An Integrated Biomarker Approach Using Flounder to Improve Chemical Risk Assessments in the Heavily Polluted Seine Estuary

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Metal Analysis
2.3. Organic Pollutants Analysis
2.4. Enzymatic Activities
2.4.1. Glucose 6-Phosphate Dehydrogenase (G6PDH) & Cytochrome C Oxidase (CCO)
- -
- Glucose 6-phosphate dehydrogenase (G6PDH; EC 1.1.1.49, Gauthier et al. 2008): Imidazole buffer ((imidazole 50 mM, MgCl2 25 mM, KCl 70 mM), pH 7.5, β-nicotinamide adenine dinucleotide phosphate (NADP) 0.3 mM, glucose-6-phosphate 200 mM (omitted in controls). G6PDH activity was undetected in muscle samples and was only measured in liver at 340 nm.
- -
2.4.2. Ethoxyresorufin-O-Deethylase (EROD) & Acetylcholinesterase (AChE)
2.5. Lipids Analysis
2.6. Genetic Diversity
2.6.1. Genotyping
2.6.2. Analysis of Genetic Diversity
2.7. Statistical Analyses
3. Results
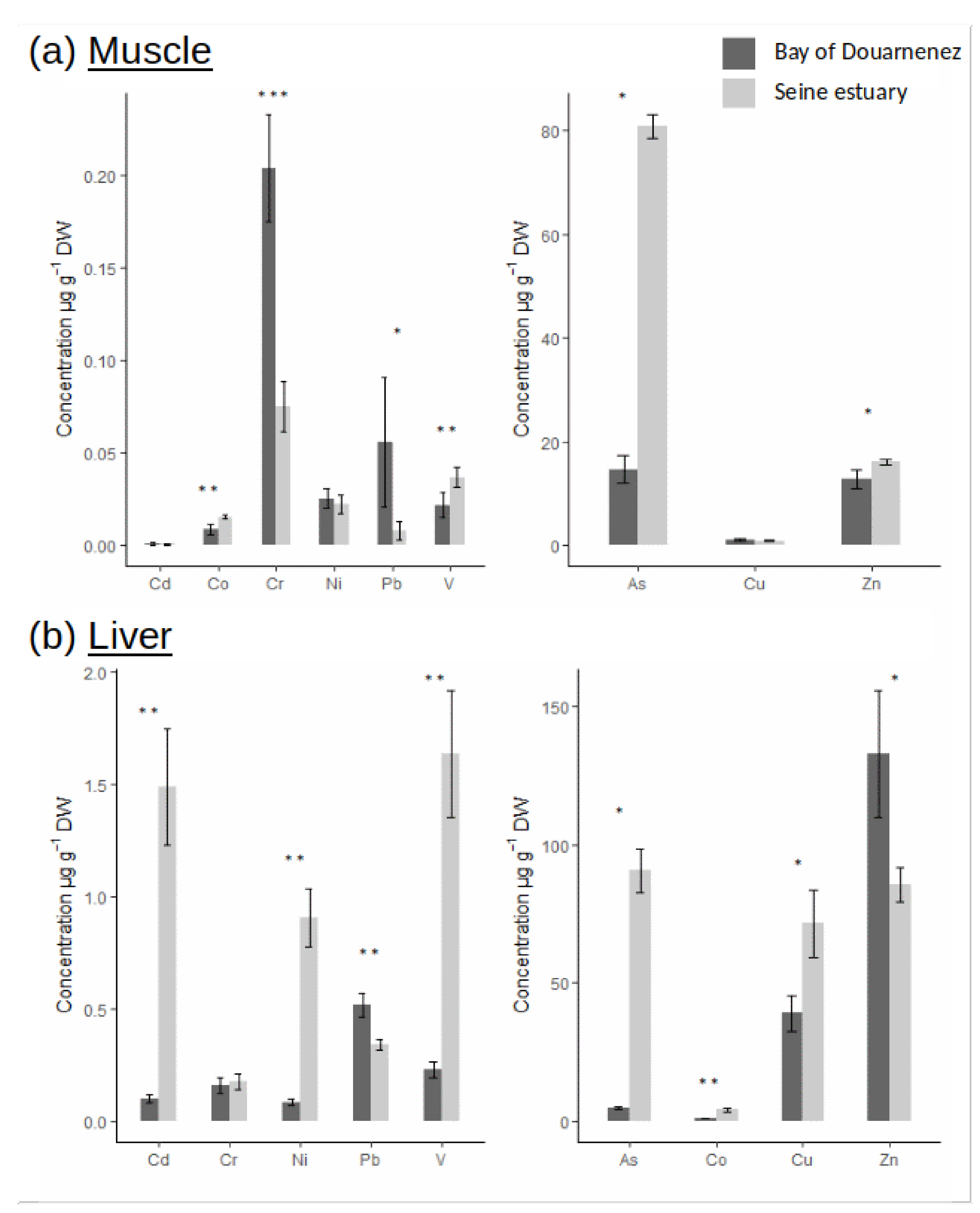
3.1. Contaminant Concentrations
3.2. Fish Biometry, Condition Index, Hepato-Somatic Index, Muscle Protein Concentration
3.3. G6PDH, CCO, AChE and EROD Activities
3.4. Muscle Lipids
3.5. Integration of the Phenotypic Responses
3.6. Genetic Variability
4. Discussion
4.1. Chemical Contamination & Hypothesis on the Flounder Use of Habitat in the Seine Estuary and Douarnenez Bay
4.2. Condition Index, Muscle Protein Content, Antioxidant Defences and Bioenergetics
4.3. Neurotoxicity and Xenobiotic Metabolism
4.4. Storage Lipids and Membrane Lipids in Muscle
4.5. Potential Relationships between Phospholipids, Xenobiotic Metabolism, Antioxidant Defense and Bioenergetics in Contrasted Environments
4.6. Genetic Variability in the Two Populations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wolanski, E.; Elliott, M. Estuarine Ecohydrology: An Introduction; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Robins, P.E.; Skov, M.W.; Lewis, M.; Giménez, L.; Davies, A.G.; Malham, S.K.; Neill, S.P.; E McDonald, J.; Whitton, T.A.; Jackson, S.E.; et al. Impact of climate change on UK estuaries: A review of past trends and potential projections. Estuar. Coast. Shelf Sci. 2016, 169, 119–135. [Google Scholar] [CrossRef] [Green Version]
- Gunderson, A.R.; King, E.E.; Boyer, K.; Tsukimura, B.; Stillman, J.H. Species as Stressors: Heterospecific Interactions and the Cellular Stress Response under Global Change. Integr. Comp. Biol. 2017, 57, 90–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallett, C.S.; Hobday, A.J.; Tweedley, J.R.; Thompson, P.A.; McMahon, K.; Valesini, F.J. Observed and predicted impacts of climate change on the estuaries of south-western Australia, a Mediterranean climate region. Reg. Environ. Chang. 2017, 18, 1357–1373. [Google Scholar] [CrossRef]
- Sinha, E.; Michalak, A.M.; Balaji, V. Eutrophication will increase during the 21st century as a result of precipitation changes. Science 2017, 357, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hylland, K.; Robinson, C.D.; Burgeot, T.; Martínez-Gómez, C.; Lang, T.; Svavarsson, J.; Thain, J.E.; Vethaak, A.D.; Gubbins, M.J. Integrated chemical and biological assessment of contaminant impacts in selected European coastal and offshore marine areas. Mar. Environ. Res. 2017, 124, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vethaak, A.D.; Davies, I.M.; Thain, J.E.; Gubbins, M.J.; Martínez-Gómez, C.; Robinson, C.D.; Moffat, C.F.; Burgeot, T.; Maes, T.; Wosniok, W.; et al. Integrated indicator framework and methodology for monitoring and assessment of hazardous substances and their effects in the marine environment. Mar. Environ. Res. 2017, 124, 11–20. [Google Scholar] [CrossRef] [Green Version]
- MSFD 2008. Directive 2008/56/EC of the European Parliament and the Council of 17 June 2008 Establishing a Framework for Community Action in the Field of Marine Environmental Policy. Available online: http://ec.europa.eu/environment/water/marine/index_en.htm (accessed on 10 September 2020).
- Dando, P. Site fidelity, homing and spawning migrations of flounder Platichthys flesus in the Tamar estuary, South West England. Mar. Ecol. Prog. Ser. 2011, 430, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Marchand, J.; Quiniou, L.; Riso, R.; Thebaut, M.-T.; Laroche, J. Physiological cost of tolerance to toxicants in the European flounder Platichthys flesus, along the French Atlantic Coast. Aquat. Toxicol. 2004, 70, 327–343. [Google Scholar] [CrossRef]
- Falciani, F.; Diab, A.; Sabine, V.; Williams, T.D.; Ortega, F.; George, S.G.; Chipman, J. Hepatic transcriptomic profiles of European flounder (Platichthys flesus) from field sites and computational approaches to predict site from stress gene responses following exposure to model toxicants. Aquat. Toxicol. 2008, 90, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laroche, J.; Gauthier, O.; Quiniou, L.; Devaux, A.; Bony, S.; Evrard, E.; Cachot, J.; Cherel, Y.; Larcher, T.; Riso, R.; et al. Variation patterns in individual fish responses to chemical stress among estuaries, seasons and genders: The case of the European flounder (Platichthys flesus) in the Bay of Biscay. Environ. Sci. Pollut. Res. 2012, 20, 738–748. [Google Scholar] [CrossRef]
- Dupuy, C.; Galland, C.; Pichereau, V.; Sanchez, W.; Riso, R.; LaBonne, M.; Amara, R.; Charrier, G.; Fournier, M.; Laroche, J. Assessment of the European flounder responses to chemical stress in the English Channel, considering biomarkers and life history traits. Mar. Pollut. Bull. 2015, 95, 634–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borcier, E.; Charrier, G.; Amérand, A.; Théron, M.; Loizeau, V.; Pédron, N.; Laroche, J. Bioenergetic transcriptomic responses of European flounder (Platichthys flesus) populations in contrasted environments: Impacts of pollution and global warming. J. Xenobiotics 2016, 6. [Google Scholar] [CrossRef]
- Capela, R.; Raimundo, J.; Santos, M.M.; Caetano, C.; Vale, C.; Guimaraes, L.; Reis-Henriques, M.A. The use of biomarkers as integrative tools for transitional water bodies monitoring in the Water Framework Directive context—A holistic approcah in Minho river transitional waters. Sci. Total Environ. 2016, 539, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Burgeot, T.; Akcha, F.; Ménard, D.; Robinson, C.D.; Loizeau, V.; Brach-Papa, C.; Martínez-Gómez, C.; Le Goff, J.; Budzinski, H.; Le Menach, K.; et al. Integrated monitoring of chemicals and their effects on four sentinel species, Limanda limanda, Platichthys flesus, Nucella lapillus and Mytilus sp., in Seine Bay: A key step towards applying biological effects to monitoring. Mar. Environ. Res. 2017, 124, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Dabrowska, H.; Kopko, O.; Lehtonen, K.K.; Lang, T.; Waszak, I.; Balode, M.; Strode, E. An integrated assessment of pollution and biological effects in flounder, mussels and sediment in the southern Baltic Sea coastal area. Environ. Sci. Pollut. Res. 2016, 24, 3626–3639. [Google Scholar] [CrossRef]
- Galland, C.; Dupuy, C.; Capitaine, C.; Auffret, M.; Quiniou, L.; Laroche, J.; Pichereau, V. Comparisons of liver proteomes in the European flounder Platichthys flesus from three contrasted estuaries. J. Sea Res. 2013, 75, 135–141. [Google Scholar] [CrossRef]
- Peédron, N.; Artigaud, S.; Infante, J.-L.Z.; Le Bayon, N.; Charrier, G.; Pichereau, V.; Laroche, J. Proteomic responses of European flounder to temperature and hypoxia as interacting stressors: Differential sensitivities of populations. Sci. Total. Environ. 2017, 586, 890–899. [Google Scholar] [CrossRef]
- Pédron, N.; Le Du, J.; Charrier, G.; Zambonino-Infante, J.-L.; Le Bayon, N.; Vasconcelos, R.P.; Fonseca, V.F.; Le Grand, F.; Laroche, J. Contrasting patterns of energy metabolism in northern vs southern peripheral European flounder populations exposed to temperature rising and hypoxia. Mar. Environ. Res. 2017, 129, 258–267. [Google Scholar] [CrossRef]
- Lavergne, E.; Pédron, N.; Calves, I.; Claireaux, G.; Mazurais, D.; Zambonino-Infante, J.-L.; Le Bayon, N.; Cahu, C.; Laroche, J. Does the chronic chemical contamination of a European flounder population decrease its thermal tolerance? Mar. Pollut. Bull. 2015, 95, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Marchand, J.; Evrard, E.; Guinand, B.; Cachot, J.; Quiniou, L.; Laroche, J. Genetic polymorphism and its potential relation to environmental stress in five populations of the European flounder Platichthys flesus, along the French Atlantic coast. Mar. Environ. Res. 2010, 70, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Marchand, J.; Quiniou, L.; Laroche, J. Relationships between genotypes and phenotypes in natural populations of the European flounder (Platichthys flesus) under different types of contamination in estuaries. J. Xenobiotics 2013, 3, e14. [Google Scholar] [CrossRef]
- Calvès, I.; Lavergne, E.; Meistertzheim, A.-L.; Charrier, G.; Cabral, H.; Guinand, B.; Quiniou, L.; Laroche, J. Genetic structure of European flounder Platichthys flesus: Effects of both the southern limit of the species’ range and chemical stress. Mar. Ecol. Prog. Ser. 2013, 472, 257–273. [Google Scholar] [CrossRef] [Green Version]
- Chiffoleau, J.F. La Contamination Chimique sur le Littoral Loire-Bretagne. Résultats de 35 Années de Suivi du Réseau d’Observation de la Contamination Chimique. Ifremer, Nantes, France. Département Ressources Biologiques et Environnement; Unité Biogéochimie et Ecotoxicologie: Nantes, France, October 2017; pp. 1–72, RST.RBE-BE/2017.02. [Google Scholar]
- Wang, Y.; Salmon, A.B.; Harshman, L.G. A cost of reproduction: Oxidative stress susceptibility is associated with increased egg production in Drosophila melanogaster. Exp. Gerontol. 2001, 36, 1349–1359. [Google Scholar] [CrossRef]
- Wiersma, P.; Selman, C.; Speakman, J.R.; Verhulst, S. Birds sacrifice oxidative protection for reproduction. Proc. R. Soc. B: Boil. Sci. 2004, 271, S360–S363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopko, O.; Dabrowska, H. Variability of biological indices, biomarkers, and organochlorine contaminants in flounder (Platichthys flesus) in the Gulf of Gdańsk, southern Baltic Sea. Chemosphere 2018, 194, 701–713. [Google Scholar] [CrossRef]
- O’Leary, S.J.; Hice, L.A.; Feldheim, K.A.; Frisk, M.G.; McElroy, A.E.; Fast, M.D.; Chapman, D.D. Severe Inbreeding and Small Effective Number of Breeders in a Formerly Abundant Marine Fish. PLoS ONE 2013, 8, e66126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, P.B.; Cowx, I.G.; Oleksiak, M.F.; Griffiths, A.M.; Grahn, M.; Stevens, J.R.; Carvalho, G.R.; Nicol, E.; Tyler, C.R. Population-level consequences for wild fish exposed to sublethal concentrations of chemicals—A critical review. Fish Fish. 2016, 17, 545–566. [Google Scholar] [CrossRef]
- FDPPMA29. Evaluation de l’état de la population d’anguille européenne sur le bassin versant de la baie de Douarnenez (Finistère) en 2012. Contrat de Projet Etat-Région Bretagne 2007–2013; Fédération Départementale du Finistère pour la Pêche et la Protection du Milieu Aquatique: Quimper, France, 2012; pp. 1–37. [Google Scholar]
- Poisson, E. Effets de la Contamination Chimique: Des Organismes en danger? Fascicule Seine-Aval 2; Groupement d’Intérêt Publique Seine-Aval: Rouen, France, 2011; pp. 1–69. [Google Scholar]
- Peig, J.; Green, A.J. The paradigm of body condition: A critical reappraisal of current methods based on mass and length. Funct. Ecol. 2010, 24, 1323–1332. [Google Scholar] [CrossRef]
- Lambert, Y.; Dutil, J.-D. Can simple condition indices be used to monitor and quantify seasonal changes in the energy reserves of cod (Gadus morhua)? Can. J. Fish. Aquat. Sci. 1997, 54, 104–112. [Google Scholar] [CrossRef]
- Lacroix, C.; Le Cuff, N.; Receveur, J.; Moraga, D.; Auffret, M.; Guyomarch, J. Development of an innovative and “green” stir bar sorptive extraction–thermal desorption–gas chromatography–tandem mass spectrometry method for quantification of polycyclic aromatic hydrocarbons in marine biota. J. Chromatogr. A 2014, 1349, 1–10. [Google Scholar] [CrossRef]
- Shrivastava, A.; Gupta, V.B. Methods for the determination of limit of detection and limit of quantitation of the analytical methods. Chron. Young- Sci. 2011, 2, 21. [Google Scholar] [CrossRef]
- Gauthier, C.; Campbell, P.G.; Couture, P. Physiological correlates of growth and condition in the yellow perch (Perca flavescens). Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2008, 151, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.D.; Mayer, R.T. Ethoxyresorufin: Direct fluorimetric assay of a microsomal O-dealkylation which is preferentially inducible by 3-methylcholanthrene. Drug Metab. Dispos. 1974, 2, 583–588. [Google Scholar]
- Ellman, G.L.; Courtney, K.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Mathieu-Resuge, M.; Kraffe, E.; Le Grand, F.; Boens, A.; Bideau, A.; Lluch-Cota, S.E.; Racotta, I.S.; Schaal, G. Trophic ecology of suspension-feeding bivalves inhabiting a north-eastern Pacific coastal lagoon: Comparison of different biomarkers. Mar. Environ. Res. 2019, 145, 155–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amara, R.; Meziane, T.; Gilliers, C.; Hermel, G.; Laffargue, P. Growth and condition indices in juvenile sole (Solea solea L.) measured to assess the quality of essential fish habitat. Mar. Ecol. Prog. Ser. 2007, 351, 209–220. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual (No. Ed. 2); Cold Spring Harbor Laboratory Press: Suffolk, NY, USA, 1989. [Google Scholar]
- Dixon, T.J. Molecular Genetic Studies of Pollutant Response in the European Flounder, Platichthys flesus. Ph.D. Thesis, University of Stirling, Stirling, UK, 2003. [Google Scholar]
- Casas, L.; Martínez, P.; Sánchez, L. Characterization of microsatellite markers derived from sequence databases for the European flounder (Platichthys flesus). Mol. Ecol. Notes 2005, 5, 664–665. [Google Scholar] [CrossRef]
- Hemmer-Hansen, J.; Nielsen, E.E.; Grønkjaer, P.; Loeschcke, V.; Grønkjær, P. Evolutionary mechanisms shaping the genetic population structure of marine fishes; lessons from the European flounder (Platichthys flesus L.). Mol. Ecol. 2007, 16, 3104–3118. [Google Scholar] [CrossRef] [PubMed]
- Florin, A.-B.; Höglund, J. Population structure of flounder (Platichthys flesus) in the Baltic Sea: Differences among demersal and pelagic spawners. Heredity 2008, 101, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Pédron, N.; Morvezen, R.; Le Moan, A.; Guinand, B.; Zambonino-Infante, J.L.; Laroche, J.; Charrier, G. New set of candidate gene SNPs and microsatellites to disentangle selective and neutral processes shaping population responses of European flounder (Platichthys flesus) to anthropogenic stress and contrasted environments. Conserv. Genet. Resour. 2015, 7, 823–826. [Google Scholar] [CrossRef]
- Coombs, J.A.; Letcher, B.H.; Nislow, K.H. Create: A software to create input files from diploid genotypic data for 52 genetic software programs. Mol. Ecol. Resour. 2008, 8, 578–580. [Google Scholar] [CrossRef]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX, Software under WindowsTM for the Genetic of Populations; Laboratory Genome, Populations, Interactions CNRS UMR, 5000: Montpellier, France, 2001. [Google Scholar]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: AnRPackage for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Henry, F.; Amara, R.; Courcot, L.; Lacouture, D.; Bertho, M.-L. Heavy metals in four fish species from the French coast of the Eastern English Channel and Southern Bight of the North Sea. Environ. Int. 2004, 30, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Evrard, E.; Devaux, A.; Bony, S.; Burgeot, T.; Riso, R.; Budzinski, H.; Le Du, M.; Quiniou, L.; Laroche, J. Responses of the European flounder Platichthys flesus to the chemical stress in estuaries: Load of contaminants, gene expression, cellular impact and growth rate. Biomarkers 2009, 15, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.D.; Turan, N.; Diab, A.M.; Wu, H.; MacKenzie, C.; Bartie, K.L.; Hrydziuszko, O.; Lyons, B.P.; Stentiford, G.D.; Herbert, J.M.; et al. Towards a System Level Understanding of Non-Model Organisms Sampled from the Environment: A Network Biology Approach. PLoS Comput. Biol. 2011, 7, e1002126. [Google Scholar] [CrossRef]
- Mazéas, O. Evaluation de L’exposition des Organismes aux HAP dans le Milieu Marin par le Dosage des Metabolites de HAP—Cas de la Marée Noire ERIKA. Ph.D Thesis, Université de Bordeaux I, Talence, France, 2004. [Google Scholar]
- Déniel, C. Les Poissons Plats (Téléostéens, Pleuronectiformes) en baie de Douarnenez: Reproduction, Croissance et Migration des Bothidae, Scophthalmidae, Pleuronectidae et Soleidae. Ecologie, Environnement. Ph.D. Thesis, Université de Bretagne Occidentale, Brest, France, 1981. [Google Scholar]
- Gallien-Landriau, I. Etude de L’altération Fonctionnelle du Système Reproducteur par les Perturbateurs Endocriniens. Caractérisation des Effets, Identification des Xéno-Estrogènes Impliqués et Conséquences sur les Populations de Poissons en Estuaire et Baie de Seine. Ph.D. Thesis, Université du Havre, Le Havre, France, 2003; 151p. [Google Scholar]
- Rui, L. Energy Metabolism in the Liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar]
- Lushchak, V.I.; Bagnyukova, T.V. Effects of different environmental oxygen levels on free radical processes in fish. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2006, 144, 283–289. [Google Scholar] [CrossRef]
- Koehler, A. The gender-specific risk to liver toxicity and cancer of flounder (Platichthys flesus (L.)) at the German Wadden Sea coast. Aquat. Toxicol. 2004, 70, 257–276. [Google Scholar] [CrossRef]
- Grasset, J.; Bougas, B.; Campbell, P.G.; Bernatchez, L.; Couture, P. Temperature, oxygen, and diet modulate gene transcription and metabolic capacities in yellow perch. Can. J. Fish. Aquat. Sci. 2014, 71, 1635–1641. [Google Scholar] [CrossRef]
- Van Noorden, C.J.F.; Bahns, S.; Köhler, A. Adaptational changes in kinetic parameters of G6PDH but not PGDH during contamination-induced carcinogenesis in livers of North Sea flatfish. Biochim. Biophys. Acta 1997, 1342, 141–148. [Google Scholar] [CrossRef]
- Bagnyukova, T.V.; Chahrak, O.I.; Lushchak, V.I. Coordinated response of golfish antioxidant defenses to environmental stress. Aquat. Tox. 2006, 78, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Salin, K.; Auer, S.K.; Rudolf, A.M.; Anderson, G.J.; Selman, C.; Metcalfe, N.B. Variation in Metabolic Rate among Individuals Is Related to Tissue-Specific Differences in Mitochondrial Leak Respiration. Physiol. Biochem. Zool. 2016, 89, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Crawford, D.L.; Nacci, D.E.; Oleksiak, M.F. Heritable oxidative phosphorylation differences in a pollutant resistant Fundulus heteroclitus population. Aquat. Toxicol. 2016, 177, 44–50. [Google Scholar] [CrossRef]
- Escartin, E.; Porte, C. The use of cholinesterase and carboxylesterase activities from Mytilus galloprovincialis in pollution monitoring. Environ. Toxicol. Chem. 1997, 16, 2090–2095. [Google Scholar] [CrossRef]
- Payne, J.; Mathieu, A.; Melvin, W.; Fancey, L. Acetylcholinesterase, an old biomarker with a new future? Field trials in association with two urban rivers and a paper mill in Newfoundland. Mar. Pollut. Bull. 1996, 32, 225–231. [Google Scholar] [CrossRef]
- Guilhermino, L.; Barros, P.; Silva, M.C.; Soares, A.M.V.M. SHORT COMMUNICATION Should the use of inhibition of cholinesterases as a specific biomarker for organophosphate and carbamate pesticides be questioned. Biomarkers 1998, 3, 157–163. [Google Scholar] [CrossRef]
- Kopecka, J.; Lehtonen, K.K.; Baršienė, J.; Broeg, K.; Vuorinen, P.J.; Gercken, J.; Pempkowiak, J. Measurements of biomarker levels in flounder (Platichthys flesus) and blue mussel (Mytilus trossulus) from the Gulf of Gdańsk (southern Baltic). Mar. Pollut. Bull. 2006, 53, 406–421. [Google Scholar] [CrossRef] [Green Version]
- De La Torre, F.; Ferrari, L.; Salibián, A. Freshwater pollution biomarker: Response of brain acetylcholinesterase activity in two fish species. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2002, 131, 271–280. [Google Scholar] [CrossRef]
- Suresh, A.; Sivaramakrishna, B.; Victoriamma, P.C.; Radhakrishnaiah, K. Comparative study of inhibition of acetylcholinesteraseactivity in the freshwater fish Cyprinus carpio by mercury and zinc. Biochem. Int. 1992, 26, 367–375. [Google Scholar] [PubMed]
- Labrot, F.; Ribera, D.; Denis, M.S.; Narbonne, J.F. In vitro and in vivo studies of potential biomarkers of lead and uranium contamination: Lipid peroxidation, acetylcholinesterase, catalase and glutathione peroxidase activities in three non-mammalian species. Biomarkers 1996, 1, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Van Der Oost, R.; Beyer, J.; Vermeulen, N.P. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Arukwe, A.; Goksoyr, A. Changes in three hepatic cytochrome P450 subfamilies during a reproductive cycle in turbot (Scophthalmus maximus L.). J. Exp. Zool. 1997, 277, 313–325. [Google Scholar] [CrossRef]
- Brammell, B.F.; Price, D.J.; Birge, W.J.; Elskus, A.A. Lack of CYP1A responsiveness in species inhabiting chronically contaminated habitats: Two varieties of resistance? Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2013, 157, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Roush, R.T.; Mc Kenzie, J.A. Ecological genetics of insecticide and acaricide resistance. Unkn. J. 1987, 32, 361–380. [Google Scholar] [CrossRef]
- Kerambrun, E.; Henry, F.; Cornille, V.; Courcot, L.; Amara, R. A combined measurement of metal bioaccumulation and condition indices in juvenile European flounder, Platichthys flesus, from European estuaries. Chemosphere 2013, 91, 498–505. [Google Scholar] [CrossRef]
- Valencak, T.G.; Azzu, V. Making heads or tails of mitochondrial membranes in longevity and aging: A role for comparative studies. Longev. Heal. 2014, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Hulbert, A.; Pamplona, R.; Buffenstein, R.; Buttemer, W.A. Life and Death: Metabolic Rate, Membrane Composition, and Life Span of Animals. Physiol. Rev. 2007, 87, 1175–1213. [Google Scholar] [CrossRef]
- Brignac-Huber, L.M.; Park, J.W.; Reed, J.R.; Backes, W.L. Cytochrome P450 Organization and Function Are Modulated by Endoplasmic Reticulum Phospholipid Heterogeneity. Drug Metab. Dispos. 2016, 44, 1859–1866. [Google Scholar] [CrossRef]
- Ghosh, M.C.; Ray, A.K. Membrane Phospholipid Augments Cytochrome P4501a Enzymatic Activity by Modulating Structural Conformation during Detoxification of Xenobiotics. PLoS ONE 2013, 8, e57919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajiwara, K.; Muneoka, T.; Watanabe, Y.; Karashima, T.; Kitagaki, H.; Funato, K. Perturbation of shingolipid metabolism induces endoplasmic reticulum stress-mediated mitochondrial apoptosis in budding yeast. Mol. Microbiol. 2012, 86, 1246–1261. [Google Scholar] [CrossRef] [PubMed]
- Crawford, D.L.; Oleksiak, M.F. The biological importance of measuring individual variation. J. Exp. Biol. 2007, 210, 1613–1621. [Google Scholar] [CrossRef] [Green Version]
- Norin, T.; Malte, H.; Clark, T.D. Differential plasticity of metabolic rate phenotypes in a tropical fish facing environmental change. Funct. Ecol. 2015, 30, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Author Correction: Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Vazquez, E.J.; Moghaddas, S.; Hoppel, C.L.; Lesnefsky, E.J. Production of Reactive Oxygen Species by Mitochondria. J. Biol. Chem. 2003, 278, 36027–36031. [Google Scholar] [CrossRef] [Green Version]
- Achard-Joris, M.; Gonzalez, P.; Marie, V.; Baudrimont, M.; Bourdineaud, J.-P. Cytochrome c Oxydase Subunit I Gene is Up-regulated by Cadmium in Freshwater and Marine Bivalves. BioMetals 2006, 19, 237–244. [Google Scholar] [CrossRef]
- Fonovich, T.M.; Perez-Coll, C.S.; Fridman, O.; D’Eramo, J.L.; Herkovits, J. Phospholipid changes in Rhinella arenarum embryos under different acclimation conditions to copper. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 189, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.Y.; Wang, W.-X. A lipidomic approach to understand copper resilience in oyster Crassostrea hongkongensis. Aquat. Toxicol. 2018, 204, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Marchand, J.; Tanguy, A.; Laroche, J.; Quiniou, L.; Moraga, D. Responses of European flounder Platichthys flesus populations to contamination in different estuaries along the Atlantic coast of France. Mar. Ecol. Prog. Ser. 2003, 260, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Jayasundara, N.; Fernando, P.W.; Osterberg, J.S.; Cammen, K.M.; Schultz, T.F.; Di Giulio, R.T. Cost of Tolerance: Physiological Consequences of Evolved Resistance to Inhabit a Polluted Environment in Teleost Fish Fundulus heteroclitus. Environ. Sci. Technol. 2017, 51, 8763–8772. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DORM-4 | DOLT-5 | TORT-3 | ||||
|---|---|---|---|---|---|---|
| Measured (n = 5) | Certified | Measured (n = 5) | Certified | Measured (n = 5) | Certified | |
| As | 7.6 ± 0.7 | 6.9 ± 0.5 | 32 ± 6 | 35 ± 6 | 59 ± 11 | 60 ± 4 |
| Cd | 0.31 ± 0.04 | 0.30 ± 0.02 | 12 ± 2 | 14.5 ± 0.6 | 34 ± 5 | 42 ± 2 |
| Co | 0.28 ± 0.03 | 0.25 * | 0.29 ± 0.09 | 0.27 ± 0.03 | 1.04 ± 0.06 | 1.06 * |
| Cr | 1.5 ± 0.1 | 1.9 ± 0.2 | 1.0 ± 0.2 | 2.3 ± 0.6 | 0.9 ± 0.1 | 1.9 ± 0.3 |
| Cu | 16.1 ± 0.9 | 15.7 ± 0.5 | 31 ± 4 | 35 ± 3 | 350 ± 80 | 500 ± 30 |
| Ni | 1.20 ± 0.08 | 1.3 ± 0.2 | 0.7 ± 0.2 | 1.7 ± 0.7 | 4.2 ± 0.4 | 5.3 ± 0.3 |
| Pb | 0.30 ± 0.09 | 0.42 ± 0.06 | 0.04 ± 0.03 | 0.16 ± 0.04 | 0.26 ± 0.07 | 0.23 ± 0.02 |
| V | 1.5 ± 0.1 | 1.6 ± 0.2 | 0.50 ± 0.04 | 0.51 ± 0.06 | 8.8 ± 0.3 | 9.1 ± 0.4 |
| Zn | 49 ± 7 | 52 ± 3 | 96 ± 14 | 105 ± 6 | 125 ± 9 | 136 ± 6 |
| ng.g−1 DW | LS | LD | MS | MD |
|---|---|---|---|---|
| Naphthalene | 97.26 | - | - | - |
| PCB 101 | 91.32 | - | 70.12 | - |
| PCB 118 | 57.56 | 24.96 | 69.85 | - |
| PCB 153 | 255.20 | - | 164.93 | - |
| PCB 138 | 290.65 | 45.49 | 167.34 | - |
| PCB 180 | 26.20 | - | - | - |
| Location | DZ | SE | |||
|---|---|---|---|---|---|
| mean | sd | sign. | mean | sd | |
| Storage lipids | |||||
| ST EST | 0.02 | 0.03 | *** | 0.06 | 0.05 |
| GL ETH | 0.01 | 0.02 | NS | 0.02 | 0.03 |
| TG | 1.96 | 1.82 | ** | 1.01 | 1.21 |
| FFA | 0.05 | 0.05 | NS | 0.04 | 0.03 |
| ALC | 0.04 | 0.12 | NS | 0.02 | 0.03 |
| Membrane lipids | |||||
| FST | 0.40 | 0.09 | ** | 0.45 | 0.10 |
| SPG | 0.20 | 0.12 | *** | 0.35 | 0.10 |
| LPC | 0.19 | 0.08 | *** | 0.10 | 0.08 |
| PC | 5.40 | 0.71 | *** | 4.42 | 0.68 |
| PS | 0.27 | 0.05 | ** | 0.32 | 0.10 |
| PI | 0.87 | 0.15 | *** | 0.64 | 0.12 |
| CL | 0.33 | 0.15 | NS | 0.27 | 0.12 |
| PE | 2.47 | 0.39 | *** | 2.17 | 0.31 |
| ∑ SL | 2.08 | ** | 1.15 | ||
| ∑ ML | 10.14 | ** | 8.73 | ||
| ∑ ALL | 12.21 | ** | 9.88 | ||
| Locus | Panels | Douarnenez | Seine | ||||
|---|---|---|---|---|---|---|---|
| Na | Ho | He | Na | Ho | He | ||
| Nplaf_8 | 1 | 4 | 0.444 | 0.448 | 4 | 0pl.300 | 0.339 |
| Nplaf_14 | 3 | 3 | 0.378 | 0.413 | 4 | 0.352 | 0.342 |
| Nplaf_15 | 2 | 5 | 0.272 | 0.338 | 5 | 0.718 | 0.569 |
| Nplaf_23 | 2 | 5 | 0.500 | 0.637 | 6 | 0.419 | 0.580 |
| Nplaf_24 | 3 | 6 | 0.588 | 0.645 | 6 | 0.656 | 0.664 |
| Nplaf_25 | 3 | 6 | 0.756 | 0.673 | 4 | 0.685 | 0.658 |
| Nplaf_28 | 3 | 7 | 0.620 | 0.728 | 6 | 0.650 | 0.710 |
| Nplaf_30 | 3 | 4 | 0.166 | 0.205 | 4 | 0.437 | 0.371 |
| Nplaf_35 | 1 | 17 | 0.971 | 0.917 | 19 | 0.903 | 0.915 |
| PFUSC3 | 1 | 6 | 0.529 | 0.478 | 3 | 0.382 | 0.349 |
| PFUSC4 | 3 | 3 | 0.527 | 0.526 | 6 | 0.666 | 0.568 |
| PFUSC7 | 2 | 10 | 0.617 | 0.742 | 7 | 0.709 | 0.721 |
| PFUSC8 | 1 | 4 | 0.450 | 0.502 | 3 | 0.428 | 0.438 |
| PL142 | 2 | 14 | 0.896 | 0.878 | 11 | 0.619 | 0.833 |
| StPfl001 | 1 | 21 | 0.742 | 0.832 | 18 | 0.774 | 0.868 |
| StPfl002 | 3 | 10 | 0.638 | 0.649 | 6 | 0.558 | 0.626 |
| StPfl003 | 2 | 5 | 0.756 | 0.679 | 4 | 0.454 | 0.543 |
| StPfl005 | 2 | 3 | 0.457 | 0.452 | 4 | 0.457 | 0.530 |
| StPfl015 | 1 | 9 | 0.432 | 0.747 | 9 | 0.363 | 0.604 |
| StPfl025 | 2 | 7 | 0.500 | 0.551 | 7 | 0.687 | 0.617 |
| Multilocus | 7.45 | 0.562 | 0.602 | 6.8 | 0.561 | 0.592 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borcier, E.; Charrier, G.; Couteau, J.; Maillet, G.; Le Grand, F.; Bideau, A.; Waeles, M.; Le Floch, S.; Amara, R.; Pichereau, V.; et al. An Integrated Biomarker Approach Using Flounder to Improve Chemical Risk Assessments in the Heavily Polluted Seine Estuary. J. Xenobiot. 2020, 10, 14-35. https://doi.org/10.3390/jox10020004
Borcier E, Charrier G, Couteau J, Maillet G, Le Grand F, Bideau A, Waeles M, Le Floch S, Amara R, Pichereau V, et al. An Integrated Biomarker Approach Using Flounder to Improve Chemical Risk Assessments in the Heavily Polluted Seine Estuary. Journal of Xenobiotics. 2020; 10(2):14-35. https://doi.org/10.3390/jox10020004
Chicago/Turabian StyleBorcier, Elodie, Grégory Charrier, Jérôme Couteau, Géraldine Maillet, Fabienne Le Grand, Antoine Bideau, Matthieu Waeles, Stéphane Le Floch, Rachid Amara, Vianney Pichereau, and et al. 2020. "An Integrated Biomarker Approach Using Flounder to Improve Chemical Risk Assessments in the Heavily Polluted Seine Estuary" Journal of Xenobiotics 10, no. 2: 14-35. https://doi.org/10.3390/jox10020004
APA StyleBorcier, E., Charrier, G., Couteau, J., Maillet, G., Le Grand, F., Bideau, A., Waeles, M., Le Floch, S., Amara, R., Pichereau, V., & Laroche, J. (2020). An Integrated Biomarker Approach Using Flounder to Improve Chemical Risk Assessments in the Heavily Polluted Seine Estuary. Journal of Xenobiotics, 10(2), 14-35. https://doi.org/10.3390/jox10020004