef]
- Yoo, S.-H.; Mohawk, J.A.; Siepka, S.M.; Shan, Y.; Huh, S.K.; Hong, H.-K.; Kornblum, I.; Kumar, V.; Koike, N.; Xu, M.; et al. Competing E3 Ubiquitin Ligases Govern Circadian Periodicity by Degradation of CRY in Nucleus and Cytoplasm. Cell 2013, 152, 1091–1105. [Google Scholar] [CrossRef] [Green Version]
- Rains, J.C.; Poceta, J.S. Headache and Sleep Disorders: Review and Clinical Implications for Headache Management. Headache 2006, 46, 1344–1363. [Google Scholar] [CrossRef]
- Evers, S. Sleep and Headache: The Biological Basis. Headache 2010, 50, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Barloese, M.C.J.; Jennum, P.J.; Lund, N.T.; Jensen, R.H. Sleep in Cluster Headache-beyond a Temporal Rapid Eye Movement Relationship? Eur. J. Neurol. 2015, 22, 656-e40. [Google Scholar] [CrossRef]
- Barloese, M.; Jennum, P.; Knudsen, S.; Jensen, R. Cluster Headache and Sleep, Is There a Connection? A Review. Cephalalgia 2012, 32, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Della Marca, G.; Vollono, C.; Rubino, M.; Capuano, A.; Di Trapani, G.; Mariotti, P. A Sleep Study in Cluster Headache. Cephalalgia 2006, 26, 290–294. [Google Scholar] [CrossRef]
- Terzaghi, M.; Ghiotto, N.; Sances, G.; Rustioni, V.; Nappi, G.; Manni, R. Episodic Cluster Headache: NREM Prevalence of Nocturnal Attacks. Time to Look beyond Macrostructural Analysis? Headache 2010, 50, 1050–1054. [Google Scholar] [CrossRef] [PubMed]
- Lund, N.L.; Snoer, A.H.; Jennum, P.J.; Jensen, R.H.; Barloese, M.C.J. Sleep in Cluster Headache Revisited: Results from a Controlled Actigraphic Study. Cephalalgia 2019, 39, 742–749. [Google Scholar] [CrossRef]
- Barloese, M. Current Understanding of the Chronobiology of Cluster Headache and the Role of Sleep in Its Management. Nat. Sci. Sleep 2021, 13, 153–162. [Google Scholar] [CrossRef]
- Barloese, M.C.J. Neurobiology and Sleep Disorders in Cluster Headache. J. Headache Pain 2015, 16, 562. [Google Scholar] [CrossRef] [Green Version]
- de Coo, I.F.; van Oosterhout, W.P.J.; Wilbrink, L.A.; van Zwet, E.W.; Ferrari, M.D.; Fronczek, R. Chronobiology and Sleep in Cluster Headache. Headache 2019, 59, 1032–1041. [Google Scholar] [CrossRef]
- Lund, N.L.T.; Snoer, A.H.; Petersen, A.S.; Beske, R.P.; Jennum, P.J.; Jensen, R.H.; Barloese, M.C.J. Disturbed Sleep in Cluster Headache Is Not the Result of Transient Processes Associated with the Cluster Period. Eur. J. Neurol. 2019, 26, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Kovac, S.; Wright, M.-A.; Eriksson, S.H.; Zrinzo, L.; Matharu, M.; Walker, M.C. The Effect of Posterior Hypothalamus Region Deep Brain Stimulation on Sleep. Cephalalgia 2014, 34, 219–223. [Google Scholar] [CrossRef]
- Chervin, R.D.; Zallek, S.N.; Lin, X.; Hall, J.M.; Sharma, N.; Hedger, K.M. Sleep Disordered Breathing in Patients with Cluster Headache. Neurology 2000, 54, 2302–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobre, M.E.; Leal, A.J.; Filho, P.M.F. Investigation into Sleep Disturbance of Patients Suffering from Cluster Headache. Cephalalgia 2005, 25, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Graff-Radford, S.B.; Newman, A. Obstructive Sleep Apnea and Cluster Headache. Headache 2004, 44, 607–610. [Google Scholar] [CrossRef]
- Evers, S.; Barth, B.; Frese, A.; Husstedt, I.-W.; Happe, S. Sleep Apnea in Patients with Cluster Headache: A Case-Control Study. Cephalalgia 2014, 34, 828–832. [Google Scholar] [CrossRef]
- Szymusiak, R.; Gvilia, I.; McGinty, D. Hypothalamic Control of Sleep. Sleep Med. 2007, 8, 291–301. [Google Scholar] [CrossRef]
- Graff-Radford, S.B.; Teruel, A. Cluster Headache and Obstructive Sleep Apnea: Are They Related Disorders? Curr. Pain Headache Rep. 2009, 13, 160–163. [Google Scholar] [CrossRef]
- Siegel, J.M. Hypocretin (Orexin): Role in Normal Behavior and Neuropathology. Annu. Rev. Psychol. 2004, 55, 125–148. [Google Scholar] [CrossRef] [Green Version]
- Mieda, M.; Sakurai, T. Integrative Physiology of Orexins and Orexin Receptors. CNS Neurol. Disord. Drug Targets 2009, 8, 281–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, S. The Hypocretin/Orexin System in Health and Disease. Biol. Psychiatry 2003, 54, 87–95. [Google Scholar] [CrossRef]
- Swaab, D.F.; Chung, W.C.; Kruijver, F.P.; Hofman, M.A.; Hestiantoro, A. Sex Differences in the Hypothalamus in the Different Stages of Human Life. Neurobiol. Aging 2003, 24 (Suppl. S1), 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ofte, H.K.; Tronvik, E.; Alstadhaug, K.B. Lack of Association between Cluster Headache and PER3 Clock Gene Polymorphism. J. Headache Pain 2015, 17, 18. [Google Scholar] [CrossRef] [Green Version]
- May, A.; Bahra, A.; Büchel, C.; Frackowiak, R.S.; Goadsby, P.J. PET and MRA Findings in Cluster Headache and MRA in Experimental Pain. Neurology 2000, 55, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, T.; Levy, M.J.; Knight, Y.E.; Goadsby, P.J. Differential Modulation of Nociceptive Dural Input to [Hypocretin] Orexin A and B Receptor Activation in the Posterior Hypothalamic Area. Pain 2004, 109, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, S.-Y.; Yang, J.; Kong, J.; Liang, F.-R.; Lv, Z.-T. No Association Between G1246A Polymorphism in HCRTR2 Gene and Risk of Cluster Headache: Evidence From an Updated Meta-Analysis of Observational Studies. Front. Genet. 2020, 11, 560517. [Google Scholar] [CrossRef]
- Baumber, L.; Sjöstrand, C.; Leone, M.; Harty, H.; Bussone, G.; Hillert, J.; Trembath, R.C.; Russell, M.B. A Genome-Wide Scan and HCRTR2 Candidate Gene Analysis in a European Cluster Headache Cohort. Neurology 2006, 66, 1888–1893. [Google Scholar] [CrossRef]
- Cevoli, S.; Pizza, F.; Grimaldi, D.; Nicodemo, M.; Favoni, V.; Pierangeli, G.; Valko, P.O.; Baumann, C.R.; Montagna, P.; Bassetti, C.L.; et al. Cerebrospinal Fluid Hypocretin-1 Levels during the Active Period of Cluster Headache. Cephalalgia 2011, 31, 973–976. [Google Scholar] [CrossRef]
- Barloese, M.; Jennum, P.; Lund, N.; Knudsen, S.; Gammeltoft, S.; Jensen, R. Reduced CSF Hypocretin-1 Levels Are Associated with Cluster Headache. Cephalalgia 2015, 35, 869–876. [Google Scholar] [CrossRef]
- Malick, A.; Strassman, A.M.; Burstein, R. Trigeminohypothalamic and Reticulohypothalamic Tract Neurons in the Upper Cervical Spinal Cord and Caudal Medulla of the Rat. J. Neurophysiol. 2000, 84, 2078–2112. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.; Baca, S.M.; Akerman, S. Neurovascular Mechanisms of Migraine and Cluster Headache. J. Cereb. Blood Flow Metab. 2019, 39, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Vetrugno, R.; Pierangeli, G.; Leone, M.; Bussone, G.; Franzini, A.; Brogli, G.; D’Angelo, R.; Cortelli, P.; Montagna, P. Effect on Sleep of Posterior Hypothalamus Stimulation in Cluster Headache. Headache 2007, 47, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Cortelli, P.; Guaraldi, P.; Leone, M.; Pierangeli, G.; Barletta, G.; Grimaldi, D.; Cevoli, S.; Bussone, G.; Baruzzi, A.; Montagna, P. Effect of Deep Brain Stimulation of the Posterior Hypothalamic Area on the Cardiovascular System in Chronic Cluster Headache Patients. Eur. J. Neurol. 2007, 14, 1008–1015. [Google Scholar] [CrossRef]
- Bruera, O.; Sances, G.; Leston, J.; Levin, G.; Cristina, S.; Medina, C.; Barontini, M.; Nappi, G.; Figuerola, M.A.A.d.L. Plasma Melatonin Pattern in Chronic and Episodic Headaches: Evaluation during Sleep and Waking. Funct. Neurol. 2008, 23, 77–81. [Google Scholar] [PubMed]
- Kräuchi, K.; Cajochen, C.; Pache, M.; Flammer, J.; Wirz-Justice, A. Thermoregulatory Effects of Melatonin in Relation to Sleepiness. Chronobiol. Int. 2006, 23, 475–484. [Google Scholar] [CrossRef]
- Barloese, M.C.J. A Review of Cardiovascular Autonomic Control in Cluster Headache. Headache 2016, 56, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Iser, C.; Arca, K. Headache and Autonomic Dysfunction: A Review. Curr. Neurol. Neurosci. Rep. 2022, 22, 625–634. [Google Scholar] [CrossRef]
- Altiokka, O.; Mutluay, B.; Koksal, A.; Ciftci-Kavaklioglu, B.; Ozturk, M.; Altunkaynak, Y.; Baybas, S.; Soysal, A. Evaluation of Interictal Autonomic Function during Attack and Remission Periods in Cluster Headaches. Cephalalgia 2016, 36, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Barloese, M.; Brinth, L.; Mehlsen, J.; Jennum, P.; Lundberg, H.I.S.; Jensen, R. Blunted Autonomic Response in Cluster Headache Patients. Cephalalgia 2015, 35, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Barloese, M.C.J.; Mehlsen, J.; Brinth, L.; Lundberg, H.I.S.; Jennum, P.J.; Jensen, R.H. Reduced Baroreflex Sensitivity in Cluster Headache Patients. Headache 2015, 55, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Meineri, P.; Pellegrino, G.; Rosso, M.G.; Grasso, E. Systemic Autonomic Involvement in Episodic Cluster Headache: A Comparison between Active and Remission Periods. J. Headache Pain 2005, 6, 240–243. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, J.A.; Vein, A.A.; Ferrari, M.D.; van Dijk, J.G. Cardiovascular Autonomic Function Tests in Cluster Headache. Cephalalgia 2006, 26, 329–331. [Google Scholar] [CrossRef]
- Dodick, D.W.; Eross, E.J.; Parish, J.M.; Silber, M. Clinical, Anatomical, and Physiologic Relationship between Sleep and Headache. Headache 2003, 43, 282–292. [Google Scholar] [CrossRef]
- Burish, M.J.; Han, C.; Mawatari, K.; Wirianto, M.; Kim, E.; Ono, K.; Parakramaweera, R.; Chen, Z.; Yoo, S.-H. The First-Line Cluster Headache Medication Verapamil Alters the Circadian Period and Elicits Sex-Specific Sleep Changes in Mice. Chronobiol. Int. 2021, 38, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Pringsheim, T.; Magnoux, E.; Dobson, C.F.; Hamel, E.; Aubé, M. Melatonin as Adjunctive Therapy in the Prophylaxis of Cluster Headache: A Pilot Study. Headache 2002, 42, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Imai, N. A Cluster Headache Responsive to Ramelteon, a Selective Melatonin MT1/MT2 Receptor Agonist. Intern. Med. 2016, 55, 2483–2485. [Google Scholar] [CrossRef] [Green Version]
- Khatami, R.; Tartarotti, S.; Siccoli, M.M.; Bassetti, C.L.; Sándor, P.S. Long-Term Efficacy of Sodium Oxybate in 4 Patients with Chronic Cluster Headache. Neurology 2011, 77, 67–70. [Google Scholar] [CrossRef]
- Zallek, S.N.; Chervin, R.D. Improvement in Cluster Headache after Treatment for Obstructive Sleep Apnea. Sleep Med. 2000, 1, 135–138. [Google Scholar] [CrossRef]
- Sprenger, T.; Boecker, H.; Tolle, T.R.; Bussone, G.; May, A.; Leone, M. Specific Hypothalamic Activation during a Spontaneous Cluster Headache Attack. Neurology 2004, 62, 516–517. [Google Scholar] [CrossRef] [PubMed]
- Goadsby, P.J.; Dodick, D.W.; Leone, M.; Bardos, J.N.; Oakes, T.M.; Millen, B.A.; Zhou, C.; Dowsett, S.A.; Aurora, S.K.; Ahn, A.H.; et al. Trial of Galcanezumab in Prevention of Episodic Cluster Headache. N. Engl. J. Med. 2019, 381, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Snoer, A.; Vollesen, A.L.H.; Beske, R.P.; Guo, S.; Hoffmann, J.; Fahrenkrug, J.; Jørgensen, N.R.; Martinussen, T.; Jensen, R.H.; Ashina, M. Calcitonin-Gene Related Peptide and Disease Activity in Cluster Headache. Cephalalgia 2019, 39, 575–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noseda, R.; Kainz, V.; Borsook, D.; Burstein, R. Neurochemical Pathways That Converge on Thalamic Trigeminovascular Neurons: Potential Substrate for Modulation of Migraine by Sleep, Food Intake, Stress and Anxiety. PLoS ONE 2014, 9, e103929. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, M.; Tessitore, A.; Orologio, I.; De Micco, R.; Tartaglione, L.; Trojsi, F.; Tedeschi, G.; Russo, A. Galcanezumab Effect on “Whole Pain Burden” and Multidimensional Outcomes in Migraine Patients with Previous Unsuccessful Treatments: A Real-World Experience. J. Headache Pain 2022, 23, 1–14. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilati, L.; Torrente, A.; Alonge, P.; Vassallo, L.; Maccora, S.; Gagliardo, A.; Pignolo, A.; Iacono, S.; Ferlisi, S.; Di Stefano, V.; et al. Sleep and Chronobiology as a Key to Understand Cluster Headache. Neurol. Int. 2023, 15, 497-507. https://doi.org/10.3390/neurolint15010029
Pilati L, Torrente A, Alonge P, Vassallo L, Maccora S, Gagliardo A, Pignolo A, Iacono S, Ferlisi S, Di Stefano V, et al. Sleep and Chronobiology as a Key to Understand Cluster Headache. Neurology International. 2023; 15(1):497-507. https://doi.org/10.3390/neurolint15010029
Chicago/Turabian StylePilati, Laura, Angelo Torrente, Paolo Alonge, Lavinia Vassallo, Simona Maccora, Andrea Gagliardo, Antonia Pignolo, Salvatore Iacono, Salvatore Ferlisi, Vincenzo Di Stefano, and et al. 2023. "Sleep and Chronobiology as a Key to Understand Cluster Headache" Neurology International 15, no. 1: 497-507. https://doi.org/10.3390/neurolint15010029
APA StylePilati, L., Torrente, A., Alonge, P., Vassallo, L., Maccora, S., Gagliardo, A., Pignolo, A., Iacono, S., Ferlisi, S., Di Stefano, V., Camarda, C., & Brighina, F. (2023). Sleep and Chronobiology as a Key to Understand Cluster Headache. Neurology International, 15(1), 497-507. https://doi.org/10.3390/neurolint15010029