Characterization and Antimicrobial Activity of Alkaloid Extracts from Seeds of Different Genotypes of Lupinus spp.

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Sampling
2.2. Alkaloid Extraction
2.3. GC-MS Analysis
2.4. Alkaloid Quantification
2.5. Antimicrobial Activity
2.5.1. Microorganisms
2.5.2. Antibacterial and Antifungal Tests
2.6. Statistical Analysis
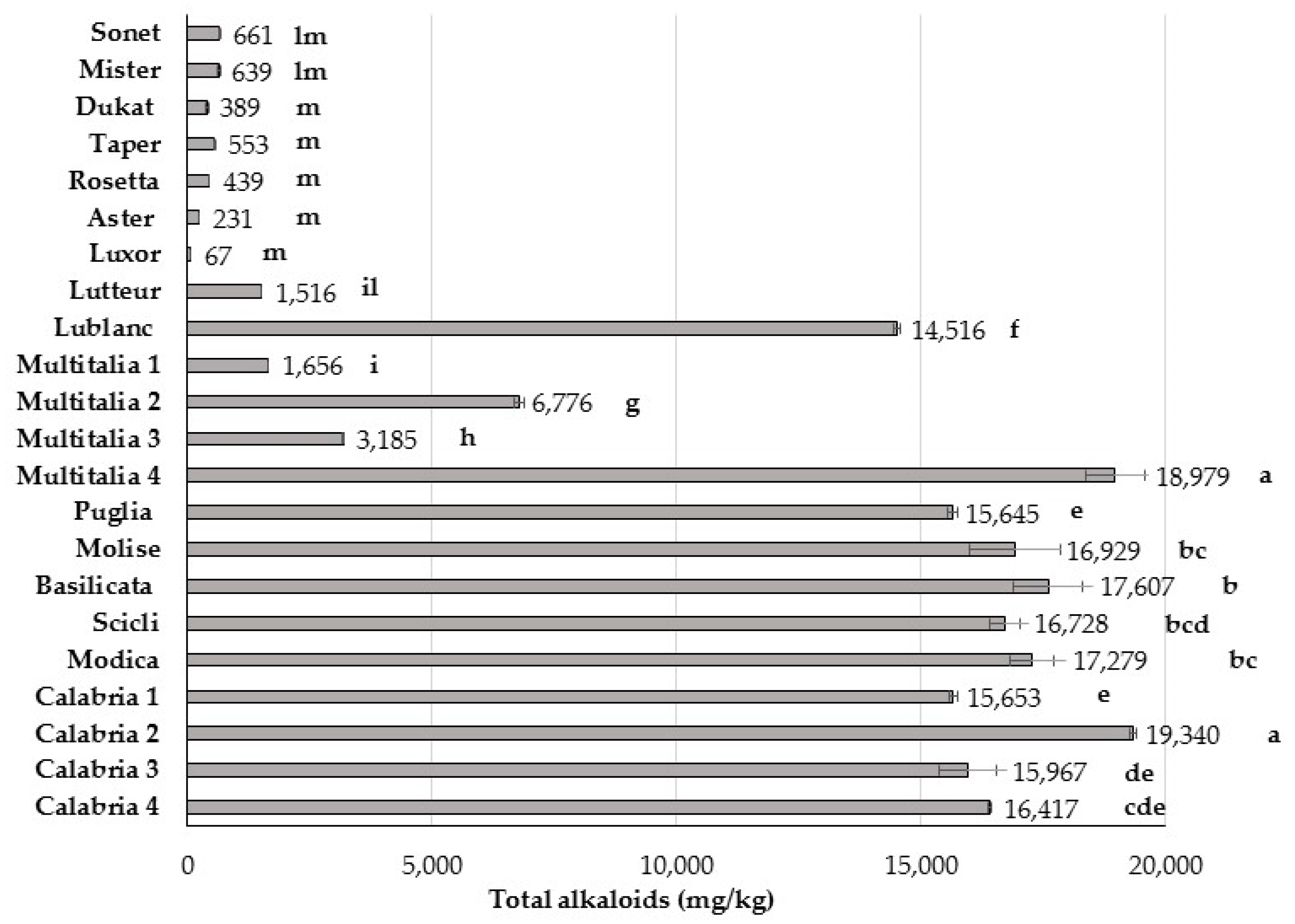
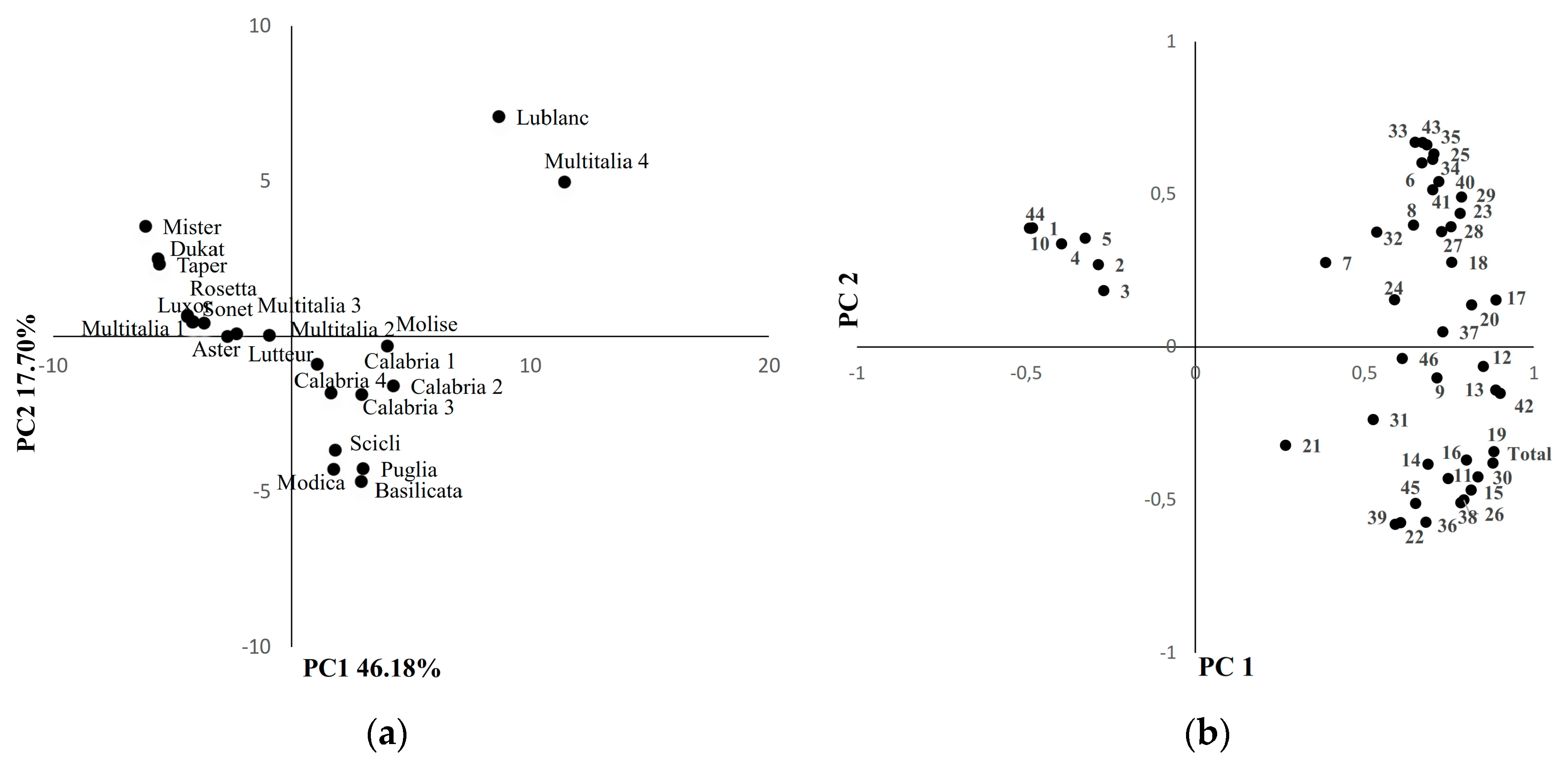
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gresta, F.; Wink, M.; Prins, U.; Abberton, M.; Capraro, J.; Scarafoni, A.; Hill, G. Lupins in European cropping systems. In Legumes in Cropping Systems; Murphy-Bokern, D., Stoddard, F., Watson, C., Eds.; CABI: Wallingford, Oxfordshire, UK, 2017; pp. 88–108. ISBN 9781780644981. [Google Scholar]
- Carvajal-Larenas, F.E.; Van Boekel, M.J.A.S.; Koziol, M.; Nout, M.J.R.; Linnemann, A.R. Effect of processing on the diffusion of alkaloids and quality of Lupinus mutabilis sweet. J. Food Process. Preserv. 2014, 38, 1461–1471. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Manunza, P.; Arnoldi, A.; Boschin, G. Quality of Lupinus albus L. (White lupin) seed: Extent of genotypic and environmental effects. J. Agric. Food Chem. 2014, 62, 6539–6545. [Google Scholar] [CrossRef] [PubMed]
- Gresta, F.; Abbate, V.; Avola, G.; Magazzù, G.; Chiofalo, B. Lupin seed for the crop-livestock food chain. Ital. J. Agron. 2010, 4, 333–340. [Google Scholar] [CrossRef]
- FAOSTAT. Statistics Database of the Food and Agriculture Organization of the United Nations. Food and Agriculture Organization of the United Nations: Rome. Available online: http://faostat3.fao.org/home/E (accessed on 18 September 2017).
- Aniszewski, T. Alkaloids: Chemistry, Biology, Ecology, and Applications, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2015; ISBN 9780444594334. [Google Scholar]
- Magalhães, S.C.; Fernandes, F.; Cabrita, A.R.; Fonseca, A.J.; Valentão, P.; Andrade, P.B. Alkaloids in the valorization of European Lupinus spp. seeds crop. Ind. Crops Prod. 2017, 95, 286–295. [Google Scholar] [CrossRef]
- Musco, N.; Cutrignelli, M.I.; Calabrò, S.; Tudisco, R.; Infascelli, F.; Grazioli, R.; Lo Presti, V.; Gresta, F.; Chiofalo, B. Comparison of nutritional and antinutritional traits among different species (Lupinus albus L.; Lupinus luteus L.; Lupinus angustifolius L.) and varieties of lupin seeds. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Muzquiz, M.; Cuadrado, C.; Ayet, G.; de la Cuadra, C.; Burbano, C.; Osagie, A. Variation of alkaloid components of Lupin seeds in 49 genotypes of Lupinus albus L. from different countries and locations. J. Agric. Food Chem. 1994, 42, 1447–1450. [Google Scholar] [CrossRef]
- Reinhard, H.; Rupp, H.; Sager, F.; Streule, M.; Zoller, O. Quinolizidine alkaloids and phomopsins in lupin seeds and lupin containing food. J. Chromatogr. A 2006, 1112, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Wink, M.; Meibner, C.; Witte, L. Pattern of quinolizidine alkaloids in 56 species of the genus Lupinus. Phytochemistry 1995, 38, 139–153. [Google Scholar] [CrossRef]
- Boschin, G.; Annicchiarico, P.; Resta, D.; D’Agostina, A.; Arnoldi, A. Quinolizidine Alkaloids in Seeds of Lupin Genotypes of Different Origins. J. Agric. Food Chem. 2008, 56, 3657–3663. [Google Scholar] [CrossRef] [PubMed]
- Resta, D.; Boschin, G.; D’Agostina, A.; Arnoldi, A. Evaluation of total quinolizidine alkaloids content in lupin flours, lupin-based ingredients, and foods. Mol. Nutr. Food Res. 2008, 52, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Canu Boido, C.; Tasso, B.; Boido, V.; Sparatore, F. Cytisine derivatives as ligands for neuronal nicotine receptors and with various pharmacological activities. Farmaco 2003, 58, 265–277. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, C.; Li, L.; Wang, Z.; Xiao, W.; Yang, Z. Hypoglycemic and hypolipidemic effects of oxymatrine in high-fat diet and streptozotocin-induced diabetic rats. Phytomedicine 2014, 21, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Huang, C.F.; Liu, X.S.; Jiang, J. In Vitro Anti-Tumour Activities of Quinolizidine Alkaloids Derived from Sophora flavescens Ait. Basic Clin. Pharmacol. 2010, 108, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Trugo, L.C.; von Baer, E.; von Baer, D. Breeding of grains: Lupin Breeding. Ref. Mod. Food Sci. 2016, 1–8. [Google Scholar] [CrossRef]
- Erdemoglu, N.; Ozkan, S.; Tosun, F. Alkaloid profile and antimicrobial activity of Lupinus angustifolius L. alkaloid extract. Phytochem. Rev. 2007, 6, 197–201. [Google Scholar] [CrossRef]
- Arnoldi, A.; Boschin, G.; Zanoni, C.; Lammi, C. The health benefits of sweet lupin seed flours and isolated proteins. J. Funct. Foods 2015, 18, 550–563. [Google Scholar] [CrossRef]
- Fontanari, G.G.; Batistuti, J.P.; da Cruz, R.J.; Saldiva, P.H.N.; Arêas, J.A.G. Cholesterol-lowering effect of whole lupin (Lupinus albus) seed and its protein isolate. Food Chem. 2012, 132, 1521–1526. [Google Scholar] [CrossRef] [PubMed]
- Sirtori, C.R.; Lovati, M.R.; Manzoni, C.M.; Castiglioni, S.; Duranti, M.; Magni, C.; Moranti, S.; D’Agostina, A.; Arnoldi, A. Proteins of white lupin seed, a naturally isoflavone-poor legume, reduce cholesterolemia in rats and increase LDL receptor activity in HepG2 cells. J. Nutr. 2004, 134, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Nasar-Abbas, S.M.; Jayasena, V. Effect of lupin flour incorporation on the physical and sensory properties of muffins. Qual. Assur. Saf. Crops Foods 2012, 4, 41–49. [Google Scholar] [CrossRef]
- Spina, A.; Cambrea, M.; Platania, A.; Roccasalva, D.; Rapisarda, P. Germoplasma di lupino (Lupinus albus L., Lupinus angustifolius L. e Lupinus luteus L.) in collezione presso il CRA-ACM di Acireale. In Conservazione, Biodiversità, Gestione Banche Dati e Miglioramento Genetico; D’Andrea, F., Ed.; Edizioni Nuova Cultura: Roma, Italy, 2013; pp. 271–279. [Google Scholar] [CrossRef]
- Campolo, O.; Romeo, F.V.; Malacrinò, A.; Laudani, F.; Carpinteri, G.; Fabroni, S.; Rapisarda, P.; Palmeri, V. Effects of inert dusts applied alone and in combination with sweet orange essential oil against Rhyzopertha dominica (Coleoptera: Bostrichidae) and wheat microbial population. Ind. Crop. Prod. 2014, 61, 361–369. [Google Scholar] [CrossRef]
- Li Destri Nicosia, M.G.; Pangallo, S.; Raphael, G.; Romeo, F.V.; Strano, M.C.; Rapisarda, P.; Droby, S.; Schena, L. Control of postharvest fungal rots on citrus fruit and sweet cherries using a pomegranate peel extract. Postharvest Biol. Technol. 2016, 114, 54–61. [Google Scholar] [CrossRef]
- Romeo, F.V.; Ballistreri, G.; Fabroni, S.; Pangallo, S.; Li Destri Nicosia, M.G.; Schena, L.; Rapisarda, P. Chemical characterization of different sumac and pomegranate extracts effective against Botrytis cinerea rots. Molecules 2015, 20, 11941–11958. [Google Scholar] [CrossRef] [PubMed]
- Calabrò, S.; Cutrignelli, M.; Lo Presti, V.; Tudisco, R.; Chiofalo, V.; Grossi, M.; Infascelli, F.; Chiofalo, B. Characterization and effect of year of harvest on the nutritional properties of three varieties of white lupine (Lupinus albus L.). J. Sci. Food Agric. 2015, 95, 3127–3136. [Google Scholar] [CrossRef] [PubMed]
- NIST Chemistry WebBook 2015. Available online: http://webbook.nist.gov/ (accessed on 05 May 2016).
- El-Shazly, A.; Ateya, A.M.M.; Wink, M. Quinolizidine Alkaloid Profiles of Lupinus varius orientalis, L. albus albus, L. hartwegii, and L. densiflorus. Z. Naturforsch. C 2001, 56, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Torres, K.B.; Quintos, N.R.; Herrera, J.M.; Tei, A.; Wink, M. Alkaloid profiles of leaves and seeds of Lupinus aschenbornii Schauer from Mexico. In Lupin, an Ancient Crop for the New Millennium, Proceedings of the 9th International Lupin Conference, Klink/Miiritz, Germany, 20–24 June 1999; van Santen, E., Wink, M., Weissmann, S., Roemer, P., Eds.; International Lupin Association Publisher: Canterbury, New Zealand, 1999; pp. 301–304. ISBN 0-86476-123-6. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard. In Clinical and Laboratory Standards Institute Document M7-A7, 7th ed.; CLSI: Wayne, PA, USA, 2006; ISBN 1-56238-587-9. [Google Scholar]
- CLSI. Reference method for broth dilution antifungal susceptibility testing of yeasts; Approved standard. In Clinical and Laboratory Standards Institute document M27-A3, 3rd ed.; CLSI: Wayne, PA, USA, 2008; ISBN 1-56238-666-2. [Google Scholar]
- Hernández, E.M.; Rangel, M.L.C.; Corona, A.E.; Cantor del Angel, A.E.; Sánchez López, J.A.; Sporer, F.; Wink, M.; Torres, K.B. Quinolizidine alkaloid composition in different organs of Lupinus aschenbornii. Rev. Bras. Farmacogn. 2011, 21, 824–828. [Google Scholar] [CrossRef]
- Law 1096 of the 25 November 1971 and Subsequent Amendments and Additions. pp. 1–28. Available online: http://scs.entecra.it/leggiEdisposizioni/NORM-NAZIONALI/Legge-1096-del-25-11-71.pdf (accessed on 18 September 2017).
- Jansen, G.; Jürgens, H.U.; Ordon, F. Effects of temperature on the alkaloid content of seeds of Lupinus angustifolius cultivars. J. Agron. Crop Sci. 2009, 195, 172–177. [Google Scholar] [CrossRef]
- Barlóg, P.K. Effect of magnesium and nitrogenous fertilisers on the growth and alkaloid content in Lupinus angustifolius L. Aust. J. Agric. Res. 2002, 53, 671–676. [Google Scholar] [CrossRef]
- Jayasena, V.; Nasar-Abbas, S.M. Development and quality evaluation of high-protein and high-dietary-fiber pasta using lupin flour. J. Texture Stud. 2012, 43, 153–163. [Google Scholar] [CrossRef]
- Erdemoglu, N.; Ozkan, S.; Duran, A.; Tosun, F. GC-MS analysis and antimicrobial activity of alkaloid extract from Genista vuralii. Pharm. Biol. 2009, 47, 81–85. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Balice, M.P.; Donelli, G.; Varaldo, P.E. Antibiotic resistance related to biofilm formation in Klebsiella pneumoniae. Pathogens 2014, 3, 743–758. [Google Scholar] [CrossRef] [PubMed]
- Lambert, P.A. Mechanisms of antibiotic resistance in Pseudomonas aeruginosa. J. R. Soc. Med. 2002, 95, 22–26. [Google Scholar] [PubMed]
- Khan, M.K.; Karnpanit, W.; Nasar-Abbas, S.M.; Huma, Z.E.; Jayasena, V. Phytochemical composition and bioactivities of lupin: A review. Int. J. Food Sci. Techol. 2015, 50, 2004–2012. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Peak n. | Alkaloid | RI | M+ | Characteristic Ions | Lupinus albus | Lupinus luteus | Lupinus angustifolius |
|---|---|---|---|---|---|---|---|
| 1 | lupinine | 1464 | 169 | 83-152-138 | X | ||
| 2 | unknown_a | 1624 | 208 | 121-175 | X | ||
| 3 | gramine | 1679 | 174 | 130-103-77 | X | ||
| 4 | genisteine (α-isosparteine) | 1759 | 234 | 98-137 | X | ||
| 5 | sparteine | 1827 | 234 | 137-98 | X | X | |
| 6 | unknown_b | 1879 | 232 | 134-232-98 | X | ||
| 7 | β-isosparteine | 1883 | 234 | 137-98 | X | X | |
| 8 | 11,12-dehydrosparteine | 1893 | 232 | 134 | X | X | X |
| 9 | ammodendrine | 1932 | 208 | 165-110 | X | X | X |
| 10 | unknown_c | 1951 | 208 | 166-136-110 | X | ||
| 11 | albine | 1984 | 232 | 191-110 | X | X | X |
| 12 | unknown_d | 2030 | 232 | 191-110 | X | ||
| 13 | isoangustifoline | 2127 | 234 | 193-112 | X | X | |
| 14 | tetrahydrhorombifoline | 2135 | 248 | 207 | X | X | |
| 15 | angustifoline | 2178 | 234 | 193-112 | X | X | |
| 16 | α-isolupanine | 2206 | 248 | 136-248 | X | X | |
| 17 | 5,6-dehydrolupanine | 2225 | 246 | 98 | X | X | |
| 18 | unknown_e | 2235 | 246 | 150-136-110 | X | ||
| 19 | lupanine | 2273 | 248 | 136-149 | X | X | X |
| 20 | 11,12-dehydrolupanine | 2296 | 246 | 134-246 | X | ||
| 21 | unknown_f | 2308 | 248 | 110-191-149 | X | ||
| 22 | 11,12-seco-12,13-didehydromultiflorine | 2330 | 246 | 58-205 | X | ||
| 23 | unknown_g | 2349 | 246 | 134 | X | ||
| 24 | 3β-hydroxylupanine | 2361 | 264 | 136-44 | X | ||
| 25 | unknown_h | 2429 | 262 | 150-164-96 | X | ||
| 26 | multiflorine | 2441 | 246 | 134-246 | X | X | X |
| 27 | unknown_i | 2460 | 264 | 134-152-246 | X | ||
| 28 | 17-oxolupanine | 2482 | 262 | 150-110-262 | X | ||
| 29 | N-formylangustifoline | 2502 | 262 | 193-112-221 | X | ||
| 30 | 13α-hydroxylupanine | 2534 | 264 | 152-246 | X | X | |
| 31 | unknown_l | 2570 | 264 | 152-246-134 | X | ||
| 32 | unknown_m | 2581 | 262 | 245-150 | X | ||
| 33 | unknown_n | 2609 | 260 | 260-148-112 | X | ||
| 34 | N-formylalbine | 2688 | 260 | 219-96 | X | ||
| 35 | unknown_o | 2712 | 262 | 149-150-148 | X | ||
| 36 | 13α-hydroxymultiflorine | 2727 | 262 | 150 | X | X | |
| 37 | 13α-isovaleroyloxylupanine | 2779 | 348 | 246-134-112 | X | ||
| 38 | 13α-angeloyloxylupanine | 2858 | 346 | 246 | X | ||
| 39 | 13α-tigloyloxylupanine | 2879 | 346 | 246 | X | X | |
| 40 | unknown_p | 2912 | 348 | 246-134-231 | X | ||
| 41 | unknown_q | 2930 | 348 | 246-134-112 | X | ||
| 42 | 3β-tigloyloxylupanine | 2944 | 346 | 134 | X | ||
| 43 | unknown_r | 3073 | 344 | 132-244-149 | X | ||
| 44 | feruloyllupinine | 3097 | 345 | 152-151 | X | ||
| 45 | 13α-tigloyloxymultiflorine | 3112 | 344 | 132 | X | ||
| 46 | unknown_s | 3262 | 394 | 246-134-112 | X |
| Genotype | Albine | Tetrahydro-Rhombifoline | Angustifoline | Lupanine | 13α-Hydroxy-Lupanine | 13α-Tigloyloxy-Lupanine | 3β-Tigloyloxy-Lupanine |
|---|---|---|---|---|---|---|---|
| Aster | 30.2 ± 1.3 n | 12.8 ± 0.1 g | 12.1 ± 0.5 h | 60.8 ± 0.3 m | 17.4 ± 1.2 g | 37.4 ± 2.9 l | 6.2 ± 0.2 c,d |
| Lublanc | 1012.4 ± 38.5 g | 24.1 ± 0.8 c,d | 268.4 ± 6.1 e | 11,218.6 ± 82.5 e,f | 491.7 ± 3.8 c,d | 102.0 ± 1.7 f,g | 16.6 ± 1.1 a |
| Luxor | 11.5 ± 0.2 n | 1.0 ± 0.4 n | 3.6 ± 0.3 h | 30.4 ± 1.2 m | 3.1 ± 0.3 g | 6.7 ± 0.5 m | 2.4 ± 0.1 e |
| Lutteur | 162.0 ± 5.7 m | 8.8 ± 0.6 i | 65.0 ± 1.2 g | 863.2 ± 6.1 i,l | 157.4 ± 1.8 f | 44.4 ± 2.2 i,l | 7.5 ± 1.4 c |
| Rosetta | 29.3 ± 0.2 n | 1.1 ± 0. 2 m,n | 6.4 ± 0.2 h | 361.8 ± 5.4 l,m | 4.7 ± 0.1 g | 6.4 ± 0.1 m | 1.5 ± 0.1 e |
| Calabria 1 | 1199.7 ± 10.5 f | 23.1 ± 1.0 d | 265.2 ± 8.4 e | 12,363.6 ± 138.0 c,d | 510.3 ± 15.5 c | 75.4 ± 5.8 h | 5.2 ± 1.4 d |
| Calabria 2 | 1436.2 ± 53.5 d | 19.4 ± 0.3 e,f | 466.8 ± 9.3 b | 14,480.3 ± 10.0 a | 687.2 ± 4.5 b | 227.8 ± 1.1 d | 12.1 ± 1.4 b |
| Calabria 3 | 1767.4 ± 17.6 b | 35.9 ± 2.5 b | 431.1 ± 19.0 b | 11,165.4 ± 157.3 e,f | 893.4 ± 66.7 a | 98.7 ± 2.3 g | 8.5 ± 1.0 c |
| Calabria 4 | 1232.3 ± 4.2 f | 44.1 ± 0.1 a | 444.2 ± 1.8 b | 12,935.3 ± 16.2 b,c | 413.9 ± 2.2 d | 87.6 ± 0.3 g,h | 11.1 ± 0.1 b |
| Modica | 2596.2 ± 2.2 a | 18.8 ± 0.8 d | 518.0 ± 15.0 a | 11,358.3 ± 140.4 e,f | 747.4 ± 25.0 b | 124.5 ± 7.6 e | 12.4 ± 0.6 b |
| Scicli | 1367.6 ± 60.6 e | 44.1 ± 1.6 a | 382.2 ± 13.6 c | 11,916.5 ± 130.1 d,e | 506.2 ± 41.9 c | 308.4 ± 1.6 c | 8.5 ± 0.1 c |
| Basilicata | 1451.2 ± 55.9 d | 23.5 ± 0.4 d | 454.0 ± 34.6 b | 11,583.0 ± 170.8 e,f | 864.6 ± 66.5 a | 419.6 ± 38.1 a | 10.7 ± 0.3 b |
| Molise | 668.2 ± 30.1 i | 10.1 ± 0.5 h,i | 315.8 ± 13.0 d | 13,243.8 ± 161.9 b | 562.3 ± 36.7 c | 315.0 ± 15.3 c | 12.0 ± 1.0 b |
| Puglia | 845.0 ± 1.2 h | 21.1 ± 1.1 e | 322.6 ± 7.7 d | 10,919.9 ± 6.6 f | 548.5 ± 70.1 c | 347.8 ± 2.2 b | 17.0 ± 2.4 a |
| Multitalia 1 | 136.8 ± 12.3 m | 2.9 ± 0.2 m | 20.7 ± 1.8 h | 1377.9 ± 7.0 i | 12.4 ± 0.3 g | 11.8 ± 1.1 m | 1.5 ± 0.3 e |
| Multitalia 2 | 838.1 ± 10.4 h | 11.1 ± 0.3 g,h | 124.1 ± 4.2 f | 4744.2 ± 88.0 g | 305.3 ± 4.1 e | 65.5 ± 0.4 h,i | 10.9 ± 0.5 b |
| Multitalia 3 | 375.1 ± 3.0 l | 6.5 ± 0.3 l | 41.9 ± 50.3g h | 2161.2 ± 11.8 h | 157.5 ± 1.2 f | 51.4 ± 0.1 i,l | 6.5 ± 1.0 c,d |
| Multitalia 4 | 1647.4 ± 12.3 c | 25.9 ± 0.4 c | 431.5 ± 15.7 b | 13,588.0 ± 188.3 b | 730.9 ± 10.7 b | 121.1 ± 4.5 e,f | 18.0 ± 0.7 a |
| Sig. | ** | ** | ** | ** | ** | ** | ** |
| Alkaloid | Dukat | Mister | Taper | Sig. |
|---|---|---|---|---|
| Lupinine | 177.1 ± 30.5 b | 281.4 ± 24.5 a | 194.6 ± 21.0 b | * |
| Sparteine | 139.0 ± 1.3 b | 233.9 ± 9.5 a | 120.0 ± 0.8 c | ** |
| β-Isosparteine | 4.5 ± 0.1 | 5.7 ± 0.9 | 4.2 ± 0.4 | n.s. |
| Ammodendrine | 26.8 ± 0.4 a,b | 31.2 ± 0.3 a | 22.7 ± 2.5 c | * |
| Unknown_c | 7.6 ± 0.2 | 8.5 ± 0.7 | 7.6 ± 0.7 | n.s. |
| Lupanine | 8.7 ± 0.5 b | 36.6 ± 4.7 a | 6.4 ± 0.6 b | ** |
| Feruloyllupinine | 12.8 ± 0.8 a | 13.1 ± 0.5 a | 9.1 ± 0.2 b | * |
| Microorganism | Multitalia 4 | Calabria 2 |
|---|---|---|
| Pseudomonas aeruginosa | 128 | 67 |
| (from skin) | ||
| Klebsiella pneumoniae | 16 | 67 |
| (from inguinal skin) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romeo, F.V.; Fabroni, S.; Ballistreri, G.; Muccilli, S.; Spina, A.; Rapisarda, P. Characterization and Antimicrobial Activity of Alkaloid Extracts from Seeds of Different Genotypes of Lupinus spp. Sustainability 2018, 10, 788. https://doi.org/10.3390/su10030788
Romeo FV, Fabroni S, Ballistreri G, Muccilli S, Spina A, Rapisarda P. Characterization and Antimicrobial Activity of Alkaloid Extracts from Seeds of Different Genotypes of Lupinus spp. Sustainability. 2018; 10(3):788. https://doi.org/10.3390/su10030788
Chicago/Turabian StyleRomeo, Flora Valeria, Simona Fabroni, Gabriele Ballistreri, Serena Muccilli, Alfio Spina, and Paolo Rapisarda. 2018. "Characterization and Antimicrobial Activity of Alkaloid Extracts from Seeds of Different Genotypes of Lupinus spp." Sustainability 10, no. 3: 788. https://doi.org/10.3390/su10030788
APA StyleRomeo, F. V., Fabroni, S., Ballistreri, G., Muccilli, S., Spina, A., & Rapisarda, P. (2018). Characterization and Antimicrobial Activity of Alkaloid Extracts from Seeds of Different Genotypes of Lupinus spp. Sustainability, 10(3), 788. https://doi.org/10.3390/su10030788