Use of Organic Wastes and Industrial By-Products to Produce Filamentous Fungi with Potential as Aqua-Feed Ingredients

 ,
,  and
and 
Abstract
:1. Introduction
2. Filamentous Fungi
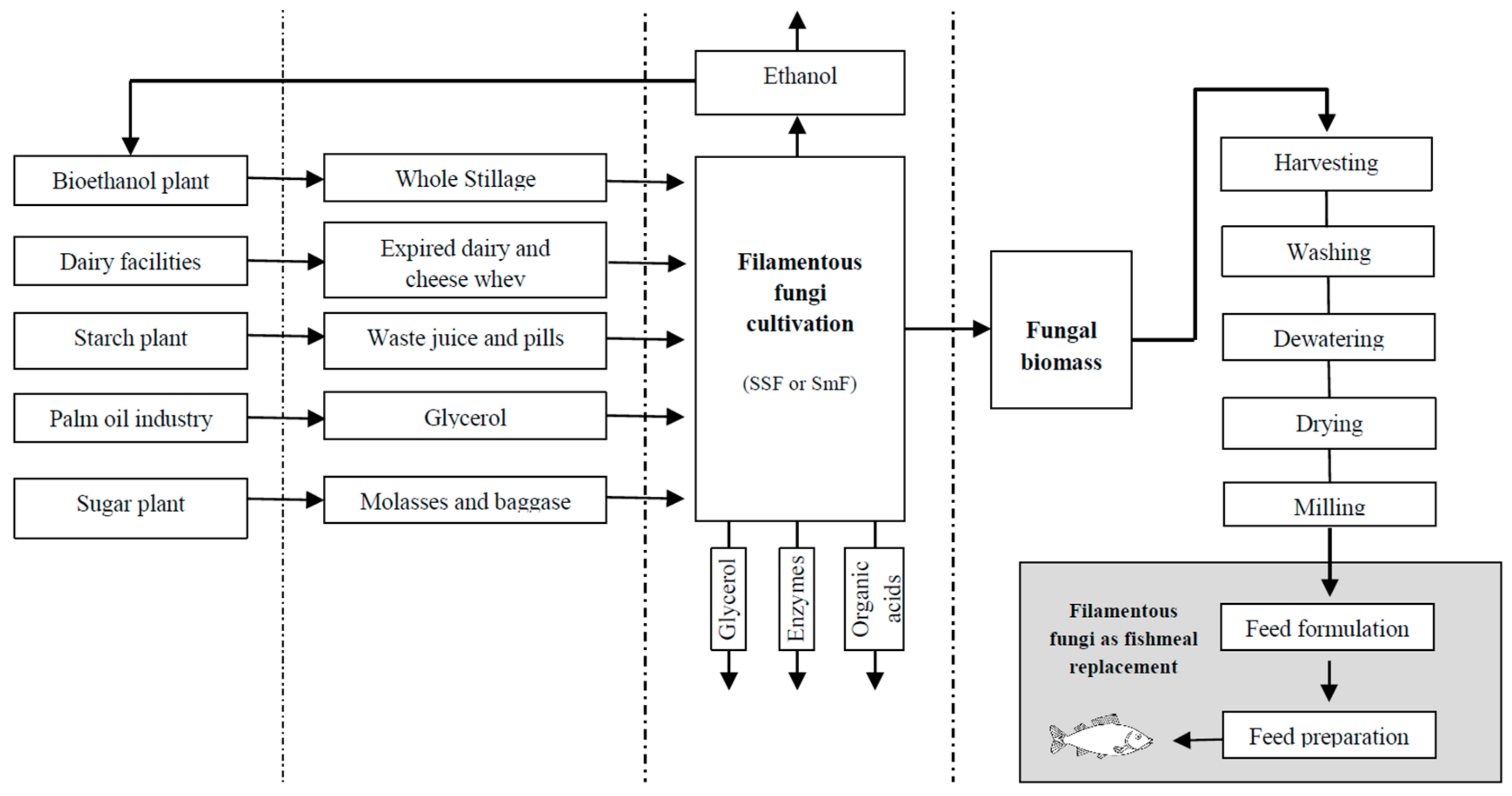
2.1. Filamentous Fungi Cultivation
2.2. Filamentous Fungi Metabolic Activity
2.3. Filamentous Fungal Biomass
3. Fish Feed
3.1. Fishmeal
3.2. Alternative Fishmeal Replacements
3.2.1. Plant-Based Sources
3.2.2. Animal-Based Sources
3.2.3. Microbial-Based Sources
Protein
Lipid
Immunostimulants
Pigments
Antioxidants
Vitamins
3.3. Challenges in the Application of Fungal Biomass as Fish Feed Supplement
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dahiya, S.; Sarkar, O.; Swamy, Y.V.; Mohan, S.V. Acidogenic fermentation of food waste for volatile fatty acid production with co-generation of biohydrogen. Bioresource Technol. 2015, 182, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Nasr, M.; Tawfik, A.; Suzuki, M.; Ookawara, S. Mathematical modeling of bio-hydrogen production from starch wastewater via up-flow anaerobic staged reactor. Desalin Water Treat. 2015, 54, 50–58. [Google Scholar] [CrossRef]
- Liang, Y.; Wen, Z. 18-bio-based nutraceuticals from biorefining. In Advances in Biorefineries; Waldron, K., Ed.; Woodhead Publishing: Cambridge, UK, 2014; pp. 596–623. [Google Scholar]
- Ferreira, J.A.; Mahboubi, A.; Lennartsson, P.R.; Taherzadeh, M.J. Waste biorefineries using filamentous ascomycetes fungi: Present status and future prospects. Bioresource Technol. 2016, 215, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Gmoser, R.; Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Filamentous ascomycetes fungi as a source of natural pigments. Fungal Biol. Biotechnol. 2017, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Lauryn, G.C.; Joshua, L.C.; Juliana Maria Leite Nobrega de Moura, B. Conversion of agricultural streams and food-processing by-products to value-added compounds using filamentous fungi. Annu. Rev. Food Sci. Technol. 2018, 9, 503–523. [Google Scholar]
- Ferreira, J.A.; Lennartsson, P.R.; Edebo, L.; Taherzadeh, M.J. Zygomycetes-based biorefinery: Present status and future prospects. Bioresource Technol. 2013, 135, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadar, Y.; Cohenarazi, E. Chemical-composition of the edible mushroom pleurotus-ostreatus produced by fermentation. Appl. Environ. Microb. 1986, 51, 1352–1354. [Google Scholar]
- Schüffler, A.; Anke, T. Secondary metabolites of basidiomycetes. In Physiology and Genetics: Selected Basic and Applied Aspects; Anke, T., Weber, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 209–231. [Google Scholar]
- Lennartsson, P.R. Zygomycetes and Cellulose Residuals: Hydrolysis, Cultivation and Applications; Chalmers Tekniska Högskola: Götenburg, Sweden, 2012. [Google Scholar]
- Tacon, A.G.J.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Azaza, M.S.; Mensi, F.; Kammoun, W.; Abdelouaheb, A.; Brini, B.; Kraiem, M. Nutritional evaluation of waste date fruit as partial substitute for soybean meal in practical diets of juvenile nile tilapia, oreochromis niloticus l. Aquacult. Nutr. 2009, 15, 262–272. [Google Scholar] [CrossRef]
- Khan, N.; Qureshi, N.A.; Nasir, M.; Vandenberg, G.W.; Mughal, M.S.; Maqbool, A.; Jabbar, M.A.; Zikria, N. Effect of artificial feed on sensory attributes of flesh of indian major carps (Labeo rohita, Catla catla and Cirrhinus mrigala) fed in monoculture and polyculture systems. Pak. Vet. J. 2012, 32, 349–353. [Google Scholar]
- Naz, S.; Javed, M. Growth responses of fish during chronic exposure of metal mixture under laboratory conditions. Pak. Vet. J. 2013, 33, 354–357. [Google Scholar]
- Yildirim, O.; Acar, U.; Turker, A.; Sunar, M.C.; Kesbic, O.S. Effects of replacing fish meal with peanut meal (Arachis hypogaea) on growth, feed utilization and body composition of mozambique tilapia fries (Oreochromis mossambicus). Pak. J. Zool. 2014, 46, 497–502. [Google Scholar]
- Kristofersson, D.; Anderson, J.L. Is there a relationship between fisheries and farming? Interdependence of fisheries, animal production and aquaculture. Mar. Policy 2006, 30, 721–725. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018; FAO: Rome, Italy, 2018; p. 227. [Google Scholar]
- Luo, Z.; Liu, C.X.; Wen, H. Effect of dietary fish meal replacement by canola meal on growth performance and hepatic intermediary metabolism of genetically improved farmed tilapia strain of nile tilapia, oreochromis niloticus, reared in fresh water. J. World Aquacult. Soc. 2012, 43, 670–678. [Google Scholar] [CrossRef]
- Overland, M.; Karlsson, A.; Mydland, L.T.; Romarheim, O.H.; Skrede, A. Evaluation of Candida utilis, Kluyveromyces marxianus and Saccharomyces cerevisiae yeasts as protein sources in diets for atlantic salmon (salmo salar). Aquaculture 2013, 402, 1–7. [Google Scholar] [CrossRef]
- Nair, R.B. Integration of First and Second Generation Bioethanol Processes: Using Edible Filamentous Fungus Neurospora Intermedia; Högskolan i Borås: Borås, Sweden, 2017. [Google Scholar]
- Nitayavardhana, S.; Issarapayup, K.; Pavasant, P.; Khanal, S.K. Production of protein-rich fungal biomass in an airlift bioreactor using vinasse as substrate. Bioresource Technol. 2013, 133, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Lucking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Nalage, D.; Khedkar, G.; Kalyankar, A.; Sarkate, A.P.; Sr, G.; Vb, B.; Khedkar, C. Single cell proteins. In The Encyclopedia of Food and Health, 1st ed.; Academic Press: Oxford/London, UK, 2016; pp. 790–794. [Google Scholar]
- Shuvaeva, G.P.; Sysoeva, M.G. Xylanase of the micromycete rhizopus var. Microsporus 595: Preparation, structural and functional characteristics, and application. Appl. Biochem. Microbiol. 2010, 46, 641–647. [Google Scholar] [CrossRef]
- Adejoye, O.D.; Adebayo-Tayo, B.C.; Ogunjobi, A.A.; Olaoye, O.A.; Fadahunsi, F.I. Effect of carbon, nitrogen and mineral sources on growth of pleurotus Florida, a Nigeria edible mushroom. Afr. J. Biotechnol. 2006, 5, 1355–1359. [Google Scholar]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli physiology in luria-bertani broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [PubMed]
- Andualem, B.; Gessesse, A. Production of microbial medium from defatted brebra (Milletia ferruginea) seed flour to substitute commercial peptone agar. Asian Pac. J. Trop. Biomed. 2013, 3, 790–797. [Google Scholar] [CrossRef]
- Demirbas, A. Political, economic and environmental impacts of biofuels: A review. Appl. Energy 2009, 86, S108–S117. [Google Scholar] [CrossRef]
- Mahboubi, A.; Ferreira, J.A.; Taherzadeh, M.J.; Lennartsson, P.R. Value-added products from dairy waste using edible fungi. Waste Manag. 2017, 59, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhao, X.F.; Strait, M.; Wen, Z.Y. Use of dry-milling derived thin stillage for producing eicosapentaenoic acid (epa) by the fungus pythium irregulare. Bioresource Technol. 2012, 111, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Jin, B.; Kelly, J.M. Production of l(+)-lactic acid using acid-adapted precultures of rhizopus arrhizus in a stirred tank reactor. Appl. Biochem. Biotechnol. 2008, 149, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Nout, M.J.R.; Aidoo, K.E. Asian fungal fermented food. In Industrial Applications; Hofrichter, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 29–58. [Google Scholar]
- Singh, O.V.; Kapur, N.; Singh, R.P. Evaluation of agro-food byproducts for gluconic acid production by Aspergillus niger ors-4.410. World J. Microbiol. Biotechnol. 2005, 21, 519–524. [Google Scholar] [CrossRef]
- Soccol, C.R.; Vandenberghe, L.P.S.; Rodrigues, C.; Pandey, A. New perspectives for citric acid production and application. Food Technol. Biotechnol. 2006, 44, 141–149. [Google Scholar]
- Taherzadeh, M.J.; Fox, M.; Hjorth, H.; Edebo, L. Production of mycelium biomass and ethanol from paper pulp sulfite liquor by rhizopus oryzae. Bioresource Technol. 2003, 88, 167–177. [Google Scholar] [CrossRef]
- Sues, A.; Millati, R.; Edebo, L.; Taherzadeh, M.J. Ethanol production from hexoses, pentoses, and dilute-acid hydrolyzate by mucor indicus. Fems Yeast Res. 2005, 5, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Asadollahzadeh, M.; Ghasemian, A.; Saraeian, A.; Resalati, H.; Taherzadeh, M.J. Production of fungal biomass protein by filamentous fungi cultivation on liquid waste streams from pulping process. BioResources 2018, 13, 5013–5031. [Google Scholar]
- Sharifia, M.; Karimi, K.; Taherzadeh, M.J. Production of ethanol by filamentous and yeast-like forms of mucor indicus from fructose, glucose, sucrose, and molasses. J. Ind. Microbiol. Biotechnol. 2008, 35, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Jasti, N.; Khanal Samir, K.; Pometto Anthony, L.; van Leeuwen, J. Converting corn we-tmilling effluent into high-value fungal biomass in a biofilm reactor. Biotechnol. Bioeng. 2008, 101, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, K.; Rajoli, S.; Taherzadeh, M.J. Techno-economic analysis of integrating first and second-generation ethanol production using filamentous fungi: An industrial case study. Energies 2016, 9, 359. [Google Scholar] [CrossRef]
- Souza Filho, P.; Brancoli, P.; Bolton, K.; Zamani, A.; Taherzadeh, M. Techno-economic and life cycle assessment of wastewater management from potato starch production: Present status and alternative biotreatments. Fermentation 2017, 3, 56. [Google Scholar] [CrossRef]
- Ferreira, J. Integration of Filamentous Fungi in Ethanol Dry-Mill Biorefinery. Ph.D. Thesis, Högskolan i Borås, Borås, Sweden, 2015. [Google Scholar]
- Souza Filho, P.F.; Zamani, A.; Taherzadeh, M.J. Edible protein production by filamentous fungi using starch plant wastewater. Waste Biomass Valoriz. 2018, 1–10. [Google Scholar] [CrossRef]
- Xu, Q.; Li, S.; Fu, Y.Q.; Tai, C.; Huang, H. Two-stage utilization of corn straw by rhizopus oryzae for fumaric acid production. Bioresource Technol. 2010, 101, 6262–6264. [Google Scholar] [CrossRef] [PubMed]
- Archer, D.B.; Connerton, I.F.; MacKenzie, D.A. Filamentous fungi for production of food additives and processing aids. In Food Biotechnology; Stahl, U., Donalies, U.E.B., Nevoigt, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 99–147. [Google Scholar]
- Yegin, S.; Fernandez-Lahore, M.; Salgado, A.J.G.; Guvenc, U.; Goksungur, Y.; Tari, C. Aspartic proteinases from Mucor spp. in cheese manufacturing. Appl. Microbiol. Biotechnol. 2011, 89, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Azeke, M.A.; Greiner, R.; Jany, K.D. Purification and characterization of two intracellular phytases from the tempeh fungus rhizopus oligosporus. J. Food Biochem. 2011, 35, 213–227. [Google Scholar] [CrossRef]
- Vattem, D.A.; Shetty, K. Solid-state production of phenolic antioxidants from cranberry pomace by rhizopus oligosporus. Food Biotechnol. 2002, 16, 189–210. [Google Scholar] [CrossRef]
- Liao, W.; Liu, Y.; Chen, S.L. Studying pellet formation of a filamentous fungus rhizopus oryzae to enhance organic acid production. Appl. Biochem. Biotechnol. 2007, 137, 689–701. [Google Scholar] [PubMed]
- Pandey, A.; Höfer, R.; Taherzadeh, M.; Nampoothiri, M.; Larroche, C. Industrial Biorefineries and White Biotechnology; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Dhillon, G.S.; Brar, S.K.; Kaur, S.; Verma, M. Bioproduction and extraction optimization of citric acid from Aspergillus niger by rotating drum type solid-state bioreactor. Ind. Crops Prod. 2013, 41, 78–84. [Google Scholar] [CrossRef]
- Okabe, M.; Lies, D.; Kanamasa, S.; Park, E.Y. Biotechnological production of itaconic acid and its biosynthesis in aspergillus terreus. Appl. Microbiol. Biotechnol. 2009, 84, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Terabayashi, Y.; Sano, M.; Yamane, N.; Marui, J.; Tamano, K.; Sagara, J.; Dohmoto, M.; Oda, K.; Ohshima, E.; Tachibana, K.; et al. Identification and characterization of genes responsible for biosynthesis of kojic acid, an industrially important compound from aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Stahl, P.D.; Klug, M.J. Characterization and differentiation of filamentous fungi based on fatty acid composition. Appl. Environ. Microb. 1996, 62, 4136–4146. [Google Scholar]
- Rumsey, G.L.; Winfree, R.A.; Hughes, S.G. Nutritional-value of dietary nucleic-acids and purine-bases to rainbow-trout (oncorhynchus-mykiss). Aquaculture 1992, 108, 97–110. [Google Scholar] [CrossRef]
- Kinsella, J.E.; German, B.; Shetty, J. Uricase from fish liver—Isolation and some properties. Comp. Biochem. Phys. B 1985, 82, 621–624. [Google Scholar] [CrossRef]
- Jobling, M. Feeding and digestive functions in fishes. J. Fish Biol. 2009, 75, 756–757. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011; p. 392. [Google Scholar]
- Alvim, M.C.; Peret, A.C. Food resources sustaining the fish fauna in a section of the upper sao francisco river in tres marias, mg, Brazil. Braz. J. Biol. 2004, 64, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Peretti, D.; Andrian, I.D.F. Trophic structure of fish assemblages in five permanent lagoons of the high parana river floodplain, Brazil. Environ. Biol. Fish 2004, 71, 95–103. [Google Scholar] [CrossRef]
- Cassemiro, F.A.D.S.; Hahn, N.S.; Rangel, T.F.L.V.D. Diet and trophic ecomorphology of the silverside, odontesthes bonariensis, of the salto caxias reservoir, Rio Iguaçu, Paraná, Brazil. Neotrop. Ichthyol. 2018, 1, 127–131. [Google Scholar] [CrossRef]
- Parzanini, C.; Parrish, C.C.; Hamel, J.-F.; Mercier, A. Trophic ecology of a deep-sea fish assemblage in the northwest Atlantic. Mar. Biol. 2017, 164, 206. [Google Scholar] [CrossRef]
- Sanchez-Muros, M.J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Moffitt, C.M.; Cajas-Cano, L. Blue growth: The 2014 fao state of world fisheries and aquaculture. Fisheries 2014, 39, 552–553. [Google Scholar] [CrossRef]
- Kobayashi, M.; Msangi, S.; Batka, M.; Vannuccini, S.; Dey, M.M.; Anderson, J.L. Fish to 2030: The role and opportunity for aquaculture. Aquacult. Econ. Manag. 2015, 19, 282–300. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Enes, P.; Peres, H. 8-replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish a2—Davis, d. Allen. In Feed and Feeding Practices in Aquaculture; Woodhead Publishing: Oxford, UK, 2015; pp. 203–233. [Google Scholar]
- Miles, R.D.; Chapman, F.A. The Benefits of Fish Meal in Aquaculture Diets; IFAS Extension; University of Florida: Gainesville, FL, USA, 2006. [Google Scholar]
- Lee, C.-S. Dietary Nutrients, Additives and Fish Health; John Wiley & Sons: New York, NY, USA, 2015. [Google Scholar]
- Glasser, F.; Ferlay, A.; Chilliard, Y. Oilseed lipid supplements and fatty acid composition of cow milk: A meta-analysis. J. Dairy Sci. 2008, 91, 4687–4703. [Google Scholar] [CrossRef] [PubMed]
- Čertík, M.; Baltészov, L.; Šajbidor, J. Lipid formation and γ-linolenic acid production by mucorales fungi grown on sunflower oil. Lett. Appl. Microbiol. 1997, 25, 101–105. [Google Scholar] [CrossRef]
- E Fraga, M.; Maria N Santana, D.; Gatti, M.J.A.; Direito, G.; Cavaglieri, L.; Rocha Rosa, C. Characterization of Aspergillus Species Based on Fatty acid Profiles; Fundação Oswaldo Cruz. Instituto Oswaldo Cruz: Rio, Brazil, 2008; Volume 103, pp. 540–544. [Google Scholar]
- Moutinho, S.; Martinez-Llorens, S.; Tomas-Vidal, A.; Jover-Cerda, M.; Oliva-Teles, A.; Peres, H. Meat and bone meal as partial replacement for fish meal in diets for gilthead seabream (Sparus aurata) juveniles: Growth, feed efficiency, amino acid utilization, and economic efficiency. Aquaculture 2017, 468, 271–277. [Google Scholar] [CrossRef]
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.S.; Krogdahl, A.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.; Makkar, H.P.S.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Berot, S.; Compoint, J.P.; Larre, C.; Malabat, C.; Gueguen, J. Large scale purification of rapeseed proteins (Brassica napus L.). J. Chromatogr. B 2005, 818, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Krogdahl, A.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Sawanboonchun, J.; Roy, W.J.; Robertson, D.A.; Bell, J.G. The impact of dietary supplementation with astaxanthin on egg quality in atlantic cod broodstock (Gadus morhua L.). Aquaculture 2008, 283, 97–101. [Google Scholar] [CrossRef]
- Estruch, G.; Collado, M.C.; Penaranda, D.S.; Vidal, A.T.; Cerda, M.J.; Martinez, G.P.; Martinez-Llorens, S. Impact of fishmeal replacement in diets for gilthead sea bream (Sparus aurata) on the gastrointestinal microbiota determined by pyrosequencing the 16s rrna gene. PLoS ONE 2015, 10, e0136389. [Google Scholar] [CrossRef] [PubMed]
- Bakke-McKellep, A.M.; Penn, M.H.; Salas, P.M.; Refstie, S.; Sperstad, S.; Landsverk, T.; Ringo, E.; Krogdahl, A. Effects of dietary soyabean meal, inulin and oxytetracycline on intestinal microbiota and epithelial cell stress, apoptosis and proliferation in the teleost atlantic salmon (Salmo salar L.). Br. J. Nutr. 2007, 97, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Ringo, E.; Sperstad, S.; Myklebust, R.; Refstie, S.; Krogdahl, A. Characterisation of the microbiota associated with intestine of atlantic cod (Gadus morhua L.)—The effect of fish meal, standard soybean meal and a bioprocessed soybean meal. Aquaculture 2006, 261, 829–841. [Google Scholar] [CrossRef]
- Torrecillas, S.; Montero, D.; Caballero, M.J.; Pittman, K.A.; Custodio, M.; Campo, A.; Sweetman, J.; Izquierdo, M. Dietary mannan oligosaccharides: Counteracting the side effects of soybean meal oil inclusion on european sea bass (Dicentrarchus labrax) gut health and skin mucosa mucus production? Front. Immunol. 2015, 6, 397. [Google Scholar] [CrossRef] [PubMed]
- Hardy, R.W.; Shearer, K.D. Effect of dietary calcium-phosphate and zinc supplementation on whole-body zinc concentration of rainbow-trout (salmo-gairdneri). Can. J. Fish Aquat. Sci. 1985, 42, 181–184. [Google Scholar] [CrossRef]
- Ayoola, A. Replacement of Fishmeal with Alternative Protein Source in Aquaculture Diets; North Carolina State University: Raleigh, NC, USA, 2010. [Google Scholar]
- Dufosse, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.S.; Sutthiwong, N. Filamentous fungi are large-scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Elorduy, J.; Pino Moreno, J.M.; Escamilla Prado, E.; Alvarado Perez, M.; Lagunez Otero, J.; Ladron de Guevara, O. Nutritional value of edible insects from the state of Oaxaca, Mexico. J. Food Compost. Anal. 1997, 10, 142–157. [Google Scholar] [CrossRef]
- Van Leeuwen, H.J. Fungi Cultivation on Alcohol Fermentation Stillage for Useful Products and Energy Savings. US20100196994A1. 9 July 2013. [Google Scholar]
- Barker, T.W.; Drouliscos, N.J.; Worgan, J.T. Composition and nutritional-evaluation of aspergillus-oryzae biomass grown on palm oil processing effluents. J. Sci. Food Agric. 1981, 32, 1014–1020. [Google Scholar] [CrossRef]
- Jin, B.; van Leeuwen, H.J.; Patel, B.; Doelle, H.W.; Yu, Q. Production of fungal protein and glucoamylase by rhizopus oligosporus from starch processing wastewater. Process Biochem. 1999, 34, 59–65. [Google Scholar] [CrossRef]
- Ferreira, J.A.; Lennartsson, P.R.; Niklasson, C.; Lundin, M.; Edebo, L.; Taherzadeh, M.J. Spent sulphite liquor for cultivation of an edible Rhizopus sp. Bioresources 2012, 7, 173–188. [Google Scholar]
- Mitra, D.; Rasmussen, M.L.; Chand, P.; Chintareddy, V.R.; Yao, L.X.; Grewell, D.; Verkade, J.G.; Wang, T.; van Leeuwen, J. Value-added oil and animal feed production from corn-ethanol stillage using the oleaginous fungus mucor circinelloides. Bioresource Technol. 2012, 107, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Wikandari, R.; Millati, R.; Lennartsson, P.R.; Harmayani, E.; Taherzadeh, M.J. Isolation and characterization of zygomycetes fungi from tempe for ethanol production and biomass applications. Appl. Biochem. Biotechnol. 2012, 167, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Production of ethanol and biomass from thin stillage using food-grade zygomycetes and ascomycetes filamentous fungi. Energies 2014, 7, 4199–4201. [Google Scholar] [CrossRef]
- Rasmussen, M.L.; Khanal, S.K.; Pometto, A.L.; van Leeuwen, J. Water reclamation and value-added animal feed from corn-ethanol stillage by fungal processing. Bioresource Technol. 2014, 151, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Batori, V.; Ferreira, J.A.; Taherzadeh, M.J.; Lennartsson, P.R. Ethanol and protein from ethanol plant by-products using edible fungi neurospora intermedia and aspergillus oryzae. BioMed Res. Int. 2015, 2015, 176371. [Google Scholar] [CrossRef] [PubMed]
- Yunus, F.U.N.; Nadeem, M.; Rashid, F. Single-cell protein production through microbial conversion of lignocellulosic residue (wheat bran) for animal feed. J. Inst. Brewing 2015, 121, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Mahboubi, A.; Ferreira, J.; Taherzadeh, M.; Lennartsson, P. Production of fungal biomass for feed, fatty acids, and glycerol by aspergillus oryzae from fat-rich dairy substrates. Fermentation 2017, 3, 48. [Google Scholar] [CrossRef]
- Pedneault, K.; Angers, P.; Gosselin, A.; Tweddell, R.J. Fatty acid profiles of polar and neutral lipids of ten species of higher basidiomycetes indigenous to eastern Canada. Mycol. Res. 2008, 112, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.L.; Molina-Jouve, C.; Nicaud, J.M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Amar, E.C.; Kiron, V.; Akutsu, T.; Satoh, S.; Watanabe, T. Resistance of rainbow trout oncorhynchus mykiss to infectious hematopoietic necrosis virus (ihnv) experimental infection following ingestion of natural and synthetic carotenoids. Aquaculture 2012, 330, 148–155. [Google Scholar] [CrossRef]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes. Aquaculture 2008, 274, 1–14. [Google Scholar] [CrossRef]
- Ringø, E.; Olsen, R.; González Vecino, J.; Wadsworth, S. Use of immunostimulants and nucleotides in aquaculture: A review. J. Mar. Sci. Res. Dev. 2011, 2, 104. [Google Scholar] [CrossRef]
- Staykov, Y.; Spring, P.; Denev, S.; Sweetman, J. Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss). Aquacult. Int. 2007, 15, 153–161. [Google Scholar] [CrossRef]
- Gopalakannan, A.; Arul, V. Immunomodulatory effects of dietary intake of chitin, chitosan and levamisole on the immune system of cyprinus carpio and control of aeromonas hydrophila infection in ponds. Aquaculture 2006, 255, 179–187. [Google Scholar] [CrossRef]
- Ai, Q.H.; Xu, H.G.; Mai, K.S.; Xu, W.; Wang, J.; Zhang, W.B. Effects of dietary supplementation of bacillus subtilis and fructooligosaccharide on growth performance, survival, non-specific immune response and disease resistance of juvenile large yellow croaker, larimichthys crocea. Aquaculture 2011, 317, 155–161. [Google Scholar] [CrossRef]
- Awad, E.; Austin, B. Use of lupin, lupinus perennis, mango, mangifera indica, and stinging nettle, urtica dioica, as feed additives to prevent aeromonas hydrophila infection in rainbow trout, oncorhynchus mykiss (walbaum). J. Fish Dis. 2010, 33, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Dalmo, R.A.; Bogwald, J. Beta-glucans as conductors of immune symphonies. Fish Shellfish Immunol. 2008, 25, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Jung-Schroers, V.; Adamek, M.; Jung, A.; Harris, S.; Doza, O.S.; Baumer, A.; Steinhagen, D. Feeding of beta-1,3/1,6-glucan increases the diversity of the intestinal microflora of carp (Cyprinus carpio). Aquacult. Nutr. 2016, 22, 1026–1039. [Google Scholar] [CrossRef]
- Peng, M.; Xu, W.; Ai, Q.H.; Mai, K.S.; Liufu, Z.G.; Zhang, K.K. Effects of nucleotide supplementation on growth, immune responses and intestinal morphology in juvenile turbot fed diets with graded levels of soybean meal (Scophthalmus maximus L.). Aquaculture 2013, 392, 51–58. [Google Scholar] [CrossRef]
- Nimrichter, L.; Rodrigues, M.L.; Rodrigues, E.G.; Travassos, L.R. The multitude of targets for the immune system and drug therapy in the fungal cell wall. Microbes Infect. 2005, 7, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Fleet, G.H.; Rogers, P.L. Composition of the cell walls of several yeast species. Appl. Microbiol. Biotechnol. 1998, 50, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Free, S.J. Fungal cell wall organization and biosynthesis. Adv. Genet. 2013, 81, 33–82. [Google Scholar] [PubMed]
- Peter Martin, G.; Vandamme Erick, J.; De Baets, S.; Steinbüchel, A. Chitin and Chitosan in Fungi. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/3527600035.bpol6005 (accessed on 14 September 2018).
- Whyte, S.K. The innate immune response of finfish—A review of current knowledge. Fish Shellfish Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Ringo, E.; Olsen, R.E.; Gifstad, T.O.; Dalmo, R.A.; Amlund, H.; Hemre, G.I.; Bakke, A.M. Prebiotics in aquaculture: A review. Aquacult. Nutr. 2010, 16, 117–136. [Google Scholar] [CrossRef]
- Bagni, M.; Romano, N.; Finoia, M.G.; Abelli, L.; Scapigliati, G.; Tiscar, P.G.; Sarti, M.; Marino, G. Short- and long-term effects of a dietary yeast beta-glucan (macrogard) and alginic acid (ergosan) preparation on immune response in sea bass (Dicentrarchus labrax). Fish Shellfish Immunol. 2005, 18, 311–325. [Google Scholar] [CrossRef] [PubMed]
- El-Boshy, M.E.; El-Ashram, A.M.; Abdelhamid, F.M.; Gadalla, H.A. Immunomodulatory effect of dietary saccharomyces cerevisiae, beta-glucan and laminaran in mercuric chloride treated nile tilapia (Oreochromis niloticus) and experimentally infected with aeromonas hydrophila. Fish Shellfish Immunol. 2010, 28, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Kim, J.S.; Balasundaram, C.; Heo, M.S. Dietary supplementation with chitin and chitosan on haematology and innate immune response in epinephelus bruneus against philasterides dicentrarchi. Exp. Parasitol. 2012, 131, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Yoo, G.; Lee, S.; Kim, Y.C.; Okorie, O.E.; Park, G.J.; Han, Y.O.; Choi, S.M.; Kang, J.C.; Sun, M.H.; Bai, S.C. Effects of dietary beta-1,3 glucan and feed stimulants in juvenile olive flounder, paralichthys olivaceus. J. World Aquacult. Soc. 2007, 38, 138–145. [Google Scholar] [CrossRef]
- Mari, L.S.S.; Jagruthi, C.; Anbazahan, S.M.; Yogeshwari, G.; Thirumurugan, R.; Arockiaraj, J.; Mariappan, P.; Balasundaram, C.; Harikrishnan, R. Protective effect of chitin and chitosan enriched diets on immunity and disease resistance in cirrhina mrigala against aphanomyces invadans. Fish Shellfish Immunol. 2014, 39, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Steglich, W. Biologically-active compounds from higher fungi. Pure Appl. Chem. 1981, 53, 1233–1240. [Google Scholar] [CrossRef]
- Velisek, J.; Cejpek, K. Pigments of higher fungi: A review. Czech. J. Food Sci. 2011, 29, 87–102. [Google Scholar] [CrossRef]
- Velmurugan, P.; Kamala-Kannan, S.; Balachandar, V.; Lakshmanaperumalsamy, P.; Chae, J.C.; Oh, B.T. Natural pigment extraction from five filamentous fungi for industrial applications and dyeing of leather. Carbohyd. Polym. 2010, 79, 262–268. [Google Scholar] [CrossRef]
- Nigam, P.S.; Luke, J.S. Food additives: Production of microbial pigments and their antioxidant properties. Curr. Opin. Food Sci. 2016, 7, 93–100. [Google Scholar] [CrossRef]
- Torrissen, O.J.; Hardy, R.W.; Shearer, K.D.; Scott, T.M.; Stone, F.E. Effects of dietary canthaxanthin level and lipid-level on apparent digestibility coefficients for canthaxanthin in rainbow-trout (oncorhynchus-mykiss). Aquaculture 1990, 88, 351–362. [Google Scholar] [CrossRef]
- Storebakken, T.; Goswami, U.C. Plasma carotenoid concentration indicates the availability of dietary astaxanthin for atlantic salmon, salmo salar. Aquaculture 1996, 146, 147–153. [Google Scholar] [CrossRef]
- Wang, Y.J.; Chien, Y.H.; Pan, C.H. Effects of dietary supplementation of carotenoids on survival, growth, pigmentation, and antioxidant capacity of characins, hyphessobrycon callistus. Aquaculture 2006, 261, 641–648. [Google Scholar] [CrossRef]
- Amar, E.C.; Kiron, V.; Satoh, S.; Watanabe, T. Enhancement of innate immunity in rainbow trout (oncorhynchus mykiss walbaum) associated with dietary intake of carotenoids from natural products. Fish Shellfish Immunol. 2004, 16, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, K.; Yagi, M.; Hara, K.; Mishima, T.; Tsuchimoto, M. Effects of feeding of β-carotene-supplemented rotifers on survival and lymphocyte proliferation reaction of fish larvae (Japanese parrotfish (Oplegnathus fasciatus) and spotted parrotfish (Oplegnathus punctatus)): Preliminary trials. Hydrobiologia 1997, 358, 313–316. [Google Scholar] [CrossRef]
- Bendich, A.; Olson, J.A. Biological actions of carotenoids. FASEB J. 1989, 3, 1927–1932. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.E.; Johnson, J.D. The vitamin a-redox hypothesis: A biochemical basis for honest signaling via carotenoid pigmentation. Am. Nat. 2012, 180, E127–E150. [Google Scholar] [CrossRef] [PubMed]
- McGraw, K.J.; Ardia, D.R. Carotenoids, immunocompetence, and the information content of sexual colors: An experimental test. Am. Nat. 2003, 162, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Von Schantz, T.; Bensch, S.; Grahn, M.; Hasselquist, D.; Wittzell, H. Good genes, oxidative stress and condition-dependent sexual signals. Proc. R. Soc. B Biol. Sci. 1999, 266, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Maan, M.E.; Sefc, K.M. Colour variation in cichlid fish: Developmental mechanisms, selective pressures and evolutionary consequences. Semin. Cell Dev. Biol. 2013, 24, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Baerends, G.P.; Wanders, J.B.W.; Vodegel, R. The relationship between marking patterns and motivational state in the prespawning behavior of the cichlid fish chromidotilapia guentheri (sauvage). Neth. J. Zool. 1986, 36, 88–116. [Google Scholar] [CrossRef]
- Kalinowski, C.T.; Robaina, L.E.; Fernandez-Palacios, H.; Schuchardt, D.; Izquierdo, M.S. Effect of different carotenoid sources and their dietary levels on red porgy (Pagrus pagrus) growth and skin colour. Aquaculture 2005, 244, 223–231. [Google Scholar] [CrossRef]
- Teles Aires, O.; Lupatsch, I.; Nengas, I. Nutrition and Feeding of Sparidae. Available online: https://cronfa.swan.ac.uk/Record/cronfa8250 (accessed on 14 September 2018).
- Barja, G. Free radicals and aging. Trends Neurosci. 2004, 27, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.M.; Buettner, G.R.; Aust, S.D. Transition-metals as catalysts of autoxidation reactions. Free Radic. Biol. Med. 1990, 8, 95–108. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Schuemann, J.; Hertweck, C. Biosynthesis of fungal polyketides. Mycota 2009, 15, 331–351. [Google Scholar]
- Tian, C.C.; Zha, X.Q.; Pan, L.H.; Luo, J.P. Structural characterization and antioxidant activity of a low-molecular polysaccharide from dendrobium huoshanense. Fitoterapia 2013, 91, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Dhale, M.A.; Divakar, S.; Kumar, S.U.; Vijayalakshmi, G. Isolation and characterization of dihydromonacolin-mv from monascus purpureus for antioxidant properties. Appl. Microbiol. Biotechnol. 2007, 73, 1197–1202. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.A. Production of Antimicrobials and Antioxidants from Filamentous Fungi; National University of Ireland Maynooth: Kildare, Ireland, 2014. [Google Scholar]
- Gopalakrishnan, K.; Van Leeuwen, H.; Brown, R. Sustainable Bioenergy and Bioproducts: Value Added Engineering Applications 001; Springer: Berlin, Germany, 2012; Volume 62. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaiah, N. A review on fungal diseases of algae, marine fishes, shrimps and corals. Indian J. Mar. Sci. 2006, 35, 380–387. [Google Scholar]
- Tisma, M.; Zelic, B.; Vasic-Racki, D. White-rot fungi in phenols, dyes and other xenobiotics treatment—A brief review. Croat. J. Food Sci. Technol. 2010, 2, 34–47. [Google Scholar]
- Justino, C.I.; Duarte, K.; Loureiro, F.; Pereira, R.; Antunes, S.C.; Marques, S.M.; Goncalves, F.; Rocha-Santos, T.A.P.; Freitas, A.C. Toxicity and organic content characterization of olive oil mill wastewater undergoing a sequential treatment with fungi and photo-fenton oxidation. J. Hazard. Mater. 2009, 172, 1560–1572. [Google Scholar] [CrossRef] [PubMed]
- Quintero, J.C.; Lu-Chau, T.A.; Moreira, M.T.; Feijoo, G.; Lema, J.M. Bioremediation of hch present in soil by the white-rot fungus bjerkandera adusta in a slurry batch bioreactor. Int. Biodeter. Biodegr. 2007, 60, 319–326. [Google Scholar] [CrossRef]
- Faraco, V.; Pezzella, C.; Miele, A.; Giardina, P.; Sannia, G. Bio-remediation of colored industrial wastewaters by the white-rot fungi Phanerochaete chrysosporium and Pleurotus ostreatus and their enzymes. Biodegradation 2009, 20, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, S.; Khanal, S.K.; Jasti, N.; Jin, B.; Pometto, A.L.; Van Leeuwen, J. Use of filamentous fungi for wastewater treatment and production of high value fungal byproducts: A review. Crit. Rev. Environ. Sci. Technol. 2010, 40, 400–449. [Google Scholar] [CrossRef]
- Kapoor, A.; Viraraghavan, T.; Cullimore, D.R. Removal of heavy metals using the fungus Aspergillus niger. Bioresource Technol. 1999, 70, 95–104. [Google Scholar] [CrossRef]
- Delgado, A.; Anselmo, A.M.; Novais, J.M. Heavy metal biosorption by dried powdered mycelium of fusarium flocciferum. Water Environ. Res. 1998, 70, 370–375. [Google Scholar] [CrossRef]


{kind=link}
| Whole Stillage [42] | Thin Stillage [42] | Wheat Starch Plant Wastes [43] | Potato Protein Liquor [37] | Expired Milk [29] | Vinasse [21] | |
|---|---|---|---|---|---|---|
| pH | 4.3 | 3.5 | 5.2 | 5.3 | 5.5 | 4.3 |
| Total solid (% w/w) | 15.6 | 77.5 | 28.5 | 43.8 | 11 | 2.2 |
| Suspended solid (% w/w) | 8.8 | 26 | 3.7 | 0.9 | - | 1.5 |
| Ash (% w/w) | 3.2 | 10 | 1.9 | 11.4 | - | - |
| Fat | - | - | - | - | 3 | - |
| Crude protein (% w/w) | 32 | 4.4 | 1.2 | 20.2 | 3 | 36.5 |
| Saccharides (g/L)) | 23.1 | 21.7 | 3.8 | 73.9 | 50 | - |
| Acetic acid (g/L) | 0.4 | - | 1 | - | - | 0.8 |
| Glycerol (g/L) | 12 | 7.6 | 0.2 | - | - | 6.8 |
| Lactic acid (g/L) | 1.7 | 11.6 | 1.2 | - | 4.9 | 7.4 |
| Essential Amino Acids | Fishmeal [58] | Soybean Meal [58] | Rendered Meat Meal [58] | Poultry By-Product Meal [58] | Blood Meal [58] | Fungal Biomass [21] |
|---|---|---|---|---|---|---|
| CP% | 64.6 | 47.5 | 54.0 | 64.1 | 77.1 | - |
| Ash% | 15.0 | 6.0 | 25.0 | 19.0 | 4.4 | - |
| Arginine | 3.8 | 3.7 | 6.2 | 6.2 | 4.3 | 3.9 |
| Histidine | 1.5 | 1.2 | 2.1 | 1.9 | 6.6 | 1.3 |
| Isoleucine | 2.7 | 2.1 | 2.9 | 3.1 | 1.2 | 2.0 |
| Leucine | 4.5 | 3.6 | 7.1 | 6.1 | 14.3 | 3.1 |
| Lysine | 4.7 | 3.1 | 5.7 | 5.2 | 9.1 | 2.9 |
| Methionine | 2.3 | 1.4 | 1.5 | 1.7 | 1.3 | 0.6 |
| Phenylalanine | 4.4 | 4.2 | 4.0 | 3.5 | 6.9 | 3.0 |
| Threonine | 2.3 | 1.9 | 3.7 | 3.4 | 5.3 | 2.8 |
| Tryptophan | 0.6 | 0.7 | 0.7 | 1.4 | 1.4 | 0.1 |
| Valine | 2.8 | 2.6 | 4.9 | 3.9 | 9.1 | 2.1 |
| Fatty Acids | Anchovy [69] | Sardine [69] | Herring [69] | Linseed oil [69] | Mucor sp. [70] | Rhizopus sp. [70] | Aspergilus flavus [71] |
|---|---|---|---|---|---|---|---|
| C14:0 | 7.2 | 7.6 | 6.2 | 0.1 | 0.1 | 0.1 | - |
| C16:0 | 17.8 | 16.2 | 12.7 | 6.1 | 8.9 | 9.7 | 14.9 |
| C16:1 | 9.8 | 9.2 | 7.5 | 0.1 | - | - | 0.5 |
| C18:0 | 3.9 | 3.5 | 1.1 | 3.4 | 4.6 | 5.8 | 10.5 |
| C18:1 | 12.0 | 11.4 | 12.9 | 18.8 | 24.5 | 29.6 | 35.8 |
| C18:2n-6 | 1.1 | 1.3 | 1.1 | 16.3 | 60 | 54.3 | 34.3 |
| C18:3n-3 | 0.8 | 0.9 | 0.7 | 54.4 | 2 | - | 0.8 |
| C18:4n-3 | 2.4 | 2.0 | 1.4 | - | - | 0.5 | - |
| C20:1 | 1.9 | 3.2 | 15.1 | - | - | - | 0.6 |
| C20:4n-6 | 0.3 | 1.6 | 0.3 | - | - | - | - |
| C20:5n-3 | 18.3 | 16.9 | 6.8 | - | - | - | - |
| C22:1 | 1.4 | 3.8 | 22.0 | - | - | - | - |
| C22:5n-3 | 1.5 | 2.5 | 0.8 | - | - | - | - |
| C22:6n-3 | 8.5 | 21.9 | 5.8 | - | - | - | - |
| ∑n-3 | 31.5 | 44.2 | 15.5 | 54.4 | - | - | - |
| ∑n-6 | 1.4 | 2.9 | 1.4 | 16.3 | - | - | - |
| n-3/n-6 | 22.5 | 15.2 | 11 | 3.3 | - | - | - |
| Fungal Strain | Substrate | Protein Content (% w/w Dry Weight) | Ref. |
|---|---|---|---|
| Aspergillus oryzae | Palm oil waste | 39 | [88] |
| Rhizopus oligosporus | Starch processing wastewater | 46 | [89] |
| Rhizopus sp. | Spent sulphite liquor | 50–60 | [90] |
| Mucor circinelloides | Corn ethanol stillage | 30.4 | [91] |
| Pythium irregulare | Corn ethanol stillage | 28 | [30] |
| Rhizopus, Mucor, Rhizomucor | Isolated from Tempe | 47–63 | [92] |
| Mucor indicus Rhizopus sp. | Spent sulphite liquor | 30–50 | [10] |
| Rizhopus oryzae | Vinasse | 49.7 | [21] |
| Neurospora intermedia Aspergillus oryzae Rhizopus sp. | Thin stillage | 56 48 55 | [93] |
| Rhizopus oligosporus | Corn ethanol stillage | 43 | [94] |
| Aspergillus oryzae Neurospora intermedia | Stillage | 43 | [95] |
| Rhizopus oligosporus | Wheat bran | 40 | [96] |
| Neurospora intermedia Aspergillus oryzae | Dairy waste | 40 | [29] |
| Neurospora intermedia | Lignocellulose | 50 | [20] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karimi, S.; Mahboobi Soofiani, N.; Mahboubi, A.; Taherzadeh, M.J. Use of Organic Wastes and Industrial By-Products to Produce Filamentous Fungi with Potential as Aqua-Feed Ingredients. Sustainability 2018, 10, 3296. https://doi.org/10.3390/su10093296
Karimi S, Mahboobi Soofiani N, Mahboubi A, Taherzadeh MJ. Use of Organic Wastes and Industrial By-Products to Produce Filamentous Fungi with Potential as Aqua-Feed Ingredients. Sustainability. 2018; 10(9):3296. https://doi.org/10.3390/su10093296
Chicago/Turabian StyleKarimi, Sajjad, Nasrollah Mahboobi Soofiani, Amir Mahboubi, and Mohammad J. Taherzadeh. 2018. "Use of Organic Wastes and Industrial By-Products to Produce Filamentous Fungi with Potential as Aqua-Feed Ingredients" Sustainability 10, no. 9: 3296. https://doi.org/10.3390/su10093296
APA StyleKarimi, S., Mahboobi Soofiani, N., Mahboubi, A., & Taherzadeh, M. J. (2018). Use of Organic Wastes and Industrial By-Products to Produce Filamentous Fungi with Potential as Aqua-Feed Ingredients. Sustainability, 10(9), 3296. https://doi.org/10.3390/su10093296