
Multiple Mechanisms Synergistically Induce Pseudomonas Aeruginosa Multiple Drug Resistance

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Antimicrobial Susceptibility Testing
2.3. Detection of Carbapenemase Production
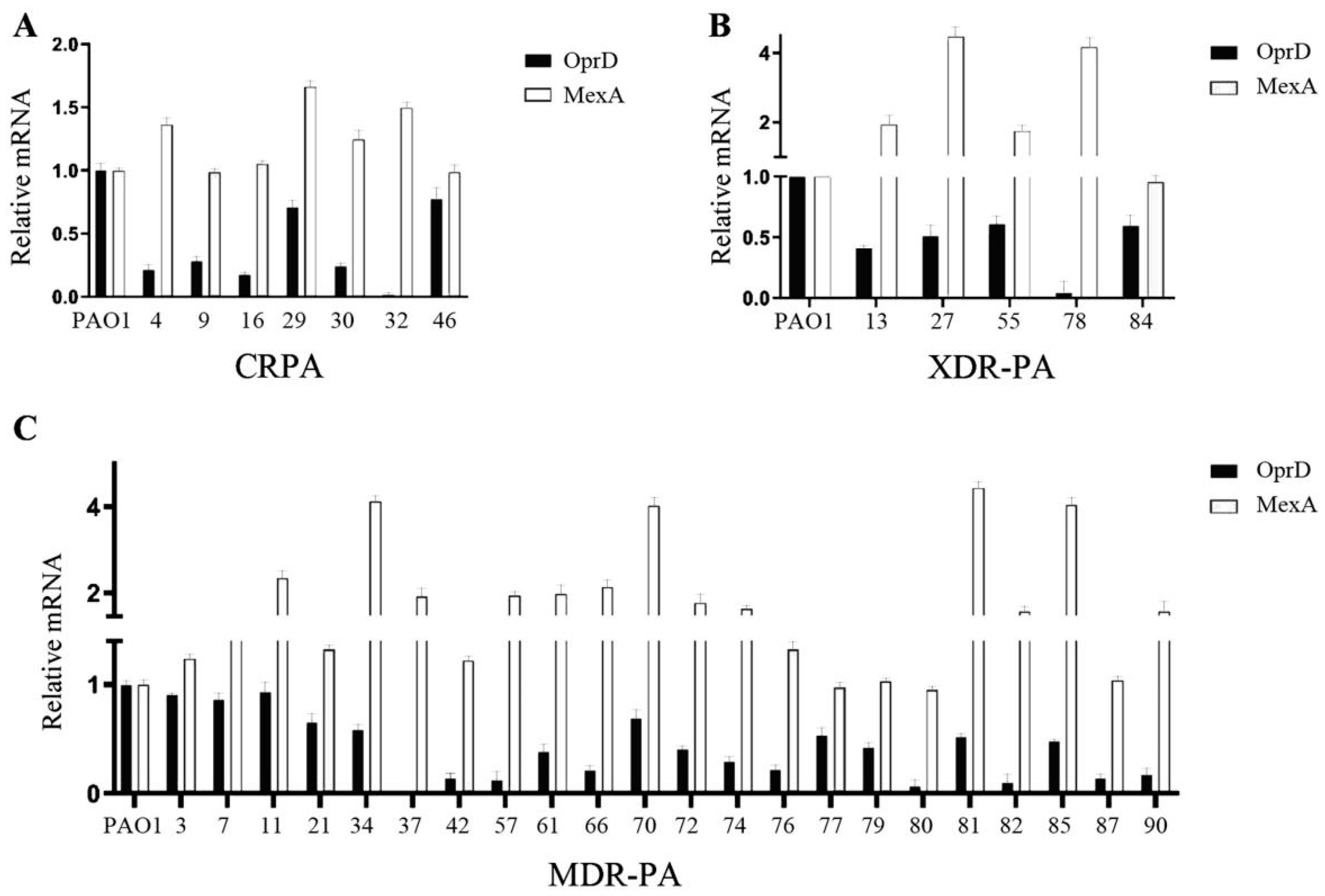
2.4. RT-qPCR Was Used to Detect the Expression Levels of Mex A and OprD
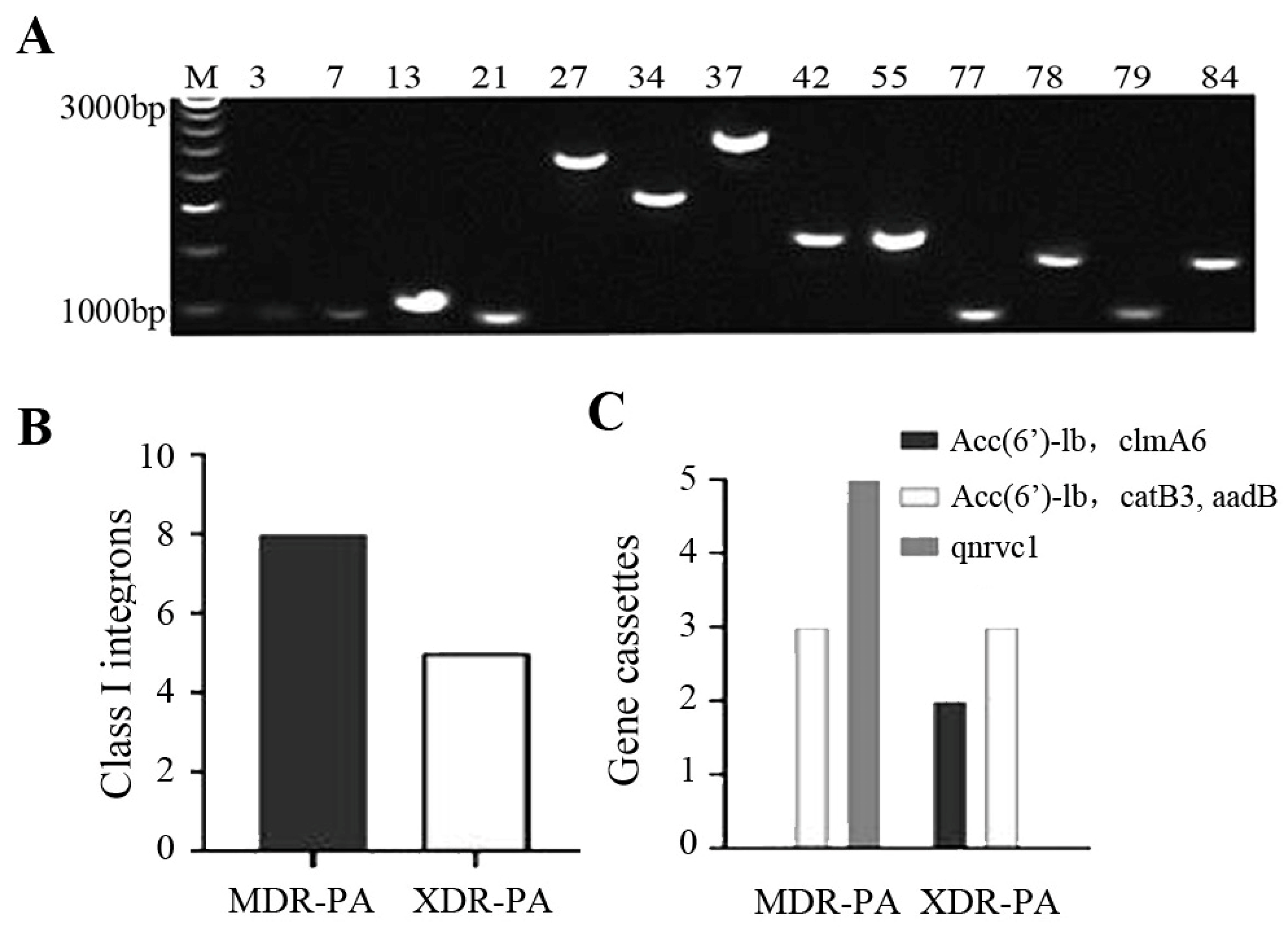
2.5. PCR Amplification and Sequencing of Class I Integron
2.6. Statistical Analysis
3. Results
3.1. Antibiotic Sensitivity
3.2. Carbapenemase Production
3.3. Gene Expression Analysis
3.4. PCR Amplification and Sequencing of Class I Integron
3.5. CRPA Resistance Characteristics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nordmann, P.; Poirel, L. Epidemiology and Diagnostics of Carbapenem Resistance in Gram-negative Bacteria. Clin. Infect. Dis. 2019, 69, S521–S528. [Google Scholar] [CrossRef] [PubMed]
- Ruden, S.; Rieder, A.; Chis Ster, I.; Schwartz, T.; Mikut, R.; Hilpert, K. Synergy Pattern of Short Cationic Antimicrobial Peptides Against Multidrug-Resistant Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 2740. [Google Scholar] [CrossRef] [PubMed]
- Cacci, L.C.; Chuster, S.G.; Martins, N.; Carmo, P.R.; Girao, V.B.; Nouer, S.A.; Freitas, W.V.; Matos, J.A.; Magalhaes, A.C.; Ferreira, A.L.; et al. Mechanisms of carbapenem resistance in endemic Pseudomonas aeruginosa isolates after an S.P.M.-1 metallo-beta-lactamase producing strain subsided in an intensive care unit of a teaching hospital in Brazil. Mem. Inst. Oswaldo Cruz 2016, 111, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorli, L.; Luque, S.; Gomez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32, e00031-19. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, D.; Zhou, W.; Sang, H.; Liu, X.; Ge, Z.; Zhang, J.; Lan, L.; Yang, C.G.; Chen, H. Novobiocin binding to NalD induces the expression of the MexAB-OprM pump in Pseudomonas aeruginosa. Mol. Microbiol. 2016, 100, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.P.; Xu, Y.H.; Wang, Z.X.; Fang, Y.P.; Shen, J.L. Overexpression of MexAB-OprM efflux pump in carbapenem-resistant Pseudomonas aeruginosa. Arch. Microbiol. 2016, 198, 565–571. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, A.; Mutters, N.T.; Mastroianni, C.M.; Kaiser, S.J.; Gunther, F. Distribution of carbapenem resistance mechanisms in clinical isolates of XDR Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Bouslah, Z. Carba NP test for the detection of carbapenemase-producing Pseudomonas aeruginosa. Med. Mal. Infect. 2020, 50, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Suresh, M.; Nithya, N.; Jayasree, P.R.; Vimal, K.P.; Manish Kumar, P.R. Mutational analyses of regulatory genes, mexR, nalC, nalD and mexZ of mexAB-oprM and mexXY operons, in efflux pump hyperexpressing multidrug-resistant clinical isolates of Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2018, 34, 83. [Google Scholar] [CrossRef] [PubMed]
- Levesque, C.; Piche, L.; Larose, C.; Roy, P.H. PCR mapping of integrons reveals several novel combinations of resistance genes. Antimicrob. Agents Chemother. 1995, 39, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Ayoub Moubareck, C.; Hammoudi Halat, D.; Akkawi, C.; Nabi, A.; AlSharhan, M.A.; AlDeesi, Z.O.; Peters, C.C.; Celiloglu, H.; Karam Sarkis, D. Role of outer membrane permeability, efflux mechanism, and carbapenemases in carbapenem-nonsusceptible Pseudomonas aeruginosa from Dubai hospitals: Results of the first cross-sectional survey. Int. J. Infect. Dis. 2019, 84, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Kang, H.Y.; Kim, B.R.; Jeon, H.; Lee, Y.C.; Lee, S.H.; Lee, J.C. Mutational inactivation of OprD in carbapenem-resistant Pseudomonas aeruginosa isolates from Korean hospitals. J. Microbiol. 2016, 54, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Le-Vo, H.N.; Tran, P.T.; Le, L.; Matsumoto, Y.; Motooka, D.; Nakamura, S.; Jones, J.W.; Iida, T.; Cao, V. Complex Class 1 Integron in a Clinical Escherichia coli Strain From Vietnam Carrying Both mcr-1 and bla NDM-1. Front. Microbiol. 2019, 10, 2472. [Google Scholar] [CrossRef] [PubMed]
- Karampatakis, T.; Antachopoulos, C.; Tsakris, A.; Roilides, E. Molecular epidemiology of carbapenem-resistant Pseudomonas aeruginosa in an endemic area: Comparison with global data. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Kiiru, J.; Butaye, P.; Goddeeris, B.M.; Kariuki, S. Analysis for prevalence and physical linkages amongst integrons, ISEcp1, ISCR1, Tn21 and Tn7 encountered in Escherichia coli strains from hospitalized and non-hospitalized patients in Kenya during a 19-year period (1992–2011). BMC Microbiol. 2013, 13, 109. [Google Scholar] [CrossRef] [PubMed]
- Khalili, Y.; Yekani, M.; Goli, H.R.; Memar, M.Y. Characterization of carbapenem-resistant but cephalosporin-susceptible Pseudomonas aeruginosa. Acta Microbiol. Immunol. Hung. 2019, 66, 529–540. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genes | Primers | Primers Sequences(5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| rpsL | F | CGCAACGTCGTGGCGTAT | 226 |
| R | ACCCGAGGTGTCCAGCGAAC | ||
| OprD | F | TTTCAACATCTACCGCACAAA | 389 |
| R | CGTAGCCGTAGTTCTTATAGCC | ||
| MexA | F | GGCCGTGAGCAAGCAGCAGT | 377 |
| R | CGACGGAAACCTCGGAGAA | ||
| IntI1 | F | GGCATCCAAGCAGCAAG | Variable |
| R | AAGCAGACTTGACCTGA |
| Strains | Drug Resistance Classification | Size | Resistance Genes |
|---|---|---|---|
| 34, 37, 42 | MDR | 1.1–3.8 k | Acc(6′)-lb, catB3, aadB3 |
| 13, 27, 55 | XDR | ||
| 78, 84 | XDR | 1.4 k | Acc(6′)-lb, clmA6 |
| 3, 7, 21, 77, 79 | MDR | 1.1 k | qnrvc1 |
| Strains | Carbe NP Test | Class I Integron | Relative mRNA | IMP | MEM | CAZ | Drug Resistance | |
|---|---|---|---|---|---|---|---|---|
| OprD | Mex A | MIC (µg/mL) | ||||||
| 3 | − | + | − | − | ≥32 | ≥32 | 32 | MDR-PA |
| 4 | − | − | ↓ | − | ≥32 | 4 | 0.5 | CRPA |
| 7 | − | + | − | − | ≥32 | ≥32 | 1 | MDR-PA |
| 9 | − | − | ↓ | − | ≥32 | 4 | 1 | CRPA |
| 11 | − | − | − | ↑ | ≥32 | ≥32 | 0.5 | MDR-PA |
| 13 | + | + | − | − | ≥32 | ≥32 | 16 | XDR-PA |
| 16 | − | − | ↓ | − | ≥32 | ≥32 | 0.5 | CRPA |
| 21 | − | + | − | − | ≥32 | ≥32 | 1 | MDR-PA |
| 27 | + | + | − | ↑ | ≥32 | ≥32 | 256 | XDR-PA |
| 29 | − | − | − | − | 8 | ≥32 | 0.5 | CRPA |
| 30 | − | − | ↓ | − | ≥32 | ≥32 | 12 | CRPA |
| 32 | − | − | ↓ | − | ≥32 | 4 | 0.5 | CRPA |
| 34 | + | + | − | ↑ | ≥32 | ≥32 | 0.75 | MDR-PA |
| 37 | − | + | ↓ | − | ≥32 | ≥32 | 1 | MDR-PA |
| 42 | − | + | ↓ | − | ≥32 | ≥32 | 1 | MDR-PA |
| 46 | − | − | − | − | ≥32 | 4 | 0.5 | CRPA |
| 55 | + | + | − | − | ≥32 | ≥32 | 0.5 | XDR-PA |
| 57 | − | − | ↓ | − | ≥32 | ≥32 | 0.5 | MDR-PA |
| 61 | − | − | − | − | ≥32 | ≥32 | 32 | MDR-PA |
| 66 | − | − | ↓ | ↑ | ≥32 | ≥32 | 1 | MDR-PA |
| 70 | − | − | − | ↑ | ≥32 | ≥32 | 32 | MDR-PA |
| 72 | − | − | − | − | ≥32 | ≥32 | 256 | MDR-PA |
| 74 | − | − | ↓ | − | ≥32 | 4 | 1 | MDR-PA |
| 76 | − | − | ↓ | − | ≥32 | ≥32 | 16 | MDR-PA |
| 77 | − | + | − | − | ≥32 | ≥32 | 16 | MDR-PA |
| 78 | + | + | ↓ | ↑ | ≥32 | ≥32 | 256 | XDR-PA |
| 79 | − | + | − | − | ≥32 | ≥32 | 1 | MDR-PA |
| 80 | − | − | ↓ | − | ≥32 | ≥32 | 0.5 | MDR-PA |
| 81 | − | − | − | ↑ | ≥32 | ≥32 | 0.5 | MDR-PA |
| 82 | − | − | ↓ | − | ≥32 | ≥32 | 1 | MDR-PA |
| 84 | + | + | − | − | ≥32 | ≥32 | 256 | XDR-PA |
| 85 | − | − | − | ↑ | ≥32 | ≥32 | 1 | MDR-PA |
| 87 | − | − | ↓ | − | ≥32 | ≥32 | 1 | MDR-PA |
| 90 | − | − | ↓ | − | ≥32 | 12 | 0.5 | MDR-PA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, P.; Jiao, F.; Yang, L.; Bajinka, O.; Abdelhalim, K.A.; Wu, G.; Tan, Y. Multiple Mechanisms Synergistically Induce Pseudomonas Aeruginosa Multiple Drug Resistance. Microbiol. Res. 2023, 14, 627-634. https://doi.org/10.3390/microbiolres14020044
Dai P, Jiao F, Yang L, Bajinka O, Abdelhalim KA, Wu G, Tan Y. Multiple Mechanisms Synergistically Induce Pseudomonas Aeruginosa Multiple Drug Resistance. Microbiology Research. 2023; 14(2):627-634. https://doi.org/10.3390/microbiolres14020044
Chicago/Turabian StyleDai, Pei, Fangyan Jiao, Lulu Yang, Ousman Bajinka, Khalid A. Abdelhalim, Guojun Wu, and Yurong Tan. 2023. "Multiple Mechanisms Synergistically Induce Pseudomonas Aeruginosa Multiple Drug Resistance" Microbiology Research 14, no. 2: 627-634. https://doi.org/10.3390/microbiolres14020044
APA StyleDai, P., Jiao, F., Yang, L., Bajinka, O., Abdelhalim, K. A., Wu, G., & Tan, Y. (2023). Multiple Mechanisms Synergistically Induce Pseudomonas Aeruginosa Multiple Drug Resistance. Microbiology Research, 14(2), 627-634. https://doi.org/10.3390/microbiolres14020044