
Migraine: Advances in the Pathogenesis and Treatment
 , , ,
, , ,  and
and
Abstract
:1. Introduction
1.1. Brief Overview of Migraine as a Prevalent Neurological Condition
1.2. Prevalence of Migraine in Pediatric Patients
1.3. Medical Treatments of Migraine in Children
2. Pathogenesis of Migraine: Role of Molecular Markers in Identifying Migraine Triggers and Mechanisms
2.1. Definition and Significance of Molecular Markers
2.2. Identification of Potential Molecular Markers Associated with Migraine
- High sensitivity and specificity: this ensures that the biomarker can accurately identify individuals with a specific condition, and also correctly rule out those without the condition;
- High predictive value: the biomarker should be able to accurately forecast the course of the disease, providing valuable insights for disease management;
- Analytical stability: the biomarker should remain consistent over time and across different conditions, thereby ensuring reliable results;
- Easy, cost-effective, and minimally invasive analysis: the method of assessing the biomarker should be simple, economical, and cause minimal discomfort to the patient;
- Repeatability of method: the assessment method should yield consistent results when repeated, thereby ensuring the reliability of the biomarker.
2.2.1. Markers of Inflammation and Oxidative Stress
2.2.2. Markers Associated with Pain Transmission and Emotions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Migraine Type (Chronic Migraine [CM]/Episodic Migraine [EM]) | Action in Relation to Migraine |
|---|---|---|
| Sodium [54,55] | EM |
|
| Homocysteine [56] | EM |
|
| 3,4-Dihydroxyphenylacetic acid (DOPAC) [57] | EM |
|
| Phosphatidylcholine-specific phospholipase C [58] | EM |
|
| Transforming growth factor-β1 [59] | EM, CM |
|
| Interleukin-1 receptor antagonist [59] | EM, CM |
|
| Monocyte chemoattractant protein-1 [59] | EM, CM |
|
| Corticotrophin-releasing factor [60] | CM, MOH |
|
| Orexin-A (also referred to as hypocretin-1) [60] | CM, MOH |
|
| Glial cell line-derived neurotrophic factor [61] | CM |
|
| Somatostatin [61] | CM |
|
| Glutamate [62] | CM |
|
| Tumor necrosis factor-α [63] | CM |
|
| Taurine [64] | EM, CM |
|
| Glycine [64] | EM, CM |
|
| Glutamine [64] | EM, CM |
|
| Neuropeptide Y [65] | Acute migraine |
|
3. Other Biomarkers Associated with Increased Risk for Migraine
3.1. Genetic Markers and Migraine
- -
- FHM1: CACNA1A—This gene undergoes missense mutations resulting in a gain of function, alongside rare large exonic deletions or deletions at the 5′ non-coding end promoter. It codes for the Alpha-1 subunit of the neuronal Cav2.1 (P/Q type) voltage-gated calcium channels, crucial for modulating neuronal excitability at the presynaptic end of glutamatergic synapses;
- -
- FHM2: ATP1A2—Characterized by missense mutations, rare small deletions, or truncating mutations and frameshifts. It encodes the catalytic alpha-2 subunit of glial and neuronal ATP-dependent transmembrane Na+/K+ pumps, pivotal for extracellular K+ clearance and establishing a Na+ gradient, which is indispensable for glutamate reuptake;
- -
- FHM3: SCN1A—Experiences missense mutations (gain of function) and is responsible for the Alpha-1 subunit of neuronal Nav1.1 voltage-gated sodium channels. It plays a key role in propelling action potentials of cortical neurons, predominantly in GABAergic inhibitory interneurons;
- -
- FMH4: PRRT2—Noted for missense mutations, this gene codes for the pre-synaptic proline-rich transmembrane protein. It interacts with the synaptosomal-associated protein 25 (SNAP25), implying a potential role in merging synaptic vesicles with the plasma membrane.
| Gene Product | Migraine Type/Features | Action in Relation to Migraine |
|---|---|---|
| Dopamine type 2 (D2) receptor [23] | Migraine with and without aura [23,84] |
|
| Glutathione S-transferase [85] | Migraine without aura [85] |
|
| Dopamine type 4 (D4) receptor [86] | Migraine without aura [86] |
|
| Tumor necrosis factor-α [87] | Migraine without aura [88] |
|
| Methyltetrahydrofolate reductase (MTHFR) C677T allele [89] | Migraine with aura [89] |
|
| Dopamine β-hydroxylase gene [90] | Migraine with aura [90] |
|
| Angiotensin-converting enzyme allele [91] | Migraine with and without aura [92,93,94] |
|
| Hypocretin receptor 1 [95] | Migraine without aura [95] |
|
| Syntaxin 1A [96] | Migraine without aura [96] |
|
| Cytochrome P450 (CYP) 1A2 [97] | Chronic migraine [97] |
|
3.2. Recent Genetic Findings and Migraine
3.3. Inflammatory Indicators and Migraine
3.4. Contribution of Imaging Techniques in Understanding Migraine Pathology
3.4.1. Overview of Imaging Methods Used in Migraine Research
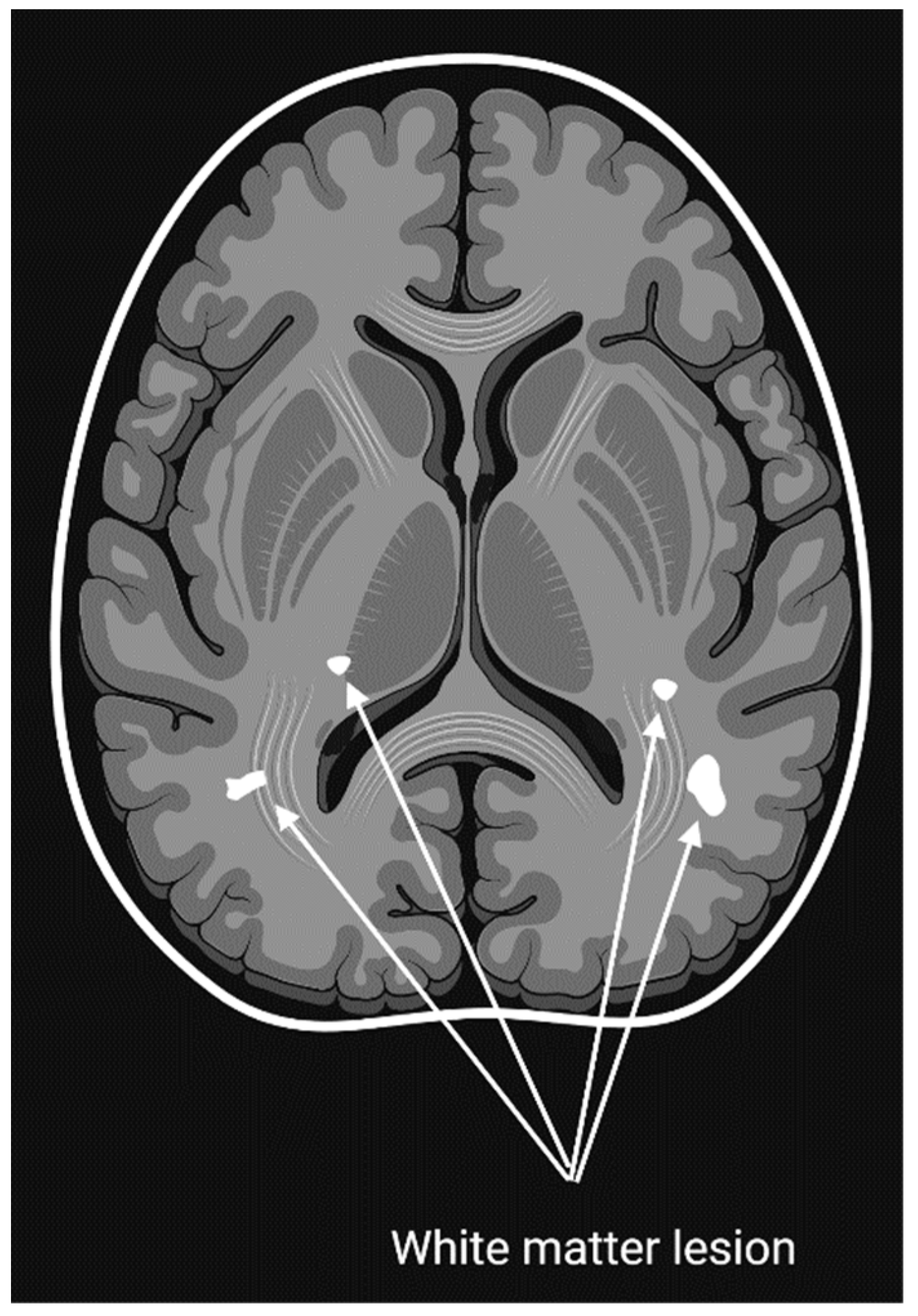
3.4.2. Findings and Insights Gained through Imaging Studies
4. Central Sensitization
Insights into Migraine Pathology: From Current Pathophysological Understanding to Peripheral Interactions and Plasma Protein Extravasation
5. Investigations into Neuropeptides
6. Headache Physiology: Central Connections and the Trigeminocervical Complex
6.1. Migraine Neuronal Activation and Therapeutic Implications
6.2. Discussion of Key Pathological Processes Involved in Migraine
6.2.1. Neurophysiology of Migraine as a Backdrop to Imaging
6.2.2. Inter-Attack Imaging Studies
- Structural Studies
- 2.
- Functional Studies
- 3.
- Premonitory Phase Studies
- 4.
- The Aura Phase
- 5.
- The Headache Stage
- 6.
- Blood–Brain Barrier (BBB): The integrity of the BBB in migraine
6.3. Calcitonin Gene-Related Peptide (CGRP) in Migraine
6.3.1. Role of CGRP in Migraine Development and Progression
- Introduction to CGRP and its significance in migraine
- 2.
- Mechanisms by which CGRP contributes to migraine symptoms
6.3.2. CGRP as a Therapeutic Target
- Overview of CGRP-targeted treatments in migraine management—Evaluation of the effectiveness of CGRP inhibitors
6.4. Medical Treatment of Migraine
6.4.1. Overview of Conventional Medical Treatments for Migraine
- Medications commonly prescribed for migraine relief—Discussion of their mechanisms of action and limitations (Figure 3)
6.4.2. Emerging Therapeutic Approaches in Migraine Management
- Introduction to novel medications and treatment strategies and Potential benefits and challenges associated with these approaches
6.5. Competitive Environment
6.5.1. The Role of Calcitonin Gene-Related Peptide in Migraine
6.5.2. CGRP Receptor Antagonists (The Gepants)
6.5.3. Ubrogepant (MK-1602)
6.5.4. Rimegepant (BMS-927711): Overview and Clinical Trials
6.5.5. Atogepant (AGN-241689): The Future of Migraine Prevention
6.5.6. Anti-CGRP and Anti-CGRP Receptor Monoclonal Antibodies
6.5.7. Erenumab (AMG334)
6.5.8. Galcanezumab (LY2951742)
6.5.9. Examination of Fremanezumab (TEV48125)
6.5.10. Eptinezumab (ALD403)
6.6. Neuromodulation in Migraine
6.6.1. Definition and Concept of Neuromodulation in Migraine Therapy
- Explanation of neuromodulation as a treatment modality
- 2.
- Overview of different types of neuromodulation techniques
- 3.
- Neuromodulation Modalities
6.6.2. Effectiveness of Neuromodulation in Managing Migraine: Review of Clinical Studies on the Use of Neuromodulation for Migraine—Assessment of the Benefits and Limitations of Neuromodulation
- Vagus Nerve Stimulation
- 2.
- Remote Electronic Neuromodulation
- 3.
- Electrical Stimulation of the Trigeminal Nerve
- 4.
- Single-Pulse Transcranial Magnetic Stimulation
- 5.
- Combined Occipital and Trigeminal Nerve Stimulation
7. Investigational Devices
7.1. Transcranial Direct Current Stimulation (tDCS)
7.2. Repetitive Transcranial Magnetic Stimulation (rTMS)
7.3. Occipital Nerve Stimulation (ONS)
7.4. Spinal Cord Stimulation (SCS)
8. Clinical Perspective
9. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stovner, L.; Hagen, K.; Jensen, R.; Katsarava, Z.; Lipton, R.; Scher, A.; Steiner, T.; Zwart, J.-A. The Global Burden of Headache: A Documentation of Headache Prevalence and Disability Worldwide. Cephalalgia 2007, 27, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Bateman, D. The future of neurology services in the UK. Pract. Neurol. 2011, 11, 134–135. [Google Scholar] [CrossRef] [PubMed]
- Sender, J.; Bradford, S.; Watson, D.; Lipscombe, S.; Rees, T.; Manley, R.; Dowson, A. Setting up a Specialist Headache Clinic in Primary Care: General Practitioners with a Special Interest (GPwSI) in Headache. Headache Care 2004, 1, 165–171. [Google Scholar] [CrossRef]
- WHO. The World Health Report 2001: Mental Health: New Understanding, New Hope; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- Solomon, G.D.; Price, K.L. Burden of Migraine: A Review of its Socioeconomic Impact. Pharmacoeconomics 1997, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Terwindt, G.M.; Ferrari, M.D.; Tijhuis, M.; Groenen, S.M.A.; Picavet, H.S.J.; Launer, L.J. The impact of migraine on quality of life in the general population: The GEM study. Neurology 2000, 55, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.; Scher, A.; Stewart, W.; Kolodner, K.; Liberman, J.; Lipton, R. The Prevalence and Disability Burden of Adult Migraine in England and their Relationships to Age, Gender and Ethnicity. Cephalalgia 2003, 23, 519–527. [Google Scholar] [CrossRef]
- Clarke, C.E.; MacMillan, L.; Sondhi, S.; Wells, N.E. Economic and social impact of migraine. QJM Mon. J. Assoc. Physicians 1996, 89, 77–84. [Google Scholar] [CrossRef]
- Ahmed, F. Headache disorders: Differentiating and managing the common subtypes. Br. J. Pain 2012, 6, 124–132. [Google Scholar] [CrossRef]
- Haut, S.R.; Bigal, E.M.; Lipton, R.B. Chronic disorders with episodic manifestations: Focus on epilepsy and migraine. Lancet Neurol. 2006, 5, 148–157. [Google Scholar] [CrossRef]
- Olesen, J. Preface to the Second Edition. Cephalalgia 2004, 24, 9–10. [Google Scholar] [CrossRef]
- Stewart, W.F.; Wood, G.C.; Manack, A.; Varon, S.F.; Buse, D.C.; Lipton, R.B. Employment and Work Impact of Chronic Migraine and Episodic Migraine. J. Occup. Environ. Med. 2010, 52, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Natoli, J.; Manack, A.; Dean, B.; Butler, Q.; Turkel, C.; Stovner, L.; Lipton, R. Global prevalence of chronic migraine: A systematic review. Cephalalgia 2009, 30, 599–609. [Google Scholar] [CrossRef]
- Blumenfeld, A.; Varon, S.; Wilcox, T.; Buse, D.; Kawata, A.; Manack, A.; Goadsby, P.; Lipton, R. Disability, HRQoL and resource use among chronic and episodic migraineurs: Results from the International Burden of Migraine Study (IBMS). Cephalalgia 2010, 31, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Durham, P.; Papapetropoulos, S. Biomarkers Associated with Migraine and Their Potential Role in Migraine Management. Headache J. Head Face Pain 2013, 53, 1262–1277. [Google Scholar] [CrossRef] [PubMed]
- Safiri, S.; Pourfathi, H.; Eagan, A.; Mansournia, M.A.; Khodayari, M.T.; Sullman, M.J.M.; Kaufman, J.; Collins, G.; Dai, H.; Bragazzi, N.L.; et al. Global, regional, and national burden of migraine in 204 countries and territories, 1990 to 2019. Pain 2021, 163, e293–e309. [Google Scholar] [CrossRef] [PubMed]
- Amiri, P.; Kazeminasab, S.; Nejadghaderi, S.A.; Mohammadinasab, R.; Pourfathi, H.; Araj-Khodaei, M.; Sullman, M.J.M.; Kolahi, A.-A.; Safiri, S. Migraine: A Review on Its History, Global Epidemiology, Risk Factors, and Comorbidities. Front. Neurol. 2022, 12, 800605. [Google Scholar] [CrossRef]
- Burch, R.C.; Loder, S.; Loder, E.; Smitherman, T.A. The Prevalence and Burden of Migraine and Severe Headache in the United States: Updated Statistics from Government Health Surveillance Studies. Headache J. Head Face Pain 2015, 55, 21–34. [Google Scholar] [CrossRef]
- Lipton, R.B.; Stewart, W.F.; Diamond, S.; Diamond, M.L.; Reed, M. Prevalence and Burden of Migraine in the United States: Data from the American Migraine Study II. Headache J. Head Face Pain 2001, 41, 646–657. [Google Scholar] [CrossRef]
- Merikangas, K.R. Contributions of Epidemiology to Our Understanding of Migraine. Headache J. Head Face Pain 2013, 53, 230–246. [Google Scholar] [CrossRef]
- Wang, S.-J. Epidemiology of migraine and other types of headache in Asia. Curr. Neurol. Neurosci. Rep. 2003, 3, 104–108. [Google Scholar] [CrossRef]
- Dzoljic, E.; Vlajinac, H.; Sipetic, S.; Marinkovic, J.; Grbatinic, I.; Kostic, V. A survey of female students with migraine: What is the influence of family history and lifestyle. Int. J. Neurosci. 2013, 124, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Latorre, M.; Roig, M. Natural History of Migraine in Childhood. Cephalalgia 2000, 20, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.B.; Olesen, J. Increased familial risk and evidence of genetic factor in migraine. BMJ 1995, 311, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.F.; Staffa, J.; Lipton, R.B.; Ottman, R. Familial risk of migraine: A population-based study. Ann. Neurol. 1997, 41, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Gervil, M.; Ulrich, V.; Kaprio, J.; Olesen, J.; Russell, M.B. The relative role of genetic and environmental factors in migraine without aura. Neurology 1999, 53, 995. [Google Scholar] [CrossRef]
- Honkasalo, M.-L.; Kaprio, J.; Winter, T.; Heikkila, K.; Sillanpaa, M.; Koskenvuo, M. Migraine and Concomitant Symptoms Among 8167 Adult Twin Pairs. Headache J. Head Face Pain 1995, 35, 70–78. [Google Scholar] [CrossRef]
- Mulder, E.J.; van Baal, C.; Gaist, D.; Kallela, M.; Kaprio, J.; Svensson, D.A.; Nyholt, D.R.; Martin, N.G.; MacGregor, A.J.; Cherkas, L.F.; et al. Genetic and Environmental Influences on Migraine: A Twin Study Across Six Countries. Twin Res. 2003, 6, 422–431. [Google Scholar] [CrossRef]
- Svensson, D.A.; Larsson, B.; Waldenlind, E.; Pedersen, N.L.; Msc, D.A.S. Shared Rearing Environment in Migraine: Results from Twins Reared Apart and Twins Reared Together. Headache J. Head Face Pain 2003, 43, 235–244. [Google Scholar] [CrossRef]
- De Vries, B.; Anttila, V.; Freilinger, T.; Wessman, M.; Kaunisto, A.M.; Kallela, M.; Artto, V.; Vijfhuizen, L.S.; Göbel, H.; Dichgans, M.; et al. Systematic re-evaluation of genes from candidate gene association studies in migraine using a large genome-wide association data set. Cephalalgia 2015, 36, 604–614. [Google Scholar] [CrossRef]
- Yeh, W.Z.; Blizzard, L.; Taylor, B.V. What is the actual prevalence of migraine. Brain Behav. 2018, 8, e00950. [Google Scholar] [CrossRef]
- Onofri, A.; Pensato, U.; Rosignoli, C.; Wells-Gatnik, W.; Stanyer, E.; Ornello, R.; Chen, H.Z.; De Santis, F.; Torrente, A.; Mikulenka, P.; et al. Primary headache epidemiology in children and adolescents: A systematic review and meta-analysis. J. Headache Pain 2023, 24, 8. [Google Scholar] [CrossRef] [PubMed]
- Loh, N.R.; Whitehouse, W.P.; Howells, R. What is new in migraine management in children and young people. Arch. Dis. Child. 2022, 107, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Loder, E.; Rizzoli, P. Biomarkers in Migraine: Their Promise, Problems, and Practical Applications. Headache J. Head Face Pain 2006, 46, 1046–1058. [Google Scholar] [CrossRef] [PubMed]
- Biomarkers Definitions Working Group; Atkinson, A.J., Jr.; Colburn, W.A.; DeGruttola, V.G.; DeMets, D.L.; Downing, G.J.; Hoth, D.F.; Oates, J.A.; Peck, C.C.; Spilker, B.A.; et al. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar] [CrossRef]
- Phillips, K.A.; Van Bebber, S.; Issa, A.M. Diagnostics and biomarker development: Priming the pipeline. Nat. Rev. Drug Discov. 2006, 5, 463–469. [Google Scholar] [CrossRef]
- Lesko, L.; Atkinson, A. Use of Biomarkers and Surrogate Endpoints in Drug Development and Regulatory Decision Making: Criteria, Validation, Strategies. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 347–366. [Google Scholar] [CrossRef]
- Packer, C.S. Biochemical markers and physiological parameters as indices for identifying patients at risk of developing pre-eclampsia. J. Hypertens. 2005, 23, 45–46. [Google Scholar] [CrossRef]
- Mayeux, R. Biomarkers: Potential uses and limitations. NeuroRx 2004, 1, 182–188. [Google Scholar] [CrossRef]
- Boćkowski, L.; Sobaniec, W.; Żelazowska-Rutkowska, B. Proinflammatory Plasma Cytokines in Children with Migraine. Pediatr. Neurol. 2009, 41, 17–21. [Google Scholar] [CrossRef]
- Kaciński, M.; Gergont, A.; Kubik, A.; Steczkowska-Klucznik, M. Proinflammatory cytokines in children with migraine with or without aura. Przeglad Lek. 2005, 62, 1276–1280. [Google Scholar]
- Kara, I.; Sazci, A.; Ergul, E.; Kaya, G.; Kilic, G. Association of the C677T and A1298C polymorphisms in the 5,10 methylenetetrahydrofolate reductase gene in patients with migraine risk. Mol. Brain Res. 2003, 111, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, M.; Prendecki, M.; Kozubski, W.; Lianeri, M.; Dorszewska, J. Molecular factors in migraine. Oncotarget 2016, 7, 50708–50718. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, G.; D’Arrigo, A.; Carbonare, M.D.; Leon, A. Pathogenesis of Migraine: Role of Neuromodulators. Headache J. Head Face Pain 2012, 52, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- D’andrea, G.; D’amico, D.; Bussone, G.; Bolner, A.; Aguggia, M.; Saracco, M.G.; Galloni, E.; De Riva, V.; Colavito, D.; Leon, A.; et al. The role of tyrosine metabolism in the pathogenesis of chronic migraine. Cephalalgia 2013, 33, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Cady, R.K.; Vause, C.V.; Ho, T.W.; Bigal, M.E.; Durham, P.L. Elevated Saliva Calcitonin Gene-Related Peptide Levels During Acute Migraine Predict Therapeutic Response to Rizatriptan. Headache J. Head Face Pain 2009, 49, 1258–1266. [Google Scholar] [CrossRef]
- Sicuteri, F.; Testi, A.; Anselmi, B. Biochemical Investigations in Headache: Increase in the Hydroxyindoleacetic Acid Excretion During Migraine Attacks. Int. Arch. Allergy Immunol. 1961, 19, 55–58. [Google Scholar] [CrossRef]
- Ferrari, M.D.; Odink, J.; Tapparelli, C.; Van Kempen, G.; Pennings, E.J.; Bruyn, G.W. Serotonin metabolism in migraine. Neurology 1989, 39, 1239. [Google Scholar] [CrossRef]
- Tsujino, N.; Sakurai, T. Orexin/Hypocretin: A Neuropeptide at the Interface of Sleep, Energy Homeostasis, and Reward System. Pharmacol. Rev. 2009, 61, 162–176. [Google Scholar] [CrossRef]
- Sarchielli, P.; Rainero, I.; Coppola, F.; Rossi, C.; Mancini, M.; Pinessi, L.; Calabresi, P. Involvement of Corticotrophin-Releasing Factor and Orexin-A in Chronic Migraine and Medication-Overuse Headache: Findings from Cerebrospinal Fluid. Cephalalgia 2008, 28, 714–722. [Google Scholar] [CrossRef]
- Barloese, M.; Jennum, P.; Lund, N.; Knudsen, S.; Gammeltoft, S.; Jensen, R. Reduced CSF hypocretin-1 levels are associated with cluster headache. Cephalalgia 2014, 35, 869–876. [Google Scholar] [CrossRef]
- Tso, A.R.; Goadsby, P.J. New Targets for Migraine Therapy. Curr. Treat. Options Neurol. 2014, 16, 318. [Google Scholar] [CrossRef]
- Diener, H.-C.; Charles, A.; Goadsby, P.J.; Holle, D. New therapeutic approaches for the prevention and treatment of migraine. Lancet Neurol. 2015, 14, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.B.; Richardson, A.K.; He, J.; Lateef, T.M.; Khoromi, S.; Merikangas, K.R. Headache and Biomarkers Predictive of Vascular Disease in a Representative Sample of US Children. Arch. Pediatr. Adolesc. Med. 2010, 164, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Tietjen, G.E.; Herial, N.A.; White, L.; Utley, C.; Kosmyna, J.M.; Khuder, S.A. Migraine and Biomarkers of Endothelial Activation in Young Women. Stroke 2009, 40, 2977–2982. [Google Scholar] [CrossRef] [PubMed]
- Ben Aissa, M.; Tipton, A.F.; Bertels, Z.; Gandhi, R.; Moye, L.S.; Novack, M.; Bennett, B.M.; Wang, Y.; Litosh, V.; Lee, S.H.; et al. Soluble guanylyl cyclase is a critical regulator of migraine-associated pain. Cephalalgia 2017, 38, 1471–1484. [Google Scholar] [CrossRef]
- Al-Hassany, L.; Boucherie, D.M.; Creeney, H.; van Drie, R.W.A.; Farham, F.; Favaretto, S.; Gollion, C.; Grangeon, L.; Lyons, H.; Marschollek, K.; et al. Future targets for migraine treatment beyond CGRP. J. Headache Pain 2023, 24, 76. [Google Scholar] [CrossRef]
- Matharu, M.S.; Bartsch, T.; Ward, N.; Frackowiak, R.S.J.; Weiner, R.; Goadsby, P.J. Central neuromodulation in chronic migraine patients with suboccipital stimulators: A PET study. Brain 2004, 127, 220–230. [Google Scholar] [CrossRef]
- Royal, P.; Andres-Bilbe, A.; Prado, P.Á.; Verkest, C.; Wdziekonski, B.; Schaub, S.; Baron, A.; Lesage, F.; Gasull, X.; Levitz, J.; et al. Migraine-Associated TRESK Mutations Increase Neuronal Excitability through Alternative Translation Initiation and Inhibition of TREK. Neuron 2019, 101, 232–245.e6. [Google Scholar] [CrossRef]
- Prado, P.; Landra-Willm, A.; Verkest, C.; Ribera, A.; Chassot, A.-A.; Baron, A.; Sandoz, G. TREK channel activation suppresses migraine pain phenotype. iScience 2021, 24, 102961. [Google Scholar] [CrossRef]
- Lim, C.S.; Shalhoub, J.; Gohel, M.S.; Shepherd, A.C.; Davies, A.H. Matrix Metalloproteinases in Vascular Disease-A Potential Therapeutic Target. Curr. Vasc. Pharmacol. 2010, 8, 75–85. [Google Scholar] [CrossRef]
- Todt, U.; Netzer, C.; Toliat, M.; Heinze, A.; Goebel, I.; Nürnberg, P.; Göbel, H.; Freudenberg, J.; Kubisch, C. New genetic evidence for involvement of the dopamine system in migraine with aura. Hum. Genet. 2009, 125, 265–279. [Google Scholar] [CrossRef]
- Naya, M.; Tsukamoto, T.; Morita, K.; Katoh, C.; Furumoto, T.; Fujii, S.; Tamaki, N.; Tsutsui, H. Plasma Interleukin-6 and Tumor Necrosis Factor-.ALPHA. Can Predict Coronary Endothelial Dysfunction in Hypertensive Patients. Hypertens. Res. 2007, 30, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Imamura, K.; Takeshima, T.; Fusayasu, E.; Nakashima, K. Increased Plasma Matrix Metalloproteinase-9 Levels in Migraineurs. Headache J. Head Face Pain 2007, 48, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.; Friberg, L.; Iversen, H.K.; Lassen, N.A.; Andersen, A.R.; Karle, A.; Olsen, T.S. Timing and topography of cerebral blood flow, aura, and headache during migraine attacks. Ann. Neurol. 1990, 28, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, L.L.; Kirchmann, M.; Bjornsson, A.; Stefansson, H.; Jensen, R.M.; Fasquel, A.C.; Petursson, H.; Stefansson, M.; Frigge, M.L.; Kong, A.; et al. The genetic spectrum of a population-based sample of familial hemiplegic migraine. Brain 2006, 130, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.L.; Diaz-Torres, S.; Bivol, S.; Cuellar-Partida, G.; Gormley, P.; Anttila, V.; Winsvold, B.S.; Palta, P.; Esko, T.; Pers, T.H.; et al. Elucidating the relationship between migraine risk and brain structure using genetic data. Brain 2022, 145, 3214–3224. [Google Scholar] [CrossRef]
- Grangeon, L.; Lange, K.S.; Waliszewska-Prosół, M.; Onan, D.; Marschollek, K.; Wiels, W.; Mikulenka, P.; Farham, F.; Gollion, C.; Ducros, A.; et al. Genetics of migraine: Where are we now. J. Headache Pain 2023, 24, 12. [Google Scholar] [CrossRef]
- van den Maagdenberg, A.M.; Pietrobon, D.; Pizzorusso, T.; Kaja, S.; Broos, L.A.; Cesetti, T.; van de Ven, R.C.; Tottene, A.; van der Kaa, J.; Plomp, J.J.; et al. A Cacna1a Knockin Migraine Mouse Model with Increased Susceptibility to Cortical Spreading Depression. Neuron 2004, 41, 701–710. [Google Scholar] [CrossRef]
- Maagdenberg, A.M.J.M.v.D.; Pizzorusso, T.; Kaja, S.; Terpolilli, N.; Shapovalova, M.; Hoebeek, F.E.; Barrett, C.F.; Gherardini, L.; van de Ven, R.C.G.; Todorov, B.; et al. High cortical spreading depression susceptibility and migraine-associated symptoms in Cav2.1 S218L mice. Ann. Neurol. 2010, 67, 85–98. [Google Scholar] [CrossRef]
- Meneghetti, N.; Cerri, C.; Vannini, E.; Tantillo, E.; Tottene, A.; Pietrobon, D.; Caleo, M.; Mazzoni, A. Synaptic alterations in visual cortex reshape contrast-dependent gamma oscillations and inhibition-excitation ratio in a genetic mouse model of migraine. J. Headache Pain 2022, 23, 125. [Google Scholar] [CrossRef]
- Leo, L.; Gherardini, L.; Barone, V.; De Fusco, M.; Pietrobon, D.; Pizzorusso, T.; Casari, G. Increased Susceptibility to Cortical Spreading Depression in the Mouse Model of Familial Hemiplegic Migraine Type 2. PLoS Genet. 2011, 7, e1002129. [Google Scholar] [CrossRef] [PubMed]
- Unekawa, M.; Ikeda, K.; Tomita, Y.; Kawakami, K.; Suzuki, N. Enhanced susceptibility to cortical spreading depression in two types of Na+,K+-ATPase α2 subunit-deficient mice as a model of familial hemiplegic migraine 2. Cephalalgia 2017, 38, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Chen, X.; Brier, L.M.; Bumstead, J.R.; Rensing, N.R.; Ringel, A.E.; Shin, H.; Oldenborg, A.; Crowley, J.R.; Bice, A.R.; et al. Astrocyte deletion of α2-Na/K ATPase triggers episodic motor paralysis in mice via a metabolic pathway. Nat. Commun. 2020, 11, 6164. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, S.; Barbieri, R.; Pusch, M.; Gavazzo, P. Gain of function of sporadic/familial hemiplegic migraine-causing SCN1A mutations: Use of an optimized cDNA. Cephalalgia 2018, 39, 477–488. [Google Scholar] [CrossRef]
- Suzuki-Muromoto, S.; Kosaki, R.; Kosaki, K.; Kubota, M. Familial hemiplegic migraine with a PRRT2 mutation: Phenotypic variations and carbamazepine efficacy. Brain Dev. 2020, 42, 293–297. [Google Scholar] [CrossRef]
- Michetti, C.; Castroflorio, E.; Marchionni, I.; Forte, N.; Sterlini, B.; Binda, F.; Fruscione, F.; Baldelli, P.; Valtorta, F.; Zara, F.; et al. The PRRT2 knockout mouse recapitulates the neurological diseases associated with PRRT2 mutations. Neurobiol. Dis. 2016, 99, 66–83. [Google Scholar] [CrossRef]
- Chasman, D.I.; Schürks, M.; Anttila, V.; De Vries, B.; Schminke, U.; Launer, L.J.; Terwindt, G.M.; van den Maagdenberg, A.M.J.M.; Fendrich, K.; Völzke, H.; et al. Genome-wide association study reveals three susceptibility loci for common migraine in the general population. Nat. Genet. 2011, 43, 695–698. [Google Scholar] [CrossRef]
- Freilinger, T.; International Headache Genetics Consortium; Anttila, V.; de Vries, B.; Malik, R.; Kallela, M.; Terwindt, G.M.; Pozo-Rosich, P.; Winsvold, B.; Nyholt, D.R.; et al. Genome-wide association analysis identifies susceptibility loci for migraine without aura. Nat. Genet. 2012, 44, 777–782. [Google Scholar] [CrossRef]
- Choquet, H.; Yin, J.; Jacobson, A.S.; Horton, B.H.; Hoffmann, T.J.; Jorgenson, E.; Avins, A.L.; Pressman, A.R. New and sex-specific migraine susceptibility loci identified from a multiethnic genome-wide meta-analysis. Commun. Biol. 2021, 4, 864. [Google Scholar] [CrossRef]
- Hautakangas, H.; Winsvold, B.S.; Ruotsalainen, S.E.; Bjornsdottir, G.; Harder, A.V.E.; Kogelman, L.J.A.; Thomas, L.F.; Noordam, R.; Benner, C.; Gormley, P.; et al. Genome-wide analysis of 102,084 migraine cases identifies 123 risk loci and subtype-specific risk alleles. Nat. Genet. 2022, 54, 152–160. [Google Scholar] [CrossRef]
- Cutrer, F.M.; Black, D.F. Imaging Findings of Migraine. Headache J. Head Face Pain 2006, 46, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Del Zompo, M.; Cherchi, A.; Palmas, M.A.; Ponti, M.; Bocchetta, A.; Gessa, G.L.; Piccardi, M.P. Association between dopamine receptor genes and migraine without aura in a Sardinian sample. Neurology 1998, 51, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, M.A.; Cutrer, F.M. Sumatriptan: A Receptor-Targeted Treatment for Migraine. Annu. Rev. Med. 1993, 44, 145–154. [Google Scholar] [CrossRef]
- Bolay, H.; Reuter, U.; Dunn, A.K.; Huang, Z.; Boas, D.A.; Moskowitz, M.A. Intrinsic brain activity triggers trigeminal meningeal afferents in a migraine model. Nat. Med. 2002, 8, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Dimtriadou, V.; Buzzi, M.; Moskowitz, M.; Theoharides, T. Trigeminal sensory fiber stimulation induces morphological changes reflecting secretion in rat dura mater mast cells. Neuroscience 1991, 44, 97–112. [Google Scholar] [CrossRef]
- Peroutka, S.J. Neurogenic inflammation and migraine: Implications for the therapeutics. Mol. Interv. 2005, 5, 304–311. [Google Scholar] [CrossRef]
- Goadsby, P.J. Pathophysiology of migraine. Ann. Indian Acad. Neurol. 2012, 15, 15. [Google Scholar] [CrossRef]
- Do, T.P.; Deligianni, C.; Amirguliyev, S.; Snellman, J.; Lopez, C.L.; Al-Karagholi, M.A.-M.; Guo, S.; Ashina, M. Second messenger signaling bypasses CGRP receptor blockade to provoke migraine attacks in humans. Brain 2023, awad261. [Google Scholar] [CrossRef]
- Amin, F.M.; Schytz, H.W. Transport of the pituitary adenylate cyclase-activating polypeptide across the blood-brain barrier: Implications for migraine. J. Headache Pain 2018, 19, 35. [Google Scholar] [CrossRef]
- Grände, G.; Nilsson, E.; Edvinsson, L. Comparison of responses to vasoactive drugs in human and rat cerebral arteries using myography and pressurized cerebral artery method. Cephalalgia 2012, 33, 152–159. [Google Scholar] [CrossRef]
- Iannetti, G.D.; Zambreanu, L.; Wise, R.G.; Buchanan, T.J.; Huggins, J.P.; Smart, T.S.; Vennart, W.; Tracey, I. Pharmacological modulation of pain-related brain activity during normal and central sensitization states in humans. Proc. Natl. Acad. Sci. USA 2005, 102, 18195–18200. [Google Scholar] [CrossRef] [PubMed]
- Birk, S.; Sitarz, J.T.; Petersen, K.A.; Oturai, P.S.; Kruuse, C.; Fahrenkrug, J.; Olesen, J. The effect of intravenous PACAP38 on cerebral hemodynamics in healthy volunteers. Regul. Pept. 2007, 140, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.; Burstein, R.; Ashina, M.; Tfelt-Hansen, P. Origin of pain in migraine: Evidence for peripheral sensitisation. Lancet Neurol. 2009, 8, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Ghanizada, H.; Al-Karagholi, M.A.-M.; Arngrim, N.; Mørch-Rasmussen, M.; Walker, C.S.; Hay, D.L.; Ashina, M. Effect of Adrenomedullin on Migraine-like Attacks in Patients 2ith Migraine: A Randomized Crossover Study. Neurology 2021, 96, e2488–e2499. [Google Scholar] [CrossRef]
- Hinson, J.P.; Kapas, S.; Smith, D.M. Adrenomedullin, a Multifunctional Regulatory Peptide. Endocr. Rev. 2000, 21, 138–167. [Google Scholar] [CrossRef] [PubMed]
- Bhakta, M.; Vuong, T.; Taura, T.; Wilson, D.S.; Stratton, J.R.; Mackenzie, K.D. Migraine therapeutics differentially modulate the CGRP pathway. Cephalalgia 2021, 41, 499–514. [Google Scholar] [CrossRef]
- Carlsson, A.; Forsgren, L.; Nylander, P.-O.; Hellman, U.; Forsman-Semb, K.; Holmgren, G.; Holmberg, D.; Holmberg, M. Identification of a susceptibility locus for migraine with and without aura on 6p12.2-p21.1. Neurology 2002, 59, 1804–1807. [Google Scholar] [CrossRef]
- Oterino, A.; Toriello, M.; Valle, N.; Castillo, J.; Alonso-Arranz, A.; Bravo, Y.; Ruiz-Alegria, C.; Quintela, E.; Pascual, J. The Relationship Between Homocysteine and Genes of Folate-Related Enzymes in Migraine Patients. Headache J. Head Face Pain 2010, 50, 99–168. [Google Scholar] [CrossRef]
- Perini, F.; D’Andrea, G.; Galloni, E.; Pignatelli, F.; Billo, G.; Alba, S.; Bussone, G.; Toso, V. Plasma Cytokine Levels in Migraineurs and Controls. Headache J. Head Face Pain 2005, 45, 926–931. [Google Scholar] [CrossRef]
- Schwedt, T.J.; Dodick, D.W. Advanced neuroimaging of migraine. Lancet Neurol. 2009, 8, 560–568. [Google Scholar] [CrossRef]
- Burstein, R.; Yamamura, H.; Malick, A.; Strassman, A.M.; Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S.; et al. Chemical Stimulation of the Intracranial Dura Induces Enhanced Responses to Facial Stimulation in Brain Stem Trigeminal Neurons. J. Neurophysiol. 1998, 79, 964–982. [Google Scholar] [CrossRef]
- Burstein, R.; Yarnitsky, D.; Goor-Aryeh, I.; Ransil, B.J.; Bajwa, Z.H. An association between migraine and cutaneous allodynia. Ann. Neurol. 2000, 47, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, M.; Silberstein, S.; Ashkenazi, A.; Burstein, R. Can allodynic migraine patients be identified interictally using a questionnaire. Neurology 2005, 65, 1419–1422. [Google Scholar] [CrossRef] [PubMed]
- Bigal, M.E.; Ashina, S.; Burstein, R.; Reed, M.L.; Buse, D.; Serrano, D.; Lipton, R.B.; On behalf of the AMPP Group. Prevalence and characteristics of allodynia in headache sufferers: A population study. Neurology 2008, 70, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Zambreanu, L.; Wise, R.G.; Brooks, J.C.; Iannetti, G.D.; Tracey, I. A role for the brainstem in central sensitisation in humans. Evidence from functional magnetic resonance imaging. Pain 2005, 114, 397–407. [Google Scholar] [CrossRef]
- Lee, M.C.; Zambreanu, L.; Menon, D.K.; Tracey, I. Identifying Brain Activity Specifically Related to the Maintenance and Perceptual Consequence of Central Sensitization in Humans. J. Neurosci. 2008, 28, 11642–11649. [Google Scholar] [CrossRef] [PubMed]
- Schulte, L.H.; Mehnert, J.; May, A. Longitudinal Neuroimaging over 30 Days: Temporal Characteristics of Migraine. Ann. Neurol. 2020, 87, 646–651. [Google Scholar] [CrossRef]
- Gross, E.C.; Lisicki, M.; Fischer, D.; Sándor, P.S.; Schoenen, J. The metabolic face of migraine—From pathophysiology to treatment. Nat. Rev. Neurol. 2019, 15, 627–643. [Google Scholar] [CrossRef]
- Ingram, E.E.; Bocklud, B.E.; Corley, S.C.; Granier, M.A.; Neuchat, E.E.; Ahmadzadeh, S.; Shekoohi, S.; Kaye, A.D. Non-CGRP Antagonist/Non-Triptan Options for Migraine Disease Treatment: Clinical Considerations. Curr. Pain Headache Rep. 2023, 1–6. [Google Scholar] [CrossRef]
- Goadsby, P.J.; Hoskin, K.L. The distribution of trigeminovascular afferents in the nonhuman primate brain Macaca nemestrina: A c-fos immunocytochemical study. J. Anat. 1997, 190, 367–375. [Google Scholar] [CrossRef]
- Bartsch, T.; Goadsby, P.J. Stimulation of the greater occipital nerve induces increased central excitability of dural afferent input. Brain 2002, 125, 1496–1509. [Google Scholar] [CrossRef] [PubMed]
- Shepheard, S.; Edvinsson, L.; Cumberbatch, M.; Williamson, D.; Mason, G.; Webb, J.; Boyce, S.; Hill, R.; Hargreaves, R. Possible Antimigraine Mechanisms of Action of the 5HT1F Receptor Agonist LY334370. Cephalalgia 1999, 19, 851–858. [Google Scholar] [CrossRef]
- Andreou, A.P.; Goadsby, P.J. Therapeutic potential of novel glutamate receptor antagonists in migraine. Expert Opin. Investig. Drugs 2009, 18, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.; Alt, A.; Ogden, A.M.; Gates, M.; Dieckman, D.K.; Clemens-Smith, A.; Ho, K.H.; Jarvie, K.; Rizkalla, G.; Wright, R.A.; et al. Pharmacological Characterization of the Competitive GLUK5 Receptor Antagonist Decahydroisoquinoline LY466195 in Vitro and in Vivo. J. Pharmacol. Exp. Ther. 2006, 318, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Coppola, G.; Vandenheede, M.; Di Clemente, L.; Ambrosini, A.; Fumal, A.; De Pasqua, V.; Schoenen, J. Somatosensory evoked high-frequency oscillations reflecting thalamo-cortical activity are decreased in migraine patients between attacks. Brain 2004, 128, 98–103. [Google Scholar] [CrossRef]
- de Tommaso, M.; Ambrosini, A.; Brighina, F.; Coppola, G.; Perrotta, A.; Pierelli, F.; Sandrini, G.; Valeriani, M.; Marinazzo, D.; Stramaglia, S.; et al. Altered processing of sensory stimuli in patients with migraine. Nat. Rev. Neurol. 2014, 10, 144–155. [Google Scholar] [CrossRef]
- Wang, W.; Timsit-Berthier, M.; Schoenen, J. Intensity dependence of auditory evoked potentials is pronounced in migraine: An indication of cortical potentiation and low serotonergic neurotransmission. Neurology 1996, 46, 1404. [Google Scholar] [CrossRef]
- Judit, Á.; Sándor, P.; Schoenen, J. Habituation of Visual and Intensity Dependence of Auditory Evoked Cortical Potentials Tends to Normalize Just Before and During the Migraine Attack. Cephalalgia 2000, 20, 714–719. [Google Scholar] [CrossRef]
- Proietti-Cecchini, A.; Áfra, J.; Schoenen, J. Intensity Dependence of the Cortical Auditory Evoked Potentials as A Surrogate Marker of Central Nervous System Serotonin Transmission in Man: Demonstration of A Central Effect for the 5Ht1B/1D Agonist Zolmitriptan (311C90, Zomig®). Cephalalgia 1997, 17, 849–854. [Google Scholar] [CrossRef]
- Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. Pathophysiology of Migraine: A Disorder of Sensory Processing. Physiol. Rev. 2017, 97, 553–622. [Google Scholar] [CrossRef]
- Valfrè, W.; Rainero, I.; Bergui, M.; Pinessi, L. Voxel-Based Morphometry Reveals Gray Matter Abnormalities in Migraine. Headache J. Head Face Pain 2007, 48, 109–117. [Google Scholar] [CrossRef]
- Maleki, N.; Becerra, L.; Nutile, L.; Pendse, G.; Brawn, J.; Bigal, M.; Burstein, R.; Borsook, D. Migraine Attacks the Basal Ganglia. Mol. Pain 2011, 7, 71. [Google Scholar] [CrossRef] [PubMed]
- DaSilva, A.F.; Granziera, C.; Tuch, D.S.; Snyder, J.; Vincent, M.; Hadjikhani, N. Interictal alterations of the trigeminal somatosensory pathway and periaqueductal gray matter in migraine. NeuroReport 2007, 18, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Coppola, G.; Tinelli, E.; Lepre, C.; Iacovelli, E.; Di Lorenzo, C.; Di Lorenzo, G.; Serrao, M.; Pauri, F.; Fiermonte, G.; Bianco, F.; et al. Dynamic changes in thalamic microstructure of migraine without aura patients: A diffusion tensor magnetic resonance imaging study. Eur. J. Neurol. 2013, 21, 287-e13. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, S.; Suh, S.-I.; Koh, S.-B.; Park, K.-W.; Oh, K. Interictal Metabolic Changes in Episodic Migraine: A Voxel-Based FDG-PET Study. Cephalalgia 2009, 30, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Vanagaite, J.; Pareja, J.; St⊘Ren, O.; White, L.; Sanc, T.; Stovner, L. Light-Induced Discomfort and Pain in Migraine. Cephalalgia 1997, 17, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Demarquay, G.; Royet, J.; Mick, G.; Ryvlin, P. Olfactory Hypersensitivity in Migraineurs: A H215O-PET Study. Cephalalgia 2008, 28, 1069–1080. [Google Scholar] [CrossRef]
- Stankewitz, A.; May, A. The phenomenon of changes in cortical excitability in migraine is not migraine-specific–A unifying thesis. Pain 2009, 145, 14–17. [Google Scholar] [CrossRef]
- Mainero, C.; Bs, J.B.; Hadjikhani, N. Altered functional magnetic resonance imaging resting-state connectivity in periaqueductal gray networks in migraine. Ann. Neurol. 2011, 70, 838–845. [Google Scholar] [CrossRef]
- Schwedt, T.J.; Berisha, V.; Chong, C.D. Temporal Lobe Cortical Thickness Correlations Differentiate the Migraine Brain from the Healthy Brain. PLoS ONE 2015, 10, e0116687. [Google Scholar] [CrossRef]
- Chong, C.D.; Gaw, N.; Fu, Y.; Li, J.; Wu, T.; Schwedt, T.J. Migraine classification using magnetic resonance imaging resting-state functional connectivity data. Cephalalgia 2016, 37, 828–844. [Google Scholar] [CrossRef] [PubMed]
- Tessitore, A.; Russo, A.; Giordano, A.; Conte, F.; Corbo, D.; De Stefano, M.; Cirillo, S.; Cirillo, M.; Esposito, F.; Tedeschi, G. Disrupted default mode network connectivity in migraine without aura. J. Headache Pain 2013, 14, 89. [Google Scholar] [CrossRef] [PubMed]
- Schoenen, J.; Jacquy, J.; Lenaerts, M. Effectiveness of high-dose riboflavin in migraine prophylaxis A randomized controlled trial. Neurology 1998, 50, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.; Levine, S.R.; D’Andrea, G.; Schultz, L.R.; Helpern, J.A. Preliminary observations on brain energy metabolism in migraine studied by in vivo phosphorus 31 NMR spectroscopy. Neurology 1989, 39, 538. [Google Scholar] [CrossRef] [PubMed]
- Montagna, P.; Cortelli, P.; Monari, L.; Pierangeli, G.; Parchi, P.; Lodi, R.; Iotti, S.; Frassineti, C.; Zaniol, P.; Lugaresi, E.; et al. 31P-Magnetic resonance spectroscopy in migraine without aura. Neurology 1994, 44, 666. [Google Scholar] [CrossRef]
- Reyngoudt, H.; Paemeleire, K.; Descamps, B.; De Deene, Y.; Achten, E. 31P-MRS demonstrates a reduction in high-energy phosphates in the occipital lobe of migraine without aura patients. Cephalalgia Int. J. Headache 2011, 31, 1243–1253. [Google Scholar] [CrossRef]
- Stankewitz, A.; Aderjan, D.; Eippert, F.; May, A. Trigeminal Nociceptive Transmission in Migraineurs Predicts Migraine Attacks. J. Neurosci. 2011, 31, 1937–1943. [Google Scholar] [CrossRef]
- Krowicki, Z.K.; Kapusta, D.R. Microinjection of Glycine into the Hypothalamic Paraventricular Nucleus Produces Diuresis, Natriuresis, and Inhibition of Central Sympathetic Outflow. J. Pharmacol. Exp. Ther. 2011, 337, 247–255. [Google Scholar] [CrossRef]
- Maniyar, F.H.; Sprenger, T.; Monteith, T.; Schankin, C.; Goadsby, P.J. Brain activations in the premonitory phase of nitroglycerin-triggered migraine attacks. Brain 2013, 137, 232–241. [Google Scholar] [CrossRef]
- Schulte, L.H.; May, A. The migraine generator revisited: Continuous scanning of the migraine cycle over 30 days and three spontaneous attacks. Brain 2016, 139, 1987–1993. [Google Scholar] [CrossRef]
- Denuelle, M.; Fabre, N.; Payoux, P.; Chollet, F.; Geraud, G. Hypothalamic Activation in Spontaneous Migraine Attacks. Headache: J. Head Face Pain 2007, 47, 1418–1426. [Google Scholar] [CrossRef]
- Lashley, K.S. Patterns of cerebral integration indicated by the scotomas of migraine. Arch. Neurol. Psychiatry 1941, 46, 331. [Google Scholar] [CrossRef]
- Leo, A.A.P.; Morison, R.S. Propagation of spreading cortical depression. J. Neurophysiol. 1945, 8, 33–45. [Google Scholar] [CrossRef]
- Lauritzen, M.; Dreier, J.P.; Fabricius, M.; Hartings, A.J.; Graf, R.; Strong, A.J. Clinical Relevance of Cortical Spreading Depression in Neurological Disorders: Migraine, Malignant Stroke, Subarachnoid and Intracranial Hemorrhage, and Traumatic Brain Injury. J. Cereb. Blood Flow Metab. 2010, 31, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.; Larsen, B.; Lauritzen, M. Focal hyperemia followed by spreading oligemia and impaired activation of rcbf in classic migraine. Ann. Neurol. 1981, 9, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Leao, A.A.P.; de Baaij, J.H.F.; Hoenderop, J.G.J.; Bindels, R.J.M.; Funke, F.; Kron, M.; Dutschmann, M.; Müller, M.; Goadsby, P.J.; Mayevsky, A.; et al. Spreading depression of activity in the cerebral cortex. J. Neurophysiol. 1944, 7, 359–390. [Google Scholar] [CrossRef]
- Hadjikhani, N.; del Rio, M.S.; Wu, O.; Schwartz, D.; Bakker, D.; Fischl, B.; Kwong, K.K.; Cutrer, F.M.; Rosen, B.R.; Tootell, R.B.H.; et al. Mechanisms of migraine aura revealed by functional MRI in human visual cortex. Proc. Natl. Acad. Sci. USA 2001, 98, 4687–4692. [Google Scholar] [CrossRef]
- Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd edition (beta version). Cephalalgia 2013, 33, 629–808. [Google Scholar] [CrossRef]
- May, A.; Kaube, H.; Büchel, C.; Eichten, C.; Rijntjes, M.; Jüptner, M.; Weiller, C.; Diener, C.H. Experimental cranial pain elicited by capsaicin: A PET study. Pain 1998, 74, 61–66. [Google Scholar] [CrossRef]
- Russell, F.A.; King, R.; Smillie, S.-J.; Kodji, X.; Brain, S.D.; Pressly, J.D.; Soni, H.; Jiang, S.; Wei, J.; Liu, R.; et al. Calcitonin Gene-Related Peptide: Physiology and Pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef]
- Stoker, K.; Baker, D.E. Erenumab-aooe. Hosp. Pharm. 2018, 53, 363–368. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, E.A. Migraine. Ann. Intern. Med. 2017, 166, ITC49–ITC64. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Adinoff, B.; DeJong, J.; Linnoila, M. Cerebrospinal fluid variables among alcoholics lack seasonal variation. Acta Psychiatr. Scand. 1991, 84, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Rashid, A.; Manghi, A. Calcitonin Gene-Related Peptide Receptor. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK560648/ (accessed on 24 July 2023).
- Messlinger, K.; Russo, A.F. Current understanding of trigeminal ganglion structure and function in headache. Cephalalgia 2018, 39, 1661–1674. [Google Scholar] [CrossRef]
- Edvinsson, J.C.A.; Warfvinge, K.; Krause, D.N.; Blixt, F.W.; Sheykhzade, M.; Edvinsson, L.; Haanes, K.A. C-fibers may modulate adjacent Aδ-fibers through axon-axon CGRP signaling at nodes of Ranvier in the trigeminal system. J. Headache Pain 2019, 20, 105. [Google Scholar] [CrossRef]
- Sacco, S.; Kurth, T. Migraine and the Risk for Stroke and Cardiovascular Disease. Curr. Cardiol. Rep. 2014, 16, 524. [Google Scholar] [CrossRef]
- Negro, A.; Martelletti, P. Gepants for the treatment of migraine. Expert Opin. Investig. Drugs 2019, 28, 555–567. [Google Scholar] [CrossRef]
- Marcus, R.; Goadsby, P.J.; Dodick, D.; Stock, D.; Manos, G.; Fischer, T.Z. BMS-927711 for the acute treatment of migraine: A double-blind, randomized, placebo controlled, dose-ranging trial. Cephalalgia 2013, 34, 114–125. [Google Scholar] [CrossRef]
- Becker, W.J. Acute Migraine Treatment in Adults. Headache J. Head Face Pain 2015, 55, 778–793. [Google Scholar] [CrossRef]
- Smith, H.S. Potential analgesic mechanisms of acetaminophen. Pain Physician 2009, 12, 269–280. [Google Scholar] [CrossRef]
- Taylor, F.R.; Kaniecki, R.G. Symptomatic Treatment of Migraine: When to Use NSAIDs, Triptans, or Opiates. Curr. Treat. Options Neurol. 2010, 13, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Colman, I.; Brown, M.D.; Innes, G.D.; Grafstein, E.; Roberts, T.E.; Rowe, B.H. Parenteral Dihydroergotamine for Acute Migraine Headache: A Systematic Review of the Literature. Ann. Emerg. Med. 2005, 45, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Silberstein, S.D.; McCrory, D.C.; Silberstein, F.S.D. Ergotamine and Dihydroergotamine: History, Pharmacology, and Efficacy. Headache J. Head Face Pain 2003, 43, 144–166. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, T.; Viana, M.; Tassorelli, C. Current Prophylactic Medications for Migraine and Their Potential Mechanisms of Action. Neurotherapeutics 2018, 15, 313–323. [Google Scholar] [CrossRef]
- Lambru, G.; Andreou, A.P.; Guglielmetti, M.; Martelletti, P. Emerging drugs for migraine treatment: An update. Expert Opin. Emerg. Drugs 2018, 23, 301–318. [Google Scholar] [CrossRef]
- Assas, M.B. Anti-migraine agents from an immunological point of view. J. Transl. Med. 2021, 19, 23. [Google Scholar] [CrossRef]
- Mathew, R.; Andreou, A.P.; Chami, L.; Bergerot, A.; van den Maagdenberg, A.M.; Ferrari, M.D.; Goadsby, P.J. Immunohistochemical characterization of calcitonin gene-related peptide in the trigeminal system of the familial hemiplegic migraine 1 knock-in mouse. Cephalalgia 2011, 31, 1368–1380. [Google Scholar] [CrossRef]
- Biella, G.; Panara, C.; Pecile, A.; Sotgiu, M. Facilitatory role of calcitonin gene-related peptide (CGRP) on excitation induced by substance P (SP) and noxious stimuli in rat spinal dorsal horn neurons. An iontophoretic study in vivo. Brain Res. 1991, 559, 352–356. [Google Scholar] [CrossRef]
- Olesen, J.; Diener, H.-C.; Husstedt, I.W.; Goadsby, P.J.; Hall, D.; Meier, U.; Pollentier, S.; Lesko, L.M. Calcitonin Gene–Related Peptide Receptor Antagonist BIBN 4096 BS for the Acute Treatment of Migraine. N. Engl. J. Med. 2004, 350, 1104–1110. [Google Scholar] [CrossRef]
- Ho, T.W.; Connor, K.M.; Zhang, Y.; Pearlman, E.; Koppenhaver, J.; Fan, X.; Lines, C.; Edvinsson, L.; Goadsby, P.J.; Michelson, D. Randomized controlled trial of the CGRP receptor antagonist telcagepant for migraine prevention. Neurology 2014, 83, 958–966. [Google Scholar] [CrossRef]
- Diener, H.-C.; Barbanti, P.; Dahlöf, C.; Reuter, U.; Habeck, J.; Podhorna, J. BI 44370 TA, an oral CGRP antagonist for the treatment of acute migraine attacks: Results from a phase II study. Cephalalgia 2010, 31, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Voss, T.; Lipton, R.B.; Dodick, D.W.; Dupre, N.; Ge, J.Y.; Bachman, R.; Assaid, C.; Aurora, S.K.; Michelson, D. A phase IIb randomized, double-blind, placebo-controlled trial of ubrogepant for the acute treatment of migraine. Cephalalgia 2016, 36, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Goadsby, P.J.; Reuter, U.; Hallström, Y.; Broessner, G.; Bonner, J.H.; Zhang, F.; Sapra, S.; Picard, H.; Mikol, D.D.; Lenz, R.A. A Controlled Trial of Erenumab for Episodic Migraine. N. Engl. J. Med. 2017, 377, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Ashina, M.; Dodick, D.; Goadsby, P.J.; Reuter, U.; Silberstein, S.; Zhang, F.; Gage, J.R.; Cheng, S.; Mikol, D.D.; Lenz, R.A. Erenumab (AMG 334) in episodic migraine: Interim analysis of an ongoing open-label study. Neurology 2017, 89, 1237–1243. [Google Scholar] [CrossRef]
- Tepper, S.; Ashina, M.; Reuter, U.; Brandes, J.L.; Doležil, D.; Silberstein, S.; Winner, P.; Leonardi, D.; Mikol, D.; Lenz, R. Safety and efficacy of erenumab for preventive treatment of chronic migraine: A randomised, double-blind, placebo-controlled phase 2 trial. Lancet Neurol. 2017, 16, 425–434. [Google Scholar] [CrossRef]
- Dodick, D.W.; Goadsby, P.J.; Spierings, E.L.H.; Scherer, J.C.; Sweeney, S.P.; Grayzel, D.S. Safety and efficacy of LY2951742, a monoclonal antibody to calcitonin gene-related peptide, for the prevention of migraine: A phase 2, randomised, double-blind, placebo-controlled study. Lancet Neurol. 2014, 13, 885–892. [Google Scholar] [CrossRef]
- Skljarevski, V.; Oakes, T.M.; Zhang, Q.; Ferguson, M.B.; Martinez, J.; Camporeale, A.; Johnson, K.W.; Shan, Q.; Carter, J.; Schacht, A.; et al. Effect of Different Doses of Galcanezumab vs Placebo for Episodic Migraine Prevention: A Randomized Clinical Trial. JAMA Neurol. 2018, 75, 187–193. [Google Scholar] [CrossRef]
- Skljarevski, V.; Matharu, M.; Millen, B.A.; Ossipov, M.H.; Kim, B.-K.; Yang, J.Y. Efficacy and safety of galcanezumab for the prevention of episodic migraine: Results of the EVOLVE-2 Phase 3 randomized controlled clinical trial. Cephalalgia 2018, 38, 1442–1454. [Google Scholar] [CrossRef]
- Bigal, M.E.; Dodick, D.W.; Rapoport, A.M.; Silberstein, S.D.; Ma, Y.; Yang, R.; Loupe, P.S.; Burstein, R.; Newman, L.C.; Lipton, R.B. Safety, tolerability, and efficacy of TEV-48125 for preventive treatment of high-frequency episodic migraine: A multicentre, randomised, double-blind, placebo-controlled, phase 2b study. Lancet Neurol. 2015, 14, 1081–1090. [Google Scholar] [CrossRef]
- Dodick, D.W.; Silberstein, S.D.; Bigal, M.E.; Yeung, P.P.; Goadsby, P.J.; Blankenbiller, T.; Grozinski-Wolff, M.; Yang, R.; Ma, Y.; Aycardi, E. Effect of Fremanezumab Compared with Placebo for Prevention of Episodic Migraine. JAMA 2018, 319, 1999–2008. [Google Scholar] [CrossRef]
- Bigal, M.E.; Dodick, D.W.; Krymchantowski, A.V.; VanderPluym, J.H.; Tepper, S.J.; Aycardi, E.; Loupe, P.S.; Ma, Y.; Goadsby, P.J. TEV-48125 for the preventive treatment of chronic migraine: Efficacy at early time points. Neurology 2016, 87, 41–48. [Google Scholar] [CrossRef]
- Silberstein, S.D.; Dodick, D.W.; Bigal, M.E.; Yeung, P.P.; Goadsby, P.J.; Blankenbiller, T.; Grozinski-Wolff, M.; Yang, R.; Ma, Y.; Aycardi, E. Fremanezumab for the Preventive Treatment of Chronic Migraine. N. Engl. J. Med. 2017, 377, 2113–2122. [Google Scholar] [CrossRef] [PubMed]
- Dodick, D.W.; Goadsby, P.J.; Silberstein, S.D.; Lipton, R.B.; Olesen, J.; Ashina, M.; Wilks, K.; Kudrow, D.; Kroll, R.; Kohrman, B.; et al. Safety and efficacy of ALD403, an antibody to calcitonin gene-related peptide, for the prevention of frequent episodic migraine: A randomised, double-blind, placebo-controlled, exploratory phase 2 trial. Lancet Neurol. 2014, 13, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M.E. Neuromodulation: Acetylcholine and memory consolidation. Trends Cogn. Sci. 1999, 3, 351–359. [Google Scholar] [CrossRef]
- Tiwari, V.; Agrawal, S. Migraine and Neuromodulation: A Literature Review. Cureus 2022, 14, e31223. [Google Scholar] [CrossRef]
- Barloese, M.; Lambru, G. Methodological Difficulties in Clinical Trials Assessing Neuromodulation Devices in the Headache Field. In Neuromodulation in Headache and Facial Pain Management; Lambru, G., Lanteri-Minet, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 227–239. [Google Scholar] [CrossRef]
- Oshinsky, M.L.; Murphy, A.L.; Hekierski, H.; Cooper, M.; Simon, B.J. Noninvasive vagus nerve stimulation as treatment for trigeminal allodynia. Pain 2014, 155, 1037–1042. [Google Scholar] [CrossRef]
- Puledda, F.; Shields, K. Non-Pharmacological Approaches for Migraine. Neurotherapeutics 2018, 15, 336–345. [Google Scholar] [CrossRef]
- Reuter, U.; McClure, C.; Liebler, E.; Pozo-Rosich, P. Non-invasive neuromodulation for migraine and cluster headache: A systematic review of clinical trials. J. Neurol. Neurosurg. Psychiatry 2019, 90, 796–804. [Google Scholar] [CrossRef]
- Yuan, H.; Chuang, T.-Y. Update of Neuromodulation in Chronic Migraine. Curr. Pain Headache Rep. 2021, 25, 71. [Google Scholar] [CrossRef]
- Schoenen, J.; Vandersmissen, B.; Jeangette, S.; Herroelen, L.; Vandenheede, M.; Gérard, P.; Magis, D. Migraine prevention with a supraorbital transcutaneous stimulator: A randomized controlled trial. Neurology 2013, 80, 697–704. [Google Scholar] [CrossRef]
- E Chou, D.; Yugrakh, M.S.; Winegarner, D.; Rowe, V.; Kuruvilla, D.; Schoenen, J. Acute migraine therapy with external trigeminal neurostimulation (ACME): A randomized controlled trial. Cephalalgia 2018, 39, 3–14. [Google Scholar] [CrossRef]
- Miller, S.; Watkins, L.; Matharu, M. Long-term outcomes of occipital nerve stimulation for chronic migraine: A cohort of 53 patients. J. Headache Pain 2016, 17, 68. [Google Scholar] [CrossRef]
- Garcia-Ortega Rn, R.; Rn, T.E.; Moir, L.; Aziz, T.Z.; Green, A.L.; Fitzgerald, J.J. Burst Occipital Nerve Stimulation for Chronic Migraine and Chronic Cluster Headache. Neuromodul. Technol. Neural Interface 2018, 22, 638–644. [Google Scholar] [CrossRef]
- Ashkan, K.; Sokratous, G.; Göbel, H.; Mehta, V.; Gendolla, A.; Dowson, A.; Wodehouse, T.; Heinze, A.; Gaul, C. Peripheral nerve stimulation registry for intractable migraine headache (RELIEF): A real-life perspective on the utility of occipital nerve stimulation for chronic migraine. Acta Neurochir. 2020, 162, 3201–3211. [Google Scholar] [CrossRef]
- De Agostino, R.; Federspiel, B.; Cesnulis, E.; Sandor, P.S. High-Cervical Spinal Cord Stimulation for Medically Intractable Chronic Migraine. Neuromodul. Technol. Neural Interface 2015, 18, 289–296. [Google Scholar] [CrossRef]
- Tassorelli, C.; Diener, H.-C.; Silberstein, S.D.; Dodick, D.W.; Goadsby, P.J.; Jensen, R.H.; Magis, D.; Pozo-Rosich, P.; Yuan, H.; Martinelli, D.; et al. Guidelines of the International Headache Society for clinical trials with neuromodulation devices for the treatment of migraine. Cephalalgia 2021, 41, 1135–1151. [Google Scholar] [CrossRef]
- Dodick, D.W. A Phase-by-Phase Review of Migraine Pathophysiology. Headache J. Head Face Pain 2018, 58, 4–16. [Google Scholar] [CrossRef]
- Blom, R.M.; Figee, M.; Vulink, N.; Denys, D. Update on repetitive transcranial magnetic stimulation in obsessive-compulsive disorder: Different targets. Curr. Psychiatry Rep. 2011, 13, 289–294. [Google Scholar] [CrossRef]
- Lan, L.; Zhang, X.; Li, X.; Rong, X.; Peng, Y. The efficacy of transcranial magnetic stimulation on migraine: A meta-analysis of randomized controlled trails. J. Headache Pain. 2017, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Shehata, H.S.; Esmail, E.H.; Abdelalim, A.; El-Jaafary, S.; Elmazny, A.; Sabbah, A.; Shalaby, N.M. Repetitive transcranial magnetic stimulation versus botulinum toxin injection in chronic migraine prophylaxis: A pilot randomized trial. J. Pain Res. 2016, 9, 771–777. [Google Scholar] [CrossRef]
- Kalita, J.; Laskar, S.; Bhoi, S.K.; Misra, U.K. Efficacy of single versus three sessions of high rate repetitive transcranial magnetic stimulation in chronic migraine and tension-type headache. J. Neurol. 2016, 263, 2238–2246. [Google Scholar] [CrossRef] [PubMed]
- Granato, A.; Fantini, J.; Monti, F.; Furlanis, G.; Musho Ilbeh, S.; Semenic, M.; Manganotti, P. Dramatic placebo effect of high frequency repetitive TMS in treatment of chronic migraine and medication overuse headache. J. Clin. Neurosci. 2019, 60, 96–100. [Google Scholar] [CrossRef] [PubMed]






| Drug Name, Type of Molecule | Indication (Acute or Prophylactic) | Development Stage |
|---|---|---|
| Atogepant, CGRP antagonist | Prophylactic | FDA-approved |
| BI 44370 TA, CGRP antagonist | Acute | Abandoned |
| Eptinezumab, CGRP monoclonal antibody | Prophylactic | FDA-approved EMA-approved |
| Erenumab, CGRP receptor monoclonal antibody | Prophylactic | FDA-approved |
| Fremanezumab, CGRP monoclonal antibody | Prophylactic | FDA-approved |
| Galcanezumab, CGRP monoclonal antibody | Prophylactic | FDA-approved |
| MK-3207, CGRP antagonist | Acute | Abandoned for liver toxicity |
| Olcegepant, CGRP antagonist | Acute | Abandoned for lack of oral availability |
| Rimegepant, CGRP antagonist | Acute | FDA-approved |
| Telcagepant, CGRP antagonist | Acute and prophylactic | Abandoned for liver toxicity |
| Ubrogepant, CGRP antagonist | Acute | FDA-approved |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pleș, H.; Florian, I.-A.; Timis, T.-L.; Covache-Busuioc, R.-A.; Glavan, L.-A.; Dumitrascu, D.-I.; Popa, A.A.; Bordeianu, A.; Ciurea, A.V. Migraine: Advances in the Pathogenesis and Treatment. Neurol. Int. 2023, 15, 1052-1105. https://doi.org/10.3390/neurolint15030067
Pleș H, Florian I-A, Timis T-L, Covache-Busuioc R-A, Glavan L-A, Dumitrascu D-I, Popa AA, Bordeianu A, Ciurea AV. Migraine: Advances in the Pathogenesis and Treatment. Neurology International. 2023; 15(3):1052-1105. https://doi.org/10.3390/neurolint15030067
Chicago/Turabian StylePleș, Horia, Ioan-Alexandru Florian, Teodora-Larisa Timis, Razvan-Adrian Covache-Busuioc, Luca-Andrei Glavan, David-Ioan Dumitrascu, Andrei Adrian Popa, Andrei Bordeianu, and Alexandru Vlad Ciurea. 2023. "Migraine: Advances in the Pathogenesis and Treatment" Neurology International 15, no. 3: 1052-1105. https://doi.org/10.3390/neurolint15030067
APA StylePleș, H., Florian, I. -A., Timis, T. -L., Covache-Busuioc, R. -A., Glavan, L. -A., Dumitrascu, D. -I., Popa, A. A., Bordeianu, A., & Ciurea, A. V. (2023). Migraine: Advances in the Pathogenesis and Treatment. Neurology International, 15(3), 1052-1105. https://doi.org/10.3390/neurolint15030067