Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate


{kind=link}
{kind=link}
{kind=link}
Abstract
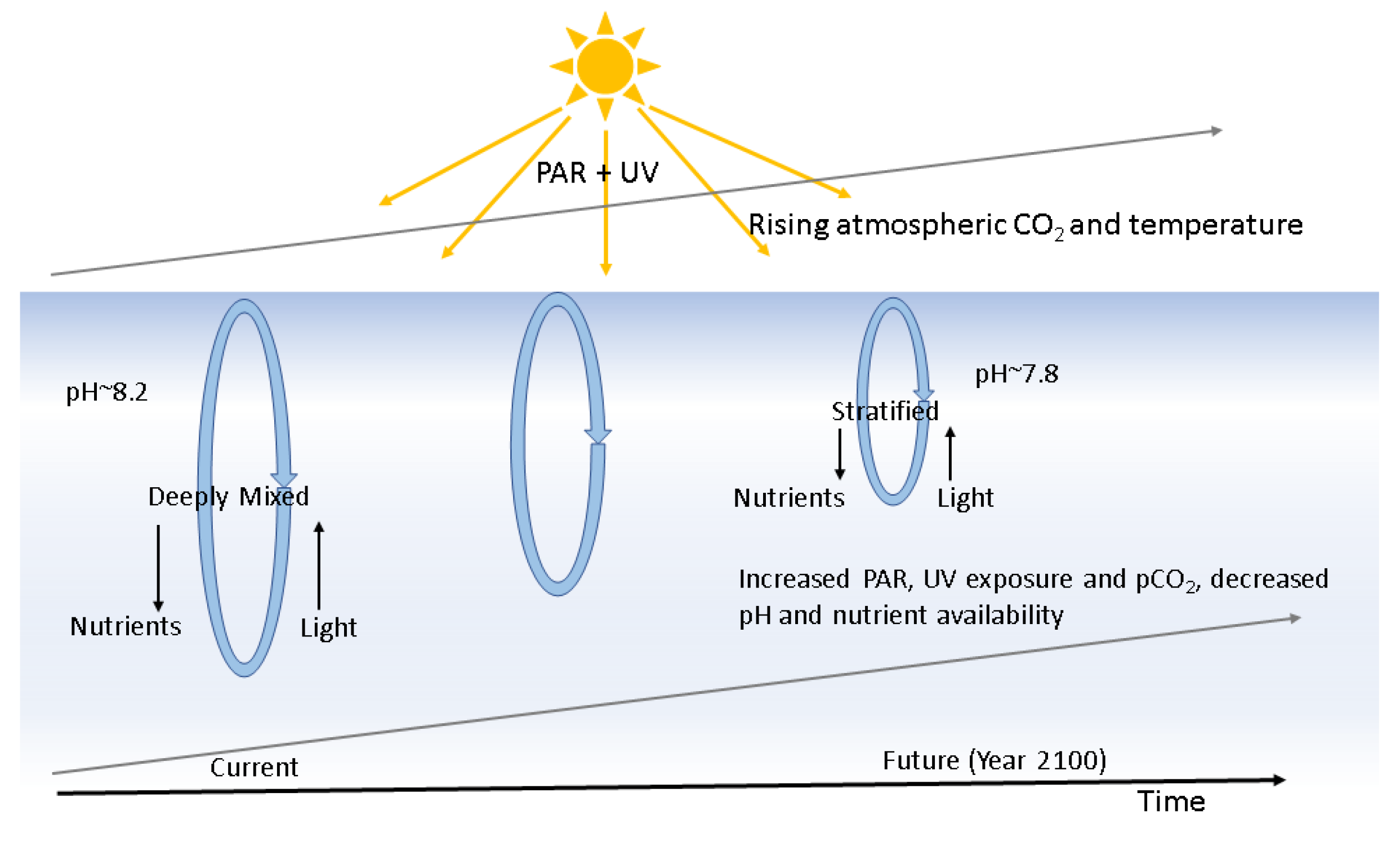
:1. Introduction
2. Components of the Biological Carbon Pump: Role of Marine Phytoplankton
3. Phytoplankton Responses to Global Change: Influence of Cell Size, Species and Elemental Composition of Phytoplankton on the Biological Carbon Pump
3.1. pCO2
3.1.1. Ocean Carbonate System
3.1.2. Role of Phytoplankton CCM
3.1.3. Influence of Phytoplankton Cell Size
3.1.4. Effect of Ocean Acidification on Coccolithophores
3.2. Temperature
3.2.1. Influence of Ocean Warming on Phytoplankton Community Composition
3.2.2. Effects of Ocean Warming on Organic Matter Partitioning
3.2.3. Effects of Ocean Warming on Phytoplankton Stoichiometry and TEP Production
3.3. Irradiance
Effects of UV-B Radiation
3.4. Nutrients
3.4.1. Influence of Phytoplankton Nutritional Quality on Higher Trophic Levels
3.4.2. Effects on Phytoplankton Community Composition
3.4.3. Effects of Stratification on Coastal Upwelling Regions
3.4.4. Trace Metal Micronutrient Availability
4. Conclusions and Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Baumert, H.Z.; Petzoldt, T. The role of temperature, cellular quota and nutrient concentrations for photosynthesis, growth and light-dark acclimation in phytoplankton. Limnologica 2008, 38, 313–326. [Google Scholar] [CrossRef]
- Simon, N.; Cras, A.-L.; Foulon, E.; Lemée, R. Diversity and evolution of marine phytoplankton. C. R. Biol. 2009, 332, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.K.; Lee, K.T.; Mohamed, A.R. Current status and challenges on microalgae-based carbon capture. Int. J. Greenh. Gas Control 2012, 10, 456–469. [Google Scholar] [CrossRef]
- Hader, D.P.; Villafane, V.E.; Helbling, E.W. Productivity of aquatic primary producers under global climate change. Photochem. Photobiol. Sci. 2014, 13, 1370–1392. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Gao, K.S.; Beardall, J. Interactive effects of ocean acidification and nitrogen-limitation on the diatom phaeodactylum tricornutum. PLoS ONE 2012, 7, e51590. [Google Scholar] [CrossRef] [PubMed]
- Hulse, D.; Arndt, S.; Wilson, J.D.; Munhoven, G.; Ridgwell, A. Understanding the causes and consequences of past marine carbon cycling variability through models. Earth-Sci. Rev. 2017, 171, 349–382. [Google Scholar] [CrossRef]
- Chisholm, S.W. The iron hypothesis: Basic research meets environmental policy. Rev. Geophys. 1995, 33, 1277–1286. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Fu, F. Microorganisms and ocean global change. Nat. Microbiol. 2017, 2, 17058. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.; Henson, S.A.; Koski, M.; De la Rocha, C.L.; Painter, S.C.; Poulton, A.J.; Riley, J.; Salihoglu, B.; Visser, A.; Yool, A.; et al. The biological carbon pump in the north Atlantic. Prog. Oceanogr. 2014, 129, 200–218. [Google Scholar] [CrossRef]
- Boyd, P.W. Toward quantifying the response of the oceans’ biological pump to climate change. Front. Mar. Sci. 2015, 2. [Google Scholar] [CrossRef]
- Passow, U.; Carlson, C.A. The biological pump in a high CO2 world. Mar. Ecol. Prog. Ser. 2012, 470, 249–271. [Google Scholar] [CrossRef]
- Bhadury, P. Effects of ocean acidification on marine invertebrates—A review. Indian J. Geo-Mar. Sci. 2015, 44, 454–464. [Google Scholar]
- Sarmiento, J.L.; Slater, R.; Barber, R.; Bopp, L.; Doney, S.C.; Hirst, A.C.; Kleypas, J.; Matear, R.; Mikolajewicz, U.; Monfray, P.; et al. Response of ocean ecosystems to climate warming. Glob. Biogeochem. Cycle 2004, 18. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Reisinger, A. Climate Change 2007 Synthesis Report Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2007. [Google Scholar]
- Riebesell, U.; Schulz, K.G.; Bellerby, R.G.; Botros, M.; Fritsche, P.; Meyerhofer, M.; Neill, C.; Nondal, G.; Oschlies, A.; Wohlers, J.; et al. Enhanced biological carbon consumption in a high CO2 ocean. Nature 2007, 450, 545–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrou, K.; Kranz, S.A.; Trimborn, S.; Hassler, C.S.; Ameijeiras, S.B.; Sackett, O.; Ralph, P.J.; Davidson, A.T. Southern ocean phytoplankton physiology in a changing climate. J. Plant Physiol. 2016, 203, 135–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.; Rost, B.; Rynearson, T.A. Evolutionary potential of marine phytoplankton under ocean acidification. Evolut. Appl. 2014, 7, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: Cell size and elemental stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef]
- Bishop, J.K.B. Autonomous observations of the ocean biological carbon pump. Oceanography 2009, 22, 182–193. [Google Scholar] [CrossRef]
- Falkowski, P.; Scholes, R.J.; Boyle, E.; Canadell, J.; Canfield, D.; Elser, J.; Gruber, N.; Hibbard, K.; Hogberg, P.; Linder, S.; et al. The global carbon cycle: A test of our knowledge of earth as a system. Science 2000, 290, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.T. Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Prog. Oceanogr. 2015, 130, 205–248. [Google Scholar] [CrossRef]
- Kim, J.M.; Lee, K.; Shin, K.; Yang, E.J.; Engel, A.; Karl, D.M.; Kim, H.C. Shifts in biogenic carbon flow from particulate to dissolved forms under high carbon dioxide and warm ocean conditions. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef] [Green Version]
- Hansell, D.A.; Carlson, C.A.; Repeta, D.J.; Schlitzer, R. Dissolved organic matter in the ocean a controversy stimulates new insights. Oceanography 2009, 22, 202–211. [Google Scholar] [CrossRef]
- Carlson, C.A.; Ducklow, H.W.; Michaels, A.F. Annual flux of dissolved organic-carbon from the euphotic zone in the northwestern Sargasso Sea. Nature 1994, 371, 405–408. [Google Scholar] [CrossRef]
- Roshan, S.; DeVries, T. Efficient dissolved organic carbon production and export in the oligotrophic ocean. Nat. Commun. 2017, 8, 2036. [Google Scholar] [CrossRef] [PubMed]
- Buesseler, K.O.; Boyd, P.W. Shedding light on processes that control particle export and flux attenuation in the twilight zone of the open ocean. Limnol. Oceanogr. 2009, 54, 1210–1232. [Google Scholar] [CrossRef]
- Herndl, G.J.; Reinthaler, T. Microbial control of the dark end of the biological pump. Nat. Geosci. 2013, 6, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, M.; Schellnhuber, H.J. Oceanic acidification affects marine carbon pump and triggers extended marine oxygen holes. Proc. Natl. Acad. Sci. USA 2009, 106, 3017–3022. [Google Scholar] [CrossRef] [PubMed]
- Ducklow, H.W.; Steinberg, D.K.; Buesseler, K.O. Upper ocean carbon export and the biological pump. Oceanography 2001, 14. [Google Scholar] [CrossRef]
- De La Rocha, C.L.; Passow, U. Factors influencing the sinking of POC and the efficiency of the biological carbon pump. Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 639–658. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, R.A.; Lee, C.; Hedges, J.I.; Honjo, S.; Wakeham, S.G. A new, mechanistic model for organic carbon fluxes in the ocean based on the quantitative association of POC with ballast minerals. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 49, 219–236. [Google Scholar] [CrossRef]
- Sato, R.; Tanaka, Y.; Ishimaru, T. Species-specific house productivity of appendicularians. Mar. Ecol. Prog. Ser. 2003, 259, 163–172. [Google Scholar] [CrossRef]
- Nishibe, Y.; Takahashi, K.; Ichikawa, T.; Hidaka, K.; Kurogi, H.; Segawa, K.; Saito, H. Degradation of discarded appendicularian houses by oncaeid copepods. Limnol. Oceanogr. 2015, 60, 967–976. [Google Scholar] [CrossRef]
- Richardson, T.L.; Jackson, G.A. Small phytoplankton and carbon export from the surface ocean. Science 2007, 315, 838–840. [Google Scholar] [CrossRef] [PubMed]
- Ragueneau, O.; Schultes, S.; Bidle, K.; Claquin, P.; Moriceau, B. Si and c interactions in the world ocean: Importance of ecological processes and implications for the role of diatoms in the biological pump. Glob. Biogeochem. Cycle 2006, 20. [Google Scholar] [CrossRef]
- Miklasz, K.A.; Denny, M.W. Diatom sinking speeds: Improved predictions and insight from a modified stokes’ law. Limnol. Oceanogr. 2010, 55, 2513–2525. [Google Scholar] [CrossRef]
- Bopp, L.; Aumont, O.; Cadule, P.; Alvain, S.; Gehlen, M. Response of diatoms distribution to global warming and potential implications: A global model study. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Iglesias-Rodriguez, M.D.; Halloran, P.R.; Rickaby, R.E.M.; Hall, I.R.; Colmenero-Hidalgo, E.; Gittins, J.R.; Green, D.R.H.; Tyrrell, T.; Gibbs, S.J.; von Dassow, P.; et al. Phytoplankton calcification in a high-CO2 world. Science 2008, 320, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.O.; Mousing, E.A.; Richardson, K. Using species distribution modelling to predict future distributions of phytoplankton: Case study using species important for the biological pump. Mar. Ecol.-Evol. Perspect. 2017, 38, e12427. [Google Scholar] [CrossRef]
- Marinov, I.; Doney, S.C.; Lima, I.D. Response of ocean phytoplankton community structure to climate change over the 21st century: Partitioning the effects of nutrients, temperature and light. Biogeosciences 2010, 7, 3941–3959. [Google Scholar] [CrossRef]
- Moreno, A.R.; Hagstrom, G.I.; Primeau, F.W.; Levin, S.A.; Martiny, A.C. Marine phytoplankton stoichiometry mediates nonlinear interactions between nutrient supply, temperature, and atmospheric CO2. Biogeosci. Discuss. 2017, 2017, 1–28. [Google Scholar] [CrossRef]
- Irwin, A.J.; Finkel, Z.V.; Muller-Karger, F.E.; Ghinaglia, L.T. Phytoplankton adapt to changing ocean environments. Proc. Natl. Acad. Sci. USA 2015, 112, 5762–5766. [Google Scholar] [CrossRef] [PubMed]
- King, A.L.; Jenkins, B.D.; Wallace, J.R.; Liu, Y.; Wikfors, G.H.; Milke, L.M.; Meseck, S.L. Effects of CO2 on growth rate, C:N:P, and fatty acid composition of seven marine phytoplankton species. Mar. Ecol. Prog. Ser. 2015, 537, 59–69. [Google Scholar] [CrossRef]
- Das, S.; Mangwani, N. Ocean acidification and marine microorganisms: Responses and consequences. Oceanologia 2015, 57, 349–361. [Google Scholar] [CrossRef]
- Mackey, K.R.M.; Morris, J.J.; Morel, F.M.M.; Kranz, S.A. Response of photosynthesis to ocean acidification. Oceanography 2015, 28, 74–91. [Google Scholar] [CrossRef]
- Häder, D.-P.; Gao, K. Interactions of anthropogenic stress factors on marine phytoplankton. Front. Environ. Sci. 2015, 3. [Google Scholar] [CrossRef]
- Beardall, J.; Stojkovic, S.; Larsen, S. Living in a high CO2 world: Impacts of global climate change on marine phytoplankton. Plant Ecol. Divers. 2009, 2, 191–205. [Google Scholar] [CrossRef]
- Mercado, J.M.; Gordillo, F.J.L. Inorganic carbon acquisition in algal communities: Are the laboratory data relevant to the natural ecosystems? Photosynth. Res. 2011, 109, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Rost, B.; Richter, K.U.; Riebesell, U.; Hansen, P.J. Inorganic carbon acquisition in red tide dinoflagellates. Plant Cell Environ. 2006, 29, 810–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Campbell, D.A.; Irwin, A.J.; Suggett, D.J.; Finkel, Z.V. Ocean acidification enhances the growth rate of larger diatoms. Limnol. Oceanogr. 2014, 59, 1027–1034. [Google Scholar] [CrossRef]
- Gazeau, F.; Salon, A.; Pitta, P.; Tsiola, A.; Maugendre, L.; Giani, M.; Celussi, M.; Pedrotti, M.L.; Marro, S.; Guieu, C. Limited impact of ocean acidification on phytoplankton community structure and carbon export in an oligotrophic environment: Results from two short-term mesocosm studies in the mediterranean sea. Estuar. Coast. Shelf Sci. 2017, 186, 72–88. [Google Scholar] [CrossRef]
- Taucher, J.; Jones, J.; James, A.; Brzezinski, M.A.; Carlson, C.A.; Riebesell, U.; Passow, U. Combined effects of CO2 and temperature on carbon uptake and partitioning by the marine diatoms thalassiosira weissflogii and dactyliosolen fragilissimus. Limnol. Oceanogr. 2015, 60, 901–919. [Google Scholar] [CrossRef]
- Tortell, P.D.; Payne, C.D.; Li, Y.; Trimborn, S.; Rost, B.; Smith, W.O.; Riesselman, C.; Dunbar, R.B.; Sedwick, P.; DiTullio, G.R. CO2 sensitivity of southern ocean phytoplankton. Geophys. Res. Lett. 2008, 35, L04605. [Google Scholar] [CrossRef]
- Fischer, G.; Karakaş, G. Sinking rates and ballast composition of particles in the Atlantic Ocean: Implications for the organic carbon fluxes to the deep ocean. Biogeosciences 2009, 6, 85–102. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Paul, C.; Moustaka-Gouni, M. Warming and ocean acidification effects on phytoplankton-from species shifts to size shifts within species in a mesocosm experiment. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rost, B.; Riebesell, U. Coccolithophores and the biological pump: Responses to environmental changes. In Coccolithophores: From Molecular Processes to Global Impact; Springer: Berlin/Heidelberg, Germany, 2004; pp. 99–125. [Google Scholar]
- Fabry, V.J. Ocean science. Marine calcifiers in a high-CO2 ocean. Science 2008, 320, 1020–1022. [Google Scholar] [CrossRef] [PubMed]
- O’dea, S.A.; Gibbs, S.J.; Bown, P.R.; Young, J.R.; Poulton, A.J.; Newsam, C.; Wilson, P.A. Coccolithophore calcification response to past ocean acidification and climate change. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Beaufort, L.; Probert, I.; de Garidel-Thoron, T.; Bendif, E.M.; Ruiz-Pino, D.; Metzl, N.; Goyet, C.; Buchet, N.; Coupel, P.; Grelaud, M.; et al. Sensitivity of coccolithophores to carbonate chemistry and ocean acidification. Nature 2011, 476, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Moran, X.A.G.; Lopez-Urrutia, A.; Calvo-Diaz, A.; Li, W.K.W. Increasing importance of small phytoplankton in a warmer ocean. Glob. Chang. Biol. 2010, 16, 1137–1144. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincon, J.; Zabala, L.L.; Jiao, N.A.Z.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine cyanobacteria prochlorococcus and synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed]
- Sommer, U.; Lengfellner, K. Climate change and the timing, magnitude, and composition of the phytoplankton spring bloom. Glob. Chang. Biol. 2008, 14, 1199–1208. [Google Scholar] [CrossRef]
- Wohlers, J.; Engel, A.; Zollner, E.; Breithaupt, P.; Jurgens, K.; Hoppe, H.G.; Sommer, U.; Riebesell, U. Changes in biogenic carbon flow in response to sea surface warming. Proc. Natl. Acad. Sci. USA 2009, 106, 7067–7072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yvon-Durocher, G.; Dossena, M.; Trimmer, M.; Woodward, G.; Allen, A.P. Temperature and the biogeography of algal stoichiometry. Glob. Ecol. Biogeogr. 2015, 24, 562–570. [Google Scholar] [CrossRef]
- Martiny, A.C.; Ma, L.Y.; Mouginot, C.; Chandler, J.W.; Zinser, E.R. Interactions between thermal acclimation, growth rate, and phylogeny influence Prochlorococcus elemental stoichiometry. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Piontek, J.; Handel, N.; Langer, G.; Wohlers, J.; Riebesell, U.; Engel, A. Effects of rising temperature on the formation and microbial degradation of marine diatom aggregates. Aquat. Microb. Ecol. 2009, 54, 305–318. [Google Scholar] [CrossRef]
- Engel, A. Direct relationship between CO2 uptake and transparent exopolymer particles production in natural phytoplankton. J. Plankton Res. 2002, 24, 49–53. [Google Scholar] [CrossRef]
- Beardall, J.; Sobrino, C.; Stojkovic, S. Interactions between the impacts of ultraviolet radiation, elevated CO2, and nutrient limitation on marine primary producers. Photochem. Photobiol. Sci. 2009, 8, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Zepp, R.G.; Erickson, D.J., III; Paul, N.D.; Sulzberger, B. Interactive effects of solar UV radiation and climate change on biogeochemical cycling. Photochem. Photobiol. Sci. 2007, 6, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Caron, D.A.; Hutchins, D.A. The effects of changing climate on microzooplankton grazing and community structure: Drivers, predictions and knowledge gaps. J. Plankton Res. 2013, 35, 235–252. [Google Scholar] [CrossRef]
- Bibby, T.S.; Mary, I.; Nield, J.; Partensky, F.; Barber, J. Low-light-adapted Prochlorococcus species possess specific antennae for each photosystem. Nature 2003, 424, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Kirilovsky, D.; Kerfeld, C.A. The orange carotenoid protein in photoprotection of photosystem II in cyanobacteria. Biochim. Biophys. Acta 2012, 1817, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Biggins, J.; Bruce, D. Regulation of excitation-energy transfer in organisms containing phycobilins. Photosynth. Res. 1989, 20, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Mackey, K.R.M.; Paytan, A.; Grossman, A.R.; Bailey, S. A photosynthetic strategy for coping in a high-light, low-nutrient environment. Limnol. Oceanogr. 2008, 53, 900–913. [Google Scholar] [CrossRef]
- Bailey, S.; Grossman, A. Photoprotection in cyanobacteria: Regulation of light harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Bibby, T.S.; Nield, J.; Barber, J. Iron deficiency induces the formation of an antenna ring around trimeric photosystem I in cyanobacteria. Nature 2001, 412, 743–745. [Google Scholar] [CrossRef] [PubMed]
- Mackey, K.R.M.; Post, A.F.; McIlvin, M.R.; Saito, M.A. Physiological and proteomic characterization of light adaptations in marine synechococcus. Environ. Microbiol. 2017, 19, 2348–2365. [Google Scholar] [CrossRef] [PubMed]
- Beardall, J.; Stojkovic, S. Microalgae under global environmental change: Implications for growth and productivity, populations and trophic flow. ScienceAsia 2006, 32, 1–10. [Google Scholar] [CrossRef]
- Villafane, V.E.; Helbling, E.W.; HolmHansen, O.; Chalker, B.E. Acclimatization of Antarctic natural phytoplankton assemblages when exposed to solar ultraviolet radiation. J. Plankton Res. 1995, 17, 2295–2306. [Google Scholar] [CrossRef]
- Overmann, J.; Pfennig, N. Bouyancy regulation and aggregate formation in Amoebobacter purpureus from Mahoney Lake. FEMS Microbiol. Lett. 1992, 101, 67–79. [Google Scholar] [CrossRef]
- Ma, Z.L.; Gao, K.S. Photosynthetically active and UV radiation act in an antagonistic way in regulating buoyancy of Arthrospira (Spirulina) platensis (cyanobacterium). Environ. Exp. Bot. 2009, 66, 265–269. [Google Scholar] [CrossRef]
- Ng, W.O.; Zentella, R.; Wang, Y.S.; Taylor, J.S.A.; Pakrasi, H.B. PhrA, the major photoreactivating factor in the cyanobacterium Synechocystis sp. strain PCC 6803 codes for a cyclobutane-pyrimidine-dimer-specific DNA photolyase. Arch. Microbiol. 2000, 173, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Wolfe-Simon, F.; Grzebyk, D.; Schofield, O.; Falkowski, P.G. The role and evolution of superoxide dismutases in algae. J. Phycol. 2005, 41, 453–465. [Google Scholar] [CrossRef]
- Bothwell, M.L.; Sherbot, D.; Roberge, A.C.; Daley, R.J. Influence of natural ultraviolet-radiation on lotic periphytic diatom community growth, biomass accrual, and species composition: Short-term versus long-term effects. J. Phycol. 1993, 29, 24–35. [Google Scholar] [CrossRef]
- Hipkin, C.R.; Thomas, R.J.; Syrett, P.J. Effects of nitrogen deficiency on nitrate reductase, nitrate assimilation and photosynthesis in unicellular marine-algae. Mar. Biol. 1983, 77, 101–105. [Google Scholar] [CrossRef]
- Cermeno, P.; Dutkiewicz, S.; Harris, R.P.; Follows, M.; Schofield, O.; Falkowski, P.G. The role of nutricline depth in regulating the ocean carbon cycle. Proc. Natl. Acad. Sci. USA 2008, 105, 20344–20349. [Google Scholar] [CrossRef] [PubMed]
- Kemp, A.E.S.; Villareal, T.A. High diatom production and export in stratified waters—A potential negative feedback to global warming. Prog. Oceanogr. 2013, 119, 4–23. [Google Scholar] [CrossRef]
- Capone, D.G.; Hutchins, D.A. Microbial biogeochemistry of coastal upwelling regimes in a changing ocean. Nat. Geosci. 2013, 6, 711–717. [Google Scholar] [CrossRef]
- Mackey, K.R.M.; Chien, C.T.; Paytan, A. Microbial and biogeochemical responses to projected future nitrate enrichment in the California upwelling system. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Rykaczewski, R.R.; Dunne, J.P. Enhanced nutrient supply to the California current ecosystem with global warming and increased stratification in an earth system model. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Breitbarth, E.; Bellerby, R.J.; Neill, C.C.; Ardelan, M.V.; Meyerhofer, M.; Zollner, E.; Croot, P.L.; Riebesell, U. Ocean acidification affects iron speciation during a coastal seawater mesocosm experiment. Biogeosciences 2010, 7, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.L.; Xu, Y.; Hopkinson, B.M.; Morel, F.M.M. Effect of ocean acidification on iron availability to marine phytoplankton. Science 2010, 327, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.J.; Breitbarth, E.; Boyd, P.W.; Hunter, K.A. Influence of ocean warming and acidification on trace metal biogeochemistry. Mar. Ecol. Prog. Ser. 2012, 470, 191–205. [Google Scholar] [CrossRef]
- Moore, J.K.; Doney, S.C.; Glover, D.M.; Fung, I.Y. Iron cycling and nutrient-limitation patterns in surface waters of the world ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 463–507. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Arrigo, K.R.; Berman-Frank, I.; Bopp, L.; Boyd, P.W.; Galbraith, E.D.; Geider, R.J.; Guieu, C.; Jaccard, S.L.; et al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Mackey, K.R.M.; Post, A.F.; McIlvin, M.R.; Cutter, G.A.; John, S.G.; Saito, M.A. Divergent responses of atlantic coastal and oceanic synechococcus to iron limitation. Proc. Natl. Acad. Sci. USA 2015, 112, 9944–9949. [Google Scholar] [CrossRef] [PubMed]
- Mackey, K.R.M.; Buck, K.N.; Casey, J.R.; Cid, A.; Lomas, M.W.; Sohrin, Y.; Paytan, A. Phytoplankton responses to atmospheric metal deposition in the coastal and open-ocean Sargasso sea. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Mahowald, N.M.; Hamilton, D.S.; Mackey, K.R.M.; Moore, J.K.; Baker, A.R.; Scanza, R.A.; Zhang, Y. Aerosol trace metal deposition dissolution and impacts on marine microorganisms and biogeochemistry. Nat. Commun. 2018, in press. [Google Scholar]
- Andersson, A.J.; Kline, D.I.; Edmunds, P.J.; Archer, S.D.; Bednaršek, N.; Carpenter, R.C.; Chadsey, M.; Goldstein, P.; Grottoli, A.G.; Hurst, T.P.; et al. Understanding ocean acidification impacts on organismal to ecological scales. Oceanography 2015, 28, 16–27. [Google Scholar] [CrossRef]
- Shi, D.; Kranz, S.A.; Kim, J.M.; Morel, F.M. Ocean acidification slows nitrogen fixation and growth in the dominant diazotroph trichodesmium under low-iron conditions. Proc. Natl. Acad. Sci. USA 2012, 109, E3094–E3100. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, S.; Morris, J.J.; Follows, M.J.; Scott, J.; Levitan, O.; Dyhrman, S.T.; Berman-Frank, I. Impact of ocean acidification on the structure of future phytoplankton communities. Nat. Clim. Chang. 2015, 5, 1002–1006. [Google Scholar] [CrossRef]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basu, S.; Mackey, K.R.M. Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate. Sustainability 2018, 10, 869. https://doi.org/10.3390/su10030869
Basu S, Mackey KRM. Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate. Sustainability. 2018; 10(3):869. https://doi.org/10.3390/su10030869
Chicago/Turabian StyleBasu, Samarpita, and Katherine R. M. Mackey. 2018. "Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate" Sustainability 10, no. 3: 869. https://doi.org/10.3390/su10030869
APA StyleBasu, S., & Mackey, K. R. M. (2018). Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate. Sustainability, 10(3), 869. https://doi.org/10.3390/su10030869