1. Introduction
Multicellular organisms represent hosts for many viruses [
1] and interact [
2,
3]. Empirical formulas and thermodynamic properties are available in the literature for human host tissues [
4], plant host organisms [
5], and over 30 viruses [
6,
7,
8,
9,
10,
11,
12,
13]. These data are necessary for research on the biothermodynamic background of virus–host interactions.
All animate matter represents open thermodynamic systems performing growth [
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25,
26]. During growth and biosynthesis, the state of the animate matter system changes [
27], consequently, the thermodynamic properties of the organism change during biosynthesis [
28,
29,
30,
31]. Biothermodynamic research in microorganisms has been conducted in various environments, from bioreactors [
32,
33], through soil [
34,
35], to human infections [
12]. During the evolution of viruses, mutations occur that change not only the thermodynamic properties due to a change in elemental composition, but also the information content [
36,
37,
38]. In this way, the virus evolves.
Infection represents a biological interaction of a multicellular host and microorganisms [
1]. However, interactions of viruses with their hosts also represent a thermodynamic process and have been studied extensively using the approaches of biothermodynamics and bioenergetics [
12,
39,
40,
41,
42,
43,
44,
45,
46,
47,
48,
49,
50,
51,
52]. Thus, biothermodynamics is used in the analysis of the interactions of organisms with their environment and with other organisms [
53,
54,
55,
56,
57]. However, virus–host interactions have still not been explored for many viruses, first of all due to a lack of data on the elemental composition and thermodynamic properties. This is a consequence of a lack of adequate biosafety levels in most biothermodynamics and chemical analysis laboratories as well as difficulties with producing virus samples of adequate purity and in sufficient amount [
58]. Thus, the atom counting method was developed to calculate the elemental composition of various viruses [
58]. The results of the atom counting method are in agreement with the experimentally determined virus empirical formulas [
58,
59,
60].
Based on known empirical formulas of viruses and biosynthesis reactions, it is possible to find standard thermodynamic properties through the Battley, Roels, and Sandler–Orbey methods. In the Battley method, the standard enthalpy of live matter is found through the Patel–Erickson equation [
61,
62], while entropy is found using the Battley equation [
63]. Enthalpy and entropy are then combined to find the Gibbs energy. On the other hand, the Roels method uses the Roels equation to find the Gibbs energy [
64,
65] and Patel–Erickson equation to find enthalpy [
61,
62]. Combining the Gibbs energy and enthalpy gives the entropy. In the Sandler–Orbey method, enthalpy and Gibbs energy are calculated using equations proposed by Sandler and Orbey [
66,
67]. These are combined to find entropy.
Virus–host interaction represents a chemical reaction [
58,
68]. Antigen–receptor interaction is similar to protein–ligand interactions [
68,
69]. The reactions of replication and translation represent the processes of polymerization of nucleotides and amino acids, respectively, catalyzed by enzymes [
70,
71,
72]. The driving force for chemical reactions is the Gibbs energy [
20,
21,
65,
73]. Thus, it is necessary to know the Gibbs energy of biosynthesis. Biosynthesis forms virus building blocks that undergo self-assembly, forming new virions [
74,
75]. Virions accumulate inside the cell and lead to its damage [
76]. The growth of viruses is reflected in the increase in the size of the virus population [
11]. Gibbs energy of biosynthesis, Δ
bsG, is proportional to biosynthesis rate,
rbs, and thereby the multiplication rate according to the phenomenological equation
where
Lbs is the phenomenological coefficient for biosynthesis and
T is temperature [
8,
9]. Phenomenological equations belong to the domain of nonequilibrium thermodynamics, which was shown to be an excellent approach for the analysis of life processes by Prigogine and coworkers [
77,
78,
79,
80].
Phenomenological equations, also known as linear phenomenological equations, are among the oldest discoveries of thermodynamics, dating back to the early 19th century [
81]. They state that the rate of a process is proportional to its thermodynamic driving force [
15,
73]. The greater the desired rate of a process, the more useful energy must be dissipated (wasted), in order to achieve the desired rate [
15,
73]. Phenomenological equations are important, since they provide a link between thermodynamic and kinetic aspects of processes [
73]. Phenomenological equations are applicable to a wide range of processes including heat flow, viscosity, electrical energy dissipation, diffusion, and chemical reactions [
15,
73]. Their wide applicability and simplicity makes them very important tools in chemical engineering [
15,
73].
SARS-CoV-2 belongs to RNA viruses, which entered the human population for the first time in December 2019 in Wuhan [
82]. Human to human transmission was reported in January 2020 [
83]. Shortly after in March 2020, the WHO declared COVID-19 a pandemic [
84]. Until today, 623,893,894 COVID-19 cases have been reported, 6,553,936 deaths were confirmed, and 12,782,955,639 vaccine doses have been administered [
85]. Despite the large number of administered vaccines, the pandemic has not been suppressed, but only its intensity has been decreased [
86]. In Germany, in mid-October 2022, over 172 thousand new cases have been recorded daily [
87].
RNA viruses exhibit a high tendency to mutate [
88]. In 2019, the wild type SARS-CoV-2 has been identified, later labeled the Hu-1 variant. The Hu-1 variant has mutated several dozen times, which led to the appearance of new variants: Alpha, Beta, Gamma, Delta… and Omicron, with several subvariants [
89,
90,
91]. The new subvariant BA.2.75 appeared in India and has spread to over 15 countries throughout the world, but has not yet become the dominant variant [
92]. It seems that the sublineage BA.2.75 has the ability to evade the immune answer [
92,
93]. Genetic sequence data for the Omicron BA.2.75 variant are available at GISAID, the global data science initiative [
94,
95,
96]. To determine the biological potential of the BA.2.75 subvariant for spreading through the population and pathogenicity, it is necessary to estimate the susceptibility and permissiveness for this variant. In the literature, it has been reported that susceptibility to BA.2.75 is greater, since the antigen–receptor binding reaction is characterized by a lower Gibbs energy of binding [
69]. The permissiveness that influences the virus multiplication rate was the subject of analysis in this paper.
The Omicron BA.2.75 subvariant was characterized by a more negative Gibbs energy of binding than the competing BA.2 and BA.5 subvariants [
69]. This leads to faster virus entry into host cells and more rapid spreading through the population, which is in accordance with the observations made in India [
97,
98].
Recently, several analyses have appeared on the influence of entropy generation on sustainability and the development of society [
99,
100,
101]. The COVID-19 pandemic and each of the individual waves caused by various variants of SARS-CoV-2 influence the entropy generation change of the entire human society as a system. It would be interesting to make a predictive model that could extrapolate the effects of the pandemic on the generation of entropy in the future and development of society.
The aim of this paper was to find empirical formulas, molar masses as well as thermodynamic properties of live matter and biosynthesis for the Omicron BA.2.75 subvariant. Based on these data, a biothermodynamic and bioenergetic analysis of evolution of SARS-CoV-2 will be made from Hu-1, through Delta, to the Omicron BA.2.75 subvariant.
3. Results
The empirical formulas of the Omicron BA.2.75 nucleocapsids were determined for the first time and are presented in
Table 1. For the Omicron BA.2.75 isolate from Germany, the nucleocapsid empirical formula was found to be CH
1.5736O
0.3426N
0.3124P
0.00601S
0.00336. The empirical formula of the Omicron BA.2.75 nucleocapsid for the isolate from India was CH
1.5735O
0.3427N
0.3124P
0.00603S
0.00336. The empirical formula of the Omicron BA.2.75 nucleocapsid for the isolate from USA was CH
1.5737O
0.3425N
0.3123P
0.00598S
0.00336. Moreover,
Table 1 gives the molar mass (molar weight) data for the nucleocapsids for the three isolates, reported in two forms: for unit carbon formulas and for entire nucleocapsids. Molar masses of the empirical formulas were 23.75 g/C-mol for the isolates from Germany and India, and 23.74 g/C-mol for the isolate from the USA. The molar mass of the entire nucleocapsid for the Omicron BA.2.75 isolates from Germany and USA was 117.2 MDa, while that of the isolate from India was 117.1 MDa.
Table 2 shows the stoichiometry of the biosynthesis for the BA.2.75 subvariant for all three isolates.
Table 3 gives the standard thermodynamic properties of the formation for the nucleocapsids of the Omicron BA.2.75 subvariant.
Table 4 presents the data on the standard thermodynamic properties of biosynthesis for the nucleocapsids of the Omicron BA.2.75 subvariant for all three isolates. The thermodynamic properties of biosynthesis refer to the production of live matter from nutrients. Gibbs energies of biosynthesis for the nucleocapsids of the isolates from Germany, India, and USA were −221.18 kJ/C-mol, −221.24 kJ/C-mol, and −221.12 kJ/C-mol, respectively.
Table 5 gives the elemental compositions of the SARS-CoV-2 Omicron BA.2.75 subvariant for nucleic acid and proteins and the entire virion. Nucleic acid and proteins denote the viral RNA genome and all copies of proteins constituting the virus particle. Entire virion denotes the entire virus particle including the nucleic acid, all proteins, and lipids in the envelope. Elemental compositions are reported as empirical formulas, normalized per mole of carbon.
Table 6 gives the stoichiometric coefficients for the biosynthesis reactions of the SARS-CoV-2 Omicron BA.2.75 subvariant for nucleic acid and all proteins and the entire virion (including envelope lipids). Biosynthesis reactions represent the formation of live matter from nutrients as they are found in the organism’s environment.
Table 7 shows the standard thermodynamic properties of the live matter of the SARS-CoV-2 Omicron BA.2.75 subvariant for nucleic acid and proteins and the entire virion. These were then combined with biosynthesis stoichiometry from
Table 6 to find the standard thermodynamic properties of biosynthesis.
Table 8 gives the standard thermodynamic properties of the biosynthesis of the SARS-CoV-2 Omicron BA.2.75 subvariant for nucleic acid and proteins and the entire virion.
4. Discussion
In 2019, the year when COVID-19 appeared and SARS-CoV-2 was identified as the cause of the disease, the entire empirical formula of only one virus was known—the poliovirus [
59,
60] as well as partial formulas of some bacteriophages [
109]. Thermodynamic properties of human viruses were not available in the literature. Enthalpy change during the multiplication of the T4 phage in
E. coli cells was measured by Guosheng et al. [
50] and the transition from the lysogenic into lytic cycle of the Lambda phage was studied by Maskow et al. [
51] using calorimetry. This situation has created a need to determine the empirical formula and thermodynamic properties of SARS-CoV-2 and other viruses. The obstacle to determining the empirical formula of viruses is the fact that viruses are hard to obtain in sufficiently pure state and in an amount required for analysis as well as the fact that most thermodynamic laboratories do not possess an adequate biosafety level. Determining the empirical formula has become possible after the development of the atom counting method [
58]. Through application of this method, elemental compositions of the poliovirus and some other viruses were determined [
110] and compared with the values obtained experimentally [
59,
60]. It was found that the calculated results obtained using the atom counting method are in good agreement with the experimentally determined values [
58]. The empirical formula of the SARS-CoV-2 wild type (Hu-1) has been reported in [
111]. Degueldre [
112] has suggested a modified empirical formula of SARS-CoV-2 and an experimental method for the accurate measurement of virus elemental composition using mass spectrometry. Şimşek et al. [
13] computationally found the formula of the Hu-1 variant of SARS-CoV-2.
The empirical formula of the Hu-1 variant of SARS-CoV-2 nucleocapsid [
111] is CH
1.5708O
0.3452N
0.3125P
0.0060S
0.0033. SARS-CoV-2 has during the last 3 years mutated multiple times. As a consequence of mutations, there have been changes in its elemental composition and thermodynamic properties. One of the goals of this paper was to find the empirical formula of the BA.2.75 subvariant nucleocapsid. Using the atom counting method, the empirical formula of Omicron BA.2.75 was calculated and is reported in
Table 1. The empirical formula of the Omicron BA.2.75 nucleocapsid isolated in Germany was CH
1.5736O
0.3426N
0.3124P
0.00601S
0.00336. The empirical formula of the Omicron BA.2.75 nucleocapsid isolated in India was CH
1.5735O
0.3427N
0.3124P
0.00603S
0.00336. The empirical formula of the Omicron BA.2.75 nucleocapsid isolated in USA was CH
1.5737O
0.3425N
0.3123P
0.00598S
0.00336. As expected, since all three formulas were for the same BA.2.75 subvariant, the empirical formulas were almost identical for all three samples taken from different continents. Additionally, the molar masses were determined, which are given in
Table 1. The molar masses have been reported on two bases: first, the molar mass of empirical formula of the nucleocapsid was in daltons (g/C-mol),
Mr, while the second was the molar mass of the entire nucleocapsid in megadaltons,
Mr(
nc). Dividing
Mr(
nc) with
Mr gives the number of empirical formulas (C-atoms) in the entire nucleocapsid. The molar masses were, as expected, almost identical for the three BA.2.75 isolates. The molar mass of the entire nucleocapsid for the Omicron BA.2.75 isolates from Germany and USA were 117.2 MDa, while that of the isolate from India was 117.1 MDa. However, if we compare the empirical formula of BA.2.75 with that of Hu-1, we can note significant differences. The differences were the greatest in H and O content.
Table 2 gives the stoichiometric coefficients for the biosynthesis reactions of the nucleocapsids of the BA.2.75 subvariant. For the Hu-1 wild type, the biosynthesis reaction is
where AA denotes amino acids and (bio) denotes the empirical formula of the Hu-1 variant (CH
1.5708O
0.3452N
0.3125P
0.0060S
0.0033) [
111]. The stoichiometric coefficients were very similar to those of all three samples of the BA.2.75 subvariant (
Table 2).
Virus multiplication represents a biological process through an increase in the number of virus particles, resulting in the phenomenon of growth. Virus multiplication includes the replication of the virus nucleic acid, transcription, translation, self-assembly, and maturation. The replication of nucleic acids represents the polymerization reaction, based on an information template. There is polymerization of nucleotides into an identical copy of nucleic acid of the mother. The driving force for the polymerization reaction represents the Gibbs energy of biosynthesis [
8]. Transcription represents a process of the polymerization of nucleotides into messenger RNA. The driving force for this reaction is the Gibbs energy of the biosynthesis of RNA. Translation represents a reaction of protein biosynthesis, based on an information template contained in the virus nucleic acid. The polymerization of amino acids into structural and functional virus proteins is driven by the Gibbs energy of the biosynthesis of proteins [
110]. Self-assembly is a physical process of the formation of new virus particles from the synthesized virus components, using the hijacked cell metabolism. The driving force for self-assembly is also the Gibbs energy of self-assembly. The virus uses the host cell’s membrane to form new virus particles.
All of these processes require energy in the form of ATP or other nucleotide triphosphates. These are supplied by the host cell’s metabolic machinery, which has been hijacked by the virus. The energy supplied by the host cell comes from catabolism, which breaks down nutrients to obtain energy. The catabolism and the rest of the cell’s metabolic machinery are shared between the virus and its host. Therefore, they are identical for both and do not influence the competition outcome. The competition outcome is determined by the ability for biosynthesis during the biosynthetic processes of replication, transcription, and translation. The organism that performs these processes the fastest will win the competition for metabolic machinery.
Table 3 presents the standard thermodynamic properties of nucleocapsids of the three isolates of Omicron BA.2.75 subvariant. Standard enthalpies and the Gibbs energies of the formation of the three isolates were only slightly different. The standard entropies of formation were identical for all three samples. Thus, the standard thermodynamic properties of formation were very similar for all three samples, since they belonged to the same Omicron BA.2.75 subvariant.
Table 4 shows the standard thermodynamic properties of the biosynthesis of nucleocapsids of Omicron BA.2.75 subvariant samples from Germany, India, and USA. Standard thermodynamic properties of biosynthesis refer to the production of nucleocapsid live matter from nutrients [
8]. Their values were very similar for all three BA.2.75 samples. However, for the Hu-1 variant, Δ
bsH0 = −233.4 kJ/C-mol, Δ
bsS0 = −37.7 kJ/C-mol, and Δ
bsG0 = −222.2 kJ/C-mol. It can immediately be noticed that the Gibbs energy of the biosynthesis of the Omicron BA.2.75 subvariant (all three samples) was less negative than that of the Hu-1 variant. According to the evolution theory, it is expected that the SARS-CoV-2 virus evolves toward an increased infectivity and decreased or constant pathogenicity. Indeed, the Gibbs energy of antigen-receptor binding, of the Hu-1 variant is −43.4 kJ/mol [
11], while that of the Omicron BA.2.75 variant is −49.41 kJ/mol [
69]. The Gibbs energy of antigen–receptor binding is proportional to the binding rate, according to the binding phenomenological equation [
69]. This leads to the conclusion that the binding rate, and thereby infectivity, has increased during evolution from Hu-1 to BA.2.75. However, the expected decrease in pathogenicity would have to be caused by decreased rates of the multiplication and biosynthesis of virus structural elements. The rates of multiplication and biosynthesis are proportional to Gibbs energy of biosynthesis. In that case, Gibbs energy of biosynthesis would have to be the same or less negative for BA.2.75 compared to Hu-1. Indeed, the Gibbs energy of the biosynthesis of the nucleocapsid for BA.2.75 is −221.2 kJ/C-mol, while that of Hu-1 is −222.2 kJ/C-mol. Thus, the Gibbs energy of the biosynthesis of BA.2.75 was slightly less negative, implying a lower rate of biosynthesis, multiplication, accumulation of viruses in host cells, and damage to host cells. Thus, it seems that the BA.2.75 subvariant has evolved exactly as expected by the theory of evolution, regarding the decrease in pathogenicity.
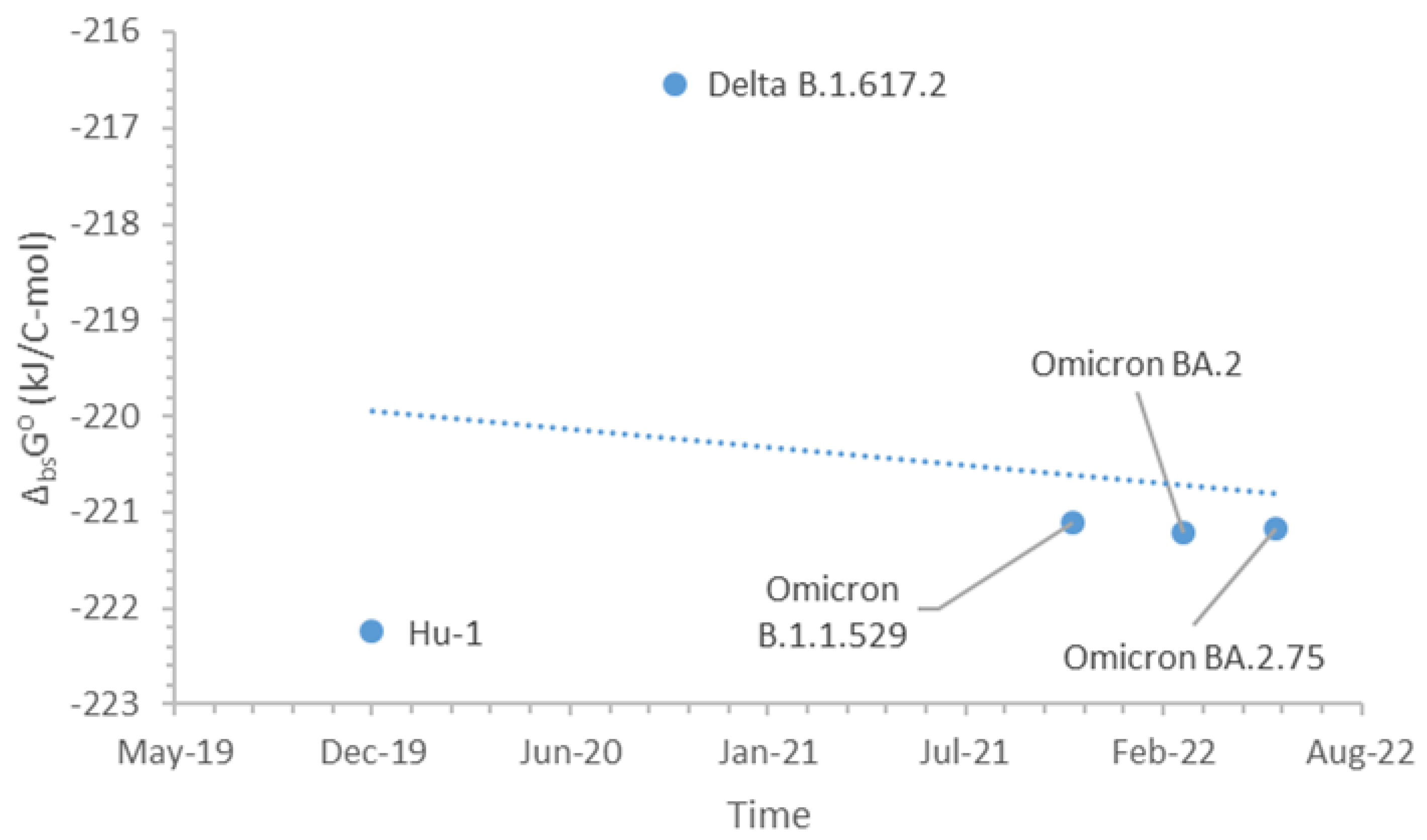
Figure 1 shows the standard Gibbs energies of the biosynthesis of nucleocapsids and their dates of appearance of SARS-CoV-2 variants including Hu-1 (Wild type), Delta B.1.617.2, Omicron B.1.1.529, Omicron BA.2, and Omicron BA.2.75. From the graph, it is possible to see the evolution of SARS-CoV-2 related to the multiplication inside the host cells. The trend of evolution is given by the dotted line. The values for the Omicron B.1.1.529, Omicron BA.2, and Omicron BA.2.75 variants were very similar. This implies that the mutations mostly occurred in the part of the nucleic acid related to the binding domain, rather than multiplication inside the host cell.
Table 8 provides the thermodynamic properties of biosynthesis for the nucleic acid and proteins as well as entire virus particles. There was a significant difference in the thermodynamic properties of the biosynthesis of nucleocapsids (nucleic acid with nucleoproteins) and nucleic acid with all of the proteins. Standard Gibbs energies of the biosynthesis of BA.2.75 nucleic acid and proteins was close to −174.5 kJ/C-mol. For the entire virus particles, the calculated standard Gibbs energy of biosynthesis was −9.45 kJ/C-mol.
The virus–host interaction occurred at two sites: at the host cell’s membrane (antigen-receptor binding) and in the cytoplasm (virus multiplication). Both processes represent chemical reactions and are driven by Gibbs energy. Antigen–receptor binding is driven by the Gibbs energy of binding, while virus multiplication is driven by the Gibbs energy of biosynthesis. Infectivity and transmissibility depend on the binding rate and Gibbs energy of binding. The Gibbs energy of binding is specific for each antigen–receptor pair and depends on the 3D structures of the antigen and receptor, which determine the intermolecular forces such as hydrogen bonds, van der Waals forces, hydrophobic interactions, etc. All of these interactions are taken into account in the Gibbs energy of binding, which is measured experimentally using methods such as surface plasmon resonance [
113,
114] or the non-competitive ELISA approach [
115,
116]. An analysis of Omicron BA.2.75 subvariant infectivity based on thee Gibbs energy of binding can be found in [
69]. This research did not take into account the immune response, which would be an interesting subject for future research. Pathogenicity is a biological phenomenon, which is a consequence of the virus multiplication rate as well as the damage that occurs on the host cell during virus multiplication. The virus multiplication rate depends on its thermodynamic driving force—the Gibbs energy of biosynthesis. The Gibbs energy of biosynthesis and other thermodynamic properties can be calculated using standard thermodynamic tools, which are used for other microorganisms including the predictive biothermodynamic models (e.g., Patel–Erickson, Battley, Roels), growth reactions, thermodynamic properties of biosynthesis, etc.
The mechanistic biothermodynamic model that was presented in this paper is applicable to pathogen–host interactions of virions, subcellular, and cellular organisms. If we are discussing virus–host interactions, the hosts are cells of various tissues. On the other hand, if we are analyzing bacteria–host interactions, the hosts are various tissues and organisms, which are the subject of the interaction with microorganisms. The model is the same, but the analysis uses the thermodynamic properties of appropriate cells in the case of interactions with viruses, or tissues in the case of interaction with bacteria. Comparing the growth rates of microorganisms with those of cells and tissues provides an assessment of the ability of microorganisms to hijack the building blocks from the host.
5. Conclusions
The empirical formulas of the BA.2.75 subvariant nucleocapsid were determined for three isolates from Germany, India, and the USA. The empirical formulas and thermodynamic properties were very similar for the three isolates. The empirical formula of the isolate from Germany was CH1.5736O0.3426N0.3124P0.00601S0.00336. For the isolate from India, it was CH1.5735O0.3427N0.3124P0.00603S0.00336. Finally, for the isolate from the USA, it was CH1.5737O0.3425N0.3123P0.00598S0.00336. The similar empirical formulas can be explained by the three isolates belonging to the same BA.2.75 subvariant.
Molar masses were determined for the Omicron BA.2.75 subvariant nucleocapsids. Molar masses were reported on two bases: the molar masses of empirical formulas and the molar masses of entire nucleocapsids. The molar masses of thee empirical formulas were 23.75 g/C-mol for the isolates from Germany and India, and 23.74 g/C-mol for the isolate from the USA. The molar masses of the entire nucleocapsids for the isolates from Germany and India were 117.2 MDa, while that of the isolate from the USA was 117.1 MDa. Therefore, the molar masses were very similar for all three isolates of the Omicron BA.2.75 subvariant.
The Gibbs energy of biosynthesis was calculated for nucleocapsids of the Omicron BA.2.75 subvariant. The Gibbs energies of the biosynthesis for the nucleocapsids of the isolates from Germany, India, and the USA were −221.18 kJ/C-mol, −221.24 kJ/C-mol, and −221.12 kJ/C-mol, respectively. The three values were very similar to each other. However, they were all slightly less negative compared to that of the Hu-1 wild type, which was −222.2 kJ/C-mol. Thus, even though there is a great homogeneity in the elemental composition and thermodynamic properties within the Omicron BA.2.75 subvariant, there is a difference compared to the Hu-1 wild type.
Due to the less negative Gibbs energy of nucleocapsid biosynthesis, the pathogenicity of the Omicron BA.2.75 subvariant should be slightly lower than that of Hu-1. This is in accordance with the predictions of evolution theory. Mutations that appeared during evolution from Hu-1 to BA.2.75 variant have led to a decrease in the Gibbs energy of biosynthesis and thereby a decrease in the rates of virion multiplication, virion accumulation inside the host cells, and damage to host cells.






{kind=link}