Heterogeneous Winter Wheat Populations Differ in Yield Stability Depending on their Genetic Background and Management System


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material and Field Site
2.2. Stability Parameters and Statistical Analysis
3. Results
3.1. Weather Data
3.2. Organic CCPs and Reference Varieties
3.3. Conventional CCPs and Reference Variety
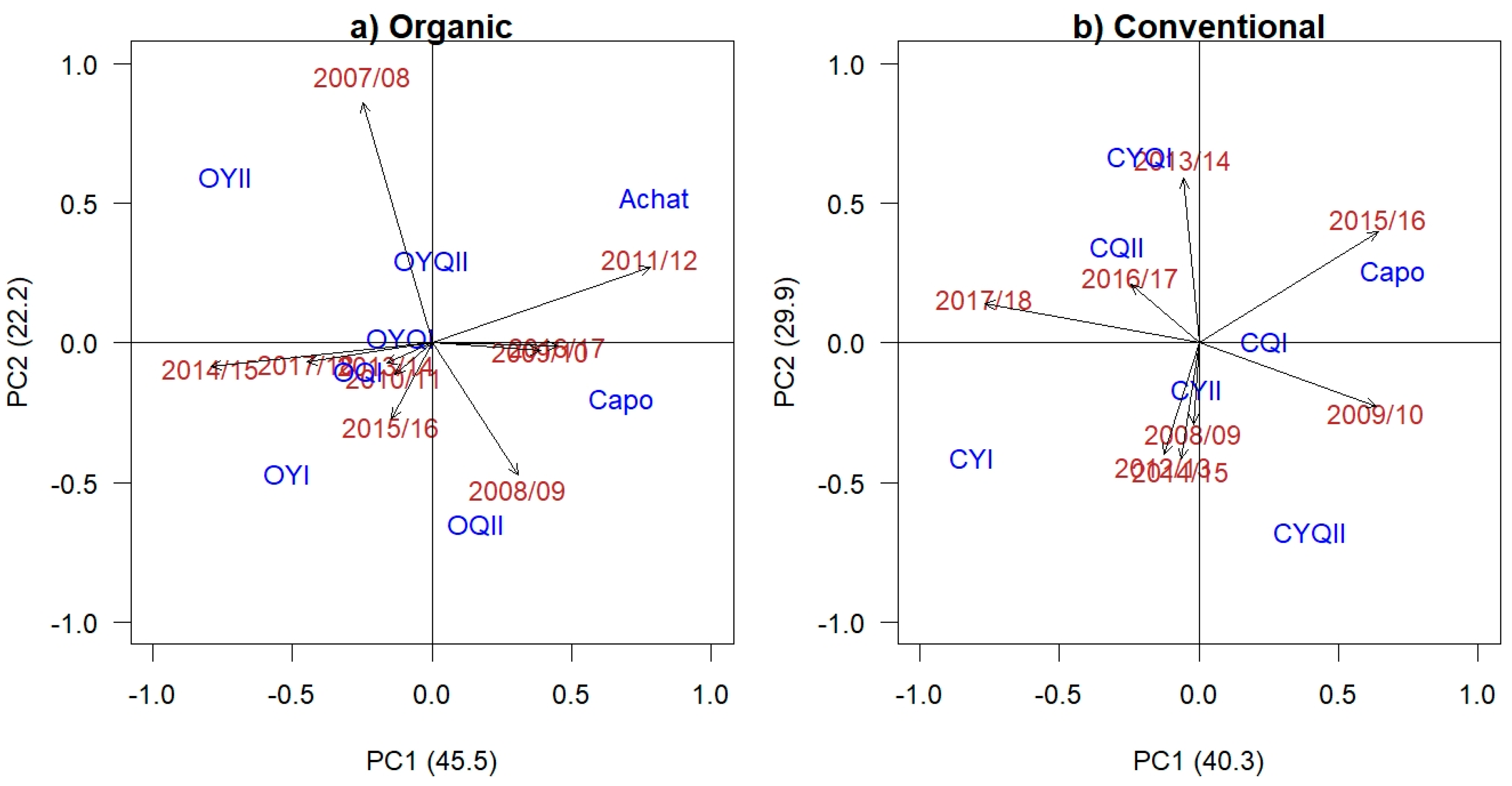
3.4. Association between Stability Parameters under Differing Management Systems
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Experimental Season | Sept | Oct | Nov | Dec | Jan | Feb | Mar | April | May | June | July | Aug | Mean | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | °C | mm | |
| 2005/2006 | 14.9 | 28 | 11.3 | 20 | 5.2 | 38 | 1.8 | 56 | −2.4 | 21 | −0.5 | 40 | 1.5 | 56 | 8.1 | 38 | 12.7 | 84 | 16.3 | 28 | 21.2 | 59 | 15.6 | 73 | 8.8 | 541 |
| 2006/2007 | 16.8 | 18 | 12.7 | 49 | 7.8 | 41 | 5.2 | 48 | 5.3 | 99 | 4.4 | 57 | 6.5 | 59 | 10.7 | 5 | 13.8 | 103 | 17.2 | 117 | 17.2 | 77 | 16.4 | 78 | 11.2 | 751 |
| 2007/2008 | 12.8 | 128 | 8.5 | 23 | 4.0 | 106 | 1.7 | 69 | 4.5 | 70 | 4.0 | 25 | 4.4 | 82 | 7.7 | 73 | 14.5 | 16 | 17.1 | 91 | 18.3 | 56 | 17.9 | 46 | 9.6 | 785 |
| 2008/2009 | 12.7 | 48 | 9.2 | 66 | 5.2 | 54 | 0.3 | 61 | −2.4 | 11 | 1.1 | 33 | 4.5 | 75 | 12.3 | 33 | 13.9 | 78 | 14.7 | 51 | 18.6 | 81 | 18.6 | 69 | 9.1 | 660 |
| 2009/2010 | 14.6 | 77 | 8.3 | 67 | 8.0 | 81 | 0.3 | 96 | −3.8 | 10 | −0.3 | 42 | 4.6 | 71 | 9.2 | 19 | 10.6 | 89 | 16.4 | 46 | 20.7 | 48 | 16.7 | 147 | 8.8 | 793 |
| 2010/2011 | 12.5 | 86 | 8.3 | 29 | 4.8 | 87 | −4.1 | 43 | 1.3 | 49 | 0.4 | 29 | 3.5 | 9 | 12.7 | 37 | 15.6 | 18 | 17.3 | 90 | 16.5 | 43 | 18.4 | 105 | 8.9 | 625 |
| 2011/2012 | 15.4 | 41 | 9.8 | 42 | 4.8 | 2 | 4.1 | 111 | 2.4 | 121 | −2.3 | 24 | 7.5 | 15 | 8.4 | 35 | 14.6 | 61 | 14.9 | 127 | 17.3 | 140 | 18.9 | 59 | 9.7 | 778 |
| 2012/2013 | 13.8 | 44 | 8.8 | 42 | 5.2 | 33 | 2.1 | 90 | −0.1 | 53 | −0.6 | 51 | −0.6 | 25 | 8.4 | 33 | 11.6 | 146 | 15.2 | 26 | 19.0 | 36 | 18.4 | 37 | 8.4 | 616 |
| 2013/2014 | 13.8 | 62 | 11.2 | 81 | 4.8 | 72 | 4.8 | 37 | 2.8 | 38 | 5.3 | 16 | 7.3 | 11 | 11.5 | 29 | 12.4 | 96 | 15.2 | 37 | 19.0 | 125 | 16.0 | 79 | 10.3 | 683 |
| 2014/2015 | 15.0 | 37 | 12.3 | 45 | 7.0 | 15 | 2.7 | 36 | 2.7 | 44 | 1.3 | 18 | 5.1 | 50 | 8.6 | 36 | 12.3 | 25 | 15.5 | 28 | 19.0 | 95 | 19.6 | 160 | 10.1 | 589 |
| 2015/2016 | 12.8 | 51 | 8.5 | 43 | 8.8 | 89 | 7.6 | 29 | 1.7 | 40 | 3.5 | 83 | 4.2 | 45 | 8.0 | 47 | 13.8 | 42 | 17.0 | 99 | 18.6 | 65 | 18.0 | 24 | 10.2 | 657 |
| 2016/2017 | 17.5 | 21 | 8.9 | 71 | 4.2 | 33 | 2.4 | 16 | −2.1 | 36 | 3.4 | 37 | 7.8 | 39 | 7.5 | 37 | 14.4 | 32 | 17.5 | 60 | 18.1 | 183 | 17.7 | 124 | 9.8 | 689 |
| 2017/2018 | 13.0 | 35 | 11.9 | 61 | 5.8 | 73 | 3.6 | 47 | 4.2 | 80 | −1.8 | 9 | 2.7 | 43 | 12.8 | 29 | 15.6 | 33 | 17.6 | 15 | 20.6 | 15 | 20.2 | 31 | 10.5 | 471 |
| Mean | 14.3 | 52 | 10.0 | 49 | 5.8 | 56 | 2.5 | 57 | 1.0 | 52 | 1.4 | 36 | 4.5 | 45 | 9.7 | 35 | 13.5 | 63 | 16.3 | 63 | 18.8 | 79 | 17.9 | 79 | 9.6 | 664 |
| Long-term mean (1971–2000) | 13.6 | 52 | 9.2 | 44 | 4.7 | 51 | 2.3 | 58 | 1.0 | 49 | 1.4 | 36 | 4.9 | 49 | 8.1 | 43 | 12.9 | 58 | 15.5 | 74 | 17.4 | 59 | 17.3 | 55 | 9.0 | 628 |
| Df | SS | % of Model SS | % of GEI SS | |
|---|---|---|---|---|
| Model | 77 | 201.4 | ||
| Env. (E) | 9 | 185.5 | 92.1 | |
| Rep(Env) | 8 | 1.9 | 0.9 | |
| Entry (G) | 6 | 2.9 | 1.4 | |
| GEI | 54 | 11.1 | 5.5 | 5.5 |
| (a) Organic CCPs and reference varieties | ||||||||||||||||||||
| Entries | 2007/2008 | 2008/2009 | 2009/2010 | 2010/2011 | 2011/2012 | 2013/2014 | 2014/2015 | 2015/2016 | 2016/2017 | 2017/2018 | ||||||||||
| Achat | 6.7 | ab | 5.3 | 6.5 | a | 7.4 | 5.2 | a | 2.9 | 4.2 | 4.3 | ab | 6.4 | 4.3 | cd | |||||
| Capo | 5.7 | ab | 5.5 | 6.6 | a | 7.3 | 4.8 | ab | 3.5 | 4.2 | 3.9 | c | 5.9 | 4.3 | cd | |||||
| OQI | 5.6 | ab | 4.8 | 5.1 | b | 7.2 | 4.2 | bc | 2.6 | 4.8 | 3.9 | c | 5.4 | 4.4 | c | |||||
| OQII | 5.3 | b | 5.2 | 6.1 | ab | 7.4 | 3.9 | cd | 2.7 | 4.6 | 4.4 | a | 5.7 | 4.0 | e | |||||
| OYI | 5.8 | ab | 5.2 | 5.9 | ab | 7.2 | 3.4 | d | 3.2 | 5.0 | 4.3 | abc | 5.5 | 4.7 | b | |||||
| OYII | 7.0 | a | 4.7 | 6.1 | ab | 7.5 | 4.0 | cd | 3.4 | 5.4 | 4.2 | abc | 5.4 | 4.9 | a | |||||
| OYQI | 6.3 | ab | 4.9 | 6.1 | ab | 7.8 | 3.9 | cd | 3.4 | 4.3 | 4.6 | a | 5.6 | 4.3 | c | |||||
| OYQII | 6.5 | ab | 5.0 | 6.4 | a | 7.5 | 4.3 | bc | 3.3 | 5.1 | 4.0 | bc | 6.1 | 4.2 | d | |||||
| (b) Conventional CCPs and Reference Variety | ||||||||||||||||||||
| Entries | 2008/2009 | 2009/2010 | 2012/2013 | 2013/2014 | 2014/2015 | 2015/2016 | 2016/2017 | 2017/2018 | ||||||||||||
| Capo | 5.9 | a | 5.1 | a | 6.2 | 5.5 | a | 6.6 | a | 5.4 | a | 6.9 | a | 4.4 | d | |||||
| CQI | 5.0 | cd | 4.3 | bc | 5.4 | 4.6 | cd | 6.0 | ab | 4.4 | b | 5.6 | c | 4.5 | cd | |||||
| CQII | 4.9 | d | 3.9 | c | 5.5 | 4.5 | cd | 6.0 | ab | 4.6 | b | 6.3 | b | 5.1 | ab | |||||
| CYI | 5.6 | abc | 3.9 | c | 6.1 | 4.7 | cd | 6.4 | ab | 3.6 | c | 6.4 | ab | 5.1 | abc | |||||
| CYII | 5.6 | ab | 4.8 | ab | 6.0 | 4.9 | bc | 6.3 | ab | 4.4 | b | 6.4 | ab | 5.0 | abc | |||||
| CYQI | 5.3 | bcd | 4.7 | ab | 5.7 | 5.4 | ab | 5.7 | b | 4.5 | b | 6.7 | ab | 5.4 | a | |||||
| CYQII | 5.6 | abc | 5.2 | a | 6.2 | 4.2 | d | 6.3 | ab | 4.5 | b | 6.2 | b | 4.7 | bcd | |||||
References
- Østergård, H.; Finckh, M.R.; Fontaine, L.; Goldringer, I.; Hoad, S.P.; Kristensen, K.; Lammerts van Bueren, E.T.; Mascher, F.; Munk, L.; Wolfe, M.S. Time for a shift in crop production: Embracing complexity through diversity at all levels. J. Sci. Food Agric. 2009, 89, 1439–1445. [Google Scholar] [CrossRef]
- Döring, T.F.; Knapp, S.; Kovacs, G.; Murphy, K.; Wolfe, M.S. Evolutionary plant breeding in cereals-into a new era. Sustainability 2011, 3, 1944–1971. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Myers, J.R. Organic Crop Breeding: Integrating Organic Agricultural Approaches and Traditional and Modern Plant Breeding Methods. In Organic Crop Breeding; Lammerts van Bueren, E.T., Myers, J.R., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 1–13. [Google Scholar]
- Akcura, M.; Kaya, Y.; Taner, S.; Ayranci, R. Parametric stability analyses for grain yield of durum wheat. Plant Soil Environ. 2006, 52, 254–261. [Google Scholar] [CrossRef]
- Kahiluoto, H.; Kaseva, J.; Balek, J.; Olesen, J.E.; Ruiz-Ramos, M.; Gobin, A.; Kersebaum, K.C.; Takáč, J.; Ruget, F.; Ferrise, R.; et al. Decline in climate resilience of European wheat. Proc. Natl. Acad. Sci. USA 2019, 116, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Piepho, H.P. Recent claim of declining climate resilience in European wheat is not supported by the statistics used. Proc. Natl. Acad. Sci. USA 2019, 166, 10625–10626. [Google Scholar] [CrossRef] [PubMed]
- Snowdon, R.J.; Stahl, A.; Wittkop, B.; Friedt, W.; Voss-Fels, K.; Ordon, F.; Frisch, M.; Dreisigacker, S.; Hearne, S.J.; Bett, K.E.; et al. Reduced response diversity does not negatively impact wheat climate resilience. Proc. Natl. Acad. Sci. USA 2019, 166, 10623–10624. [Google Scholar] [CrossRef] [PubMed]
- Kahiluoto, H.; Kaseva, J.; Olesen, J.E.; Kersebaum, K.C.; Ruiz-Ramos, M.; Gobin, A.; Takáč, J.; Ruget, F.; Ferrise, R.; Balek, J.; et al. Genetic response diversity to provide yield stability of cultivar groups deserves attention. Proc. Natl. Acad. Sci. USA 2019, 166, 10627–10629. [Google Scholar] [CrossRef] [PubMed]
- Tooker, J.F.; Frank, S.D. Genotypically diverse cultivar mixtures for insect pest management and increased crop yields. J. Appl. Ecol. 2012, 49, 974–985. [Google Scholar] [CrossRef]
- Finckh, M.R.; Wolfe, M.S. Biodiversity enhancement. In Plant Diseases and Their Management in Organic Agriculture; Finckh, M.R., van Bruggen, A.H.C., Tamm, L., Eds.; APS Press: St. Paul, MN, USA, 2015; pp. 153–174. [Google Scholar]
- Vidal, T.; Boixel, A.L.; Durand, B.; de Vallavieille-Pope, C.; Huber, L.; Saint-Jean, S. Reduction of fungal disease spread in cultivar mixtures: Impact of canopy architecture on rain-splash dispersal and on crop microclimate. Agric. For. Meteorol. 2017, 246, 154–161. [Google Scholar] [CrossRef]
- European Commission. Commission implementing decision of 18 March 2014 on the organisation of a temporary experiment providing for certain derogations for the marketing of plant populations of the plant species wheat, barley, oats and maize pursuant to council directive 66/40. Off. J. Eur. Union. 2014, 82, 29–36. [Google Scholar]
- European Commission. Commission implementing decision of of 9 October 2018 amending Implementing Decision 2014/150/EU on the organisation of a temporary experiment providing for certain derogations for the marketing of populations of the plant species wheat, barley, oats and maize pursuant to Council Directive 66/402/EEC. Off. J. Eur. Union. 2018, 256, 65–66. [Google Scholar]
- Annicchiarico, P.; Filippi, L. A field pea ideotype for organic systems of northern Italy. J. Crop Improv. 2007, 20, 193–203. [Google Scholar] [CrossRef]
- Döring, T.F.; Bocci, R.; Hitchings, R.; Howlett, S.; Lammerts van Bueren, E.T.; Pautasso, M.; Raaijmakers, M.; Rey, F.; Stubsgaard, A.; Weinhappel, M.; et al. The organic seed regulations framework in Europe-current status and recommendations for future development. Org. Agric. 2012, 2, 173–183. [Google Scholar] [CrossRef]
- Dawson, J.C.; Goldringer, I. Breeding for Genetically Diverse Populations: Variety Mixtures and Evolutionary Populations. In Organic Crop Breeding; Lammerts van Bueren, E.T., Myer, J.R., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 77–98. [Google Scholar]
- Annicchiarico, P. Genotype x Environment Interactions: Challenges and Opportunities for Plant Breeding and Cultivar Recommendations, Vol. 174; Food & Agriculture Organization: Rome, Italy, 2002; Volume 174. [Google Scholar]
- Lammerts Van Bueren, E.T.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS Wagening. J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Raggi, L.; Ciancaleoni, S.; Torricelli, R.; Terzi, V.; Ceccarelli, S.; Negri, V. Evolutionary breeding for sustainable agriculture: Selection and multi-environmental evaluation of barley populations and lines. Field Crop. Res. 2017, 204, 76–88. [Google Scholar] [CrossRef]
- Becker, H.C.; Leon, J. Stability Analysis in Plant Breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Lin, C.S.; Binns, M.R.; Lefkovitch, L.P. Stability Analysis: Where Do We Stand? Crop. Sci. 1986, 26, 894–900. [Google Scholar] [CrossRef]
- Stagnari, F.; Onofri, A.; Codianni, P.; Pisante, M. Durum wheat varieties in N-deficient environments and organic farming: A comparison of yield, quality and stability performances. Plant Breed. 2013, 132, 266–275. [Google Scholar] [CrossRef]
- Simmonds, N. Selection for local adaptation in a plant breeding programme. Theor. Appl. Genet. 1991, 82, 363–367. [Google Scholar] [CrossRef]
- Ceccarelli, S. Positive interpretation of genotype by environment interactions in relation to sustainability and biodiversity. In Plant Adaptation and Crop Improvement; Cooper, M., Hammer, G.L., Eds.; CAB International in Association with IRRI and ICRISAT: Oxford, UK, 1996; pp. 467–486. [Google Scholar]
- Cleveland, D. Is plant breeding science objective truth or social construction? The case of yield stability. Agric. Hum. Values 2001, 18, 251–270. [Google Scholar] [CrossRef]
- Frankel, O.H.; Brown, A.H.D.; Burdon, J.J. The Conservation of Plant Biodiversity; Cambridge University Press: Cambridge, UK, 1995; pp. 1–301. [Google Scholar]
- Eberhart, S.A.; Russell, W.A. Stability parameters for comparing varieties. Crop. Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability as influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Finckh, M.R.; Gacek, E.; Goyeau, H.; Lannou, C.; Merz, U.; Mundt, C.; Munk, L.; Nadziak, J.; Newton, A.; de Vallavieille-Pope, C.; et al. Cereal variety and species mixtures in practice, with emphasis on disease resistance. Agronomie 2000, 20, 813–837. [Google Scholar] [CrossRef] [Green Version]
- Moreau, M. Linking biodiversity and ecosystems. Philos. Trans. R. Soc. B 2010, 365, 49–60. [Google Scholar]
- Newton, A.C.; Gravouil, C.; Fountaine, J.M. Managing the ecology of foliar pathogens: Ecological tolerance in crops. Ann. Appl. Biol. 2010, 157, 343–359. [Google Scholar] [CrossRef]
- Barot, S.; Allard, V.; Cantarel, A.; Enjalbert, J.; Gauffreteau, A.; Goldringer, I.; Lata, J.C.; Le Roux, X.; Niboyet, A.; Porcher, E. Designing mixtures of varieties for multifunctional agriculture with the help of ecology. Rev. Agron. Sustain. Dev. 2017, 37, 13. [Google Scholar] [CrossRef]
- Döring, T.F.; Wolfe, M.; Jones, H.; Pearce, H.; Zhan, J. Breeding for resilience in wheat—Nature’s choice. In Proceedings of the Breeding for Resilience: A Strategy for Organic and Low-Input Farming Sytems? Eucarpia 2nd Conference of the Organic and Low-Input Agriculture Section, Paris, France, 1–3 December 2010; Goldringer, I., Ed.; pp. 45–48. Available online: http://www.researchgate.net/publication/255723956_Breeding_for_resilience_in_wheat_-_Natures_choice (accessed on 31 October 2019).
- Hockett, E.A.; Eslick, R.F.; Qualset, C.O.; Dubbs, A.L.; Stewart, V.R. Effects of natural selection in advanced generations of Barley composite cross II. Crop. Sci. 1983, 23, 752–756. [Google Scholar] [CrossRef]
- Clay, R.E.; Allard, R.W. A comparison of the performance of homogenous and heterogenous barley populations. Crop. Sci. 1969, 9, 407–412. [Google Scholar] [CrossRef]
- Soliman, K.M.; Allard, R.W. Grain Yield of Composite Cross Populations of Barley: Effects of Natural Selection. Crop. Sci. 1991, 31, 705–708. [Google Scholar] [CrossRef]
- Grando, S.; von Bothmer, R.; Ceccarelli, S. Genetic Diversity of Barley: Use of Locally Adapted Germplam to Enhance Yield and Yield Stability of Barley in Dry Areas. In Broadening the Genetic Base of Crop Production; Cooper, H.D., Spillane, C., Hodgkin, T., Eds.; CABI Publishing: Wallinford, UK, 2001; pp. 351–451. [Google Scholar]
- Creissen, H.E.; Jorgensen, T.H.; Brown, J.K.M. Increased yield stability of field-grown winter barley (Hordeum vulgare L.) varietal mixtures through ecological processes. Crop. Prot. 2016, 85, 1–8. [Google Scholar] [CrossRef]
- Dubin, H.J.; Wolfe, M.S. Comparative behavior of three wheat cultivars and their mixture in India, Nepal and Pakistan. Feild Crop. Res. 1994, 39, 71–83. [Google Scholar] [CrossRef]
- Döring, T.F.; Annicchiarico, P.; Clarke, S.; Haigh, Z.; Jones, H.E.; Pearce, H.; Snape, J.; Zhan, J.; Wolfe, M.S. Comparative analysis of performance and stability among composite cross populations, variety mixtures and pure lines of winter wheat in organic and conventional cropping systems. Feild Crop. Res. 2015, 183, 235–245. [Google Scholar] [CrossRef]
- Gaudin, A.C.M.; Tolhurst, T.N.; Ker, A.P.; Janovicek, K.; Tortora, C.; Martin, R.C.; Deen, W. Increasing Crop Diversity Mitigates Weather Variations and Improves Yield Stability. PLoS ONE 2015, 10, e0113261. [Google Scholar] [CrossRef] [PubMed]
- Manns, H.R.; Martin, R.C. Cropping system yield stability in response to plant diversity and soil organic carbon in temperate ecosystems. Agroecol. Sustain. Food Syst. 2018, 42, 724–750. [Google Scholar] [CrossRef]
- Brumlop, S.; Pfeiffer, T.; Finckh, M. Evolutionary Effects on Morphology and Agronomic Performance of Three Winter Wheat Composite Cross Populations Maintained for Six Years under Organic and Conventional Conditions. Org. Farming 2017, 3, 34–50. [Google Scholar] [CrossRef]
- Brumlop, S.; Weedon, O.D.; Link, W.; Finckh, M.R. Effective population size (Ne) of organically and conventionally grown composite cross winter wheat populations depending on generation. Eur. J. Agron. 2019, 109. [Google Scholar] [CrossRef]
- Kang, M.S. Using genotype-by-environment interaction for crop cultivar development. Adv. Agron. 1998, 62, 199–252. [Google Scholar]
- Jones, H.; Clarke, S.; Haigh, Z.; Pearce, H.; Wolfe, M. The effect of the year of wheat variety release on productivity and stability of performance on two organic and two non-organic farms. J. Agric. Sci. 2010, 148, 303–317. [Google Scholar] [CrossRef] [Green Version]
- Wildhagen, H. Bodenkundliche Standortbeschreibungen der Versuchsflächen des Fachbereiches; University of Kassel: Witzenhausen, Germany, 1998. [Google Scholar]
- Bertholdsson, N.O.; Weedon, O.; Brumlop, S.; Finckh, M.R. Evolutionary changes of weed competitive traits in winter wheat composite cross populations in organic and conventional farming systems. Eur. J. Agron. 2016, 79, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Vijaya Bhaskar, A.V.; Weedon, O.D.; Finckh, M.R. Exploring the differences between organic and conventional breeding in early vigour traits of winter wheat. Eur. J. Agron. 2019, 105, 86–95. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing [Internet]; R Development Core Team, Ed.; R Foundation for Statistical Computing, 2011; p. 409, (R Foundation for Statistical Computing. Volume 1); Available online: http://www.r-project.org (accessed on 31 October 2019).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S.; Scheipl, F. Lme4: Linear Mixed-Effects Models Using “Eigen” und S4 2019. Available online: https://github.com/lme4/lme4/ (accessed on 31 October 2019).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Heisterkamp, S.; Van Willigen, B. Nlme: Linear and Nonlinear Mixed Effects Models 2018. Available online: https://svn.r-project.org/R-packages/trunk/nlme (accessed on 31 October 2019).
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. emmeans: Estimated Marginal Means, aka Least-Squares Means 2019. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 31 October 2019).
- Zuur, A.; Ieno, E.; Walker, N.; Saveliev, A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R; Springer Science+Business Media: New York, NY, USA, 2009; pp. 1–563. [Google Scholar]
- Roemer, T. Sind die ertragsreichensorten ertragssicherer? Mitt. DLG 1917, 32, 87–89. [Google Scholar]
- Kataoka, S. A stochastic programming model. Econometrika 1963, 31, 181–196. [Google Scholar] [CrossRef]
- Eskridge, K.M. Selection of stable cultivars using a safety-first rule. Crop. Sci. 1990, 30, 369–374. [Google Scholar] [CrossRef]
- Wricke, G. On a method of understanding the biological diversity in field research. Z. Pflanzenzücht 1962, 47, 92–96. [Google Scholar]
- Lin, C.-S.; Binns, M.R. A superiority measure of cultivar performance for cultivar × location data. Can. J. Plant Sci. 1988, 68, 193–198. [Google Scholar] [CrossRef]
- Finlay, K.W.; Wilkinson, G.N. The analysis of adaptation in a plant breeding programme. Aust. J. Agric. Res. 1963, 14, 742–754. [Google Scholar] [CrossRef]
- Gollob, H.F. A statistical model which combines features of factor analytic and analysis of variance techniques. Psychometrika 1968, 33, 73–114. [Google Scholar] [CrossRef]
- Mandel, J. A new analysis of variance model for non-additive data. Technometrics 1971, 13, 1–18. [Google Scholar] [CrossRef]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G. Statistical analysis of yield trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. 2019. Available online: http://tarwi.lamolina.edu.pe/~fmendiburu (accessed on 31 October 2019).
- Farshadfar, E.; Mahmodi, N.; Yaghotipoor, A. AMMI stability value and simultaneous estimation of yield and yield stability in bread wheat (Triticum aestivum L.). Aust. J. Crop. Sci. 2011, 5, 1837–1844. [Google Scholar]
- Purchase, J.L.; Hatting, H.; van Deventer, C.S. Genotype × environment interaction of winter wheat (Triticum aestivum L.) in South Africa: II. Stability analysis of yield performance. S. Afr. J. Plant Soil. 2000, 17, 101–107. [Google Scholar] [CrossRef]
- Kang, M.S. A Rank-Sum method for selecting high-yielding, stable corn genotypes. Cereal Res. Commun. 1988, 16, 113–115. [Google Scholar]
- Vijaya Bhaskar, A.V.; Baresel, J.P.; Weedon, O.D.; Finckh, M.R. Effects of ten years organic and conventional farming on early seedling traits of evolving winter wheat composite cross populations. Sci. Rep. 2019, 9, 1–12. [Google Scholar]
- Goldringer, I.; Enjalbert, J.; David, J.; Paillard, S.; Pham, J.; Brabant, P. Dynamic management of genetic resources: A 13-year experiment on wheat. In Broadening the Genetic Base of Crop Production; Cooper, H., Spillane, C., Hodgkin, T., Eds.; CABI: Wallingford, UK, 2001; pp. 245–260. [Google Scholar]
- Goldringer, I.; Enjalbert, J.; Raquin, A.L.; Brabant, P. Strong selection in wheat populations during ten generations of dynamic management. Genet. Sel. Evol. 2001, 33, 441–463. [Google Scholar] [CrossRef]
- Phillips, S.L.; Wolfe, M.S. Evolutionary plant breeding for low input systems. J. Agric. Sci. 2005, 143, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Creissen, H.E.; Jorgensen, T.H.; Brown, J.K.M. Stabilisation of yield in plant genotype mixtures through compensation rather than complementation. Ann. Bot. 2013, 112, 1439–1447. [Google Scholar] [CrossRef]
- Dawson, J.C.; Huggins, D.R.; Jones, S.S. Characterizing nitrogen use efficiency in natural and agricultural ecosystems to improve the performance of cereal crops in low-input and organic agricultural systems. Field Crop. Res. 2008, 107, 89–101. [Google Scholar] [CrossRef]
- Messmer, M.; Hildermann, I.; Thorup-Kristensen, K.; Rengel, Z. Nutrient Management in Organic Farming and Consequences for Direct and Indirect Selection Strategies. In Organic Crop Breeding; Lammerts van Bueren, E.T., Myers, J.R., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 15–38. [Google Scholar]
- Baresel, J.P.; Zimmermann, G.; Reents, H.J. Effects of genotype and environment on N uptake and N partition in organically grown winter wheat (Triticum aestivum L.) in Germany. Euphytica 2008, 163, 347–354. [Google Scholar] [CrossRef]
- Osman, A.M.; Struik, P.C.; Lammerts van Bueren, E.T. Perspectives to breed for improved baking quality wheat varieties adapted to organic growing conditions. J. Sci. Food Agric. 2012, 92, 207–215. [Google Scholar] [CrossRef]
- Knapp, S.; Snape, J.; Döring, T.; Wolfe, M.S.; Griffiths, S. Genetic analysis of evolving winter wheat populations reveals reversion to wild type. In Proceedings of International Symposium on Evolutionary Breeding in Cereals; Döring, T., Howlett, S., Winkler, L.R., Wolfe, M.S., Eds.; Aston University: Birmingham, UK; The Organic Research Centre: Hamstead Marshall, UK, 2013; p. 10. [Google Scholar]
- Denison, R.F. Darwinian Agriculture. How Understanding Evolution can Improve Agriculture; Princeton University Press: Princeton, NJ, USA, 2012; pp. 1–249. [Google Scholar]
- Duarte, J.B.; Zimmermann, M.J. Correlation among yield stability parameters in common bean. Crop. Sci. 1995, 35, 905–912. [Google Scholar] [CrossRef]
- Temesgen, T.; Keneni, G.; Sefera, T.; Jarso, M. Yield stability and relationships among stability parameters in faba bean (Vicia faba L.) genotypes. Crop. J. 2015, 3, 258–268. [Google Scholar] [CrossRef]
- Kanouni, H.; Farayedi, Y.; Saeid, A. Stability Analyses for Seed Yield of Chickpea (Cicer arietinum L.) Genotypes in the Western Cold Zone of Iran. J. Agric. 2015, 7, 219–230. [Google Scholar] [CrossRef]
- Knapp, S.; Brabant, C.; Oberforster, M.; Grausgruber, H.; Hiltbrunner, J. Quality traits in winter wheat: Comparison of stability parameters and correlations between traits regarding their stability. J. Cereal Sci. 2017, 77, 186–193. [Google Scholar] [CrossRef]

| Entry | GY | EVi | W2 | Ii | bi | MSE | ASV | YSi | Pi (x1000) | Pi genetic | Pi GEI | Pi gen % | Pi GEI % | TOP 3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (a) Organic CCPs and reference varieties | |||||||||||||||
| Achat | 4.85 | ab | 1.98 | 1.85 | 4.35 | 1.07 | 0.22 | 1.71 | 9 | 130.3 | 69.6 | 60.7 | 53 | 47 | 4 |
| Capo | 4.87 | ab | 1.55 | 1.38 | 4.32 | 0.95 | 0.17 | 1.41 | 10 | 224.4 | 130.2 | 94.2 | 58 | 42 | 3 |
| OQI | 4.44 | b | 1.46 | 0.96 | 3.97 | 0.94 | 0.11 | 0.55 | 11 | 448.3 | 393.3 | 55.0 | 88 | 12 | 2 |
| OQII | 4.51 | b | 1.72 | 0.84 | 4.03 | 1.02 | 0.10 | 0.72 | 11 | 380.9 | 284.1 | 96.8 | 75 | 25 | 2 |
| OYI | 4.74 | ab | 1.44 | 1.01 | 4.19 | 0.93 | 0.12 | 1.16 | 11 | 336.8 | 222.7 | 114.1 | 66 | 34 | 1 |
| OYII | 4.93 | a | 1.73 | 1.65 | 4.37 | 1.00 | 0.21 | 1.62 | 9 | 174.1 | 88.1 | 86.0 | 51 | 49 | 4 |
| OYQI | 4.83 | ab | 1.72 | 0.76 | 4.23 | 1.03 | 0.09 | 0.24 | 6 | 244.7 | 153.8 | 90.9 | 63 | 37 | 4 |
| OYQII | 4.83 | ab | 1.81 | 0.48 | 4.32 | 1.07 | 0.05 | 0.30 | 5 | 123.4 | 97.1 | 26.4 | 79 | 21 | 6 |
| (b) Conventional CCPs and reference variety | |||||||||||||||
| Capo | 5.67 | a | 0.65 | 1.16 | 5.22 | 0.94 | 0.19 | 0.97 | 7 | 53.6 | 8.7 | 44.9 | 16 | 84 | 4 |
| CQI | 4.87 | e | 0.41 | 0.32 | 4.54 | 0.84 | 0.04 | 0.31 | 9 | 443.9 | 421.1 | 22.8 | 95 | 5 | 4 |
| CQII | 5.01 | de | 0.63 | 0.65 | 4.56 | 0.99 | 0.11 | 0.53 | 9 | 365.0 | 314.2 | 50.8 | 86 | 14 | 2 |
| CYI | 5.07 | cd | 1.24 | 1.40 | 4.47 | 1.45 | 0.10 | 1.17 | 12 | 401.0 | 227.0 | 174.0 | 57 | 43 | 1 |
| CYII | 5.35 | b | 0.56 | 0.07 | 4.95 | 1.00 | 0.01 | 0.17 | 3 | 126.8 | 95.8 | 31.0 | 76 | 24 | 8 |
| CYQI | 5.37 | b | 0.43 | 0.89 | 4.97 | 0.78 | 0.12 | 0.72 | 7 | 164.0 | 115.5 | 48.5 | 70 | 30 | 5 |
| CYQII | 5.23 | bc | 0.68 | 1.03 | 4.80 | 0.99 | 0.17 | 0.87 | 9 | 235.9 | 141.2 | 94.6 | 60 | 40 | 0 |
| (a) Organic CCPs and Reference Varieties | (b) Conventional CCPs and Reference Variety | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Df | SS | % of Model SS | % of GEI SS | Df | SS | % of Model SS | % of GEI SS | ||
| Model | 87 | 234.7 | Model | 61 | 61.8 | ||||
| Env. (E) | 9 | 214.4 | 91.3 | Env. (E) | 7 | 45.0 | 73.0 | ||
| Rep(Env) | 8 | 1.8 | 0.8 | Rep(Env) | 6 | 0.5 | 0.7 | ||
| Entry (G) | 7 | 4.0 | 1.7 | Entry (G) | 6 | 6.1 | 9.9 | ||
| GEI | 63 | 14.5 | 6.2 | 6.2 | GEI | 42 | 10.1 | 16.4 | 16.4 |
| GY | EVi | W2 | Ii | bi | MSE | ASV | YSi | Pi | |
|---|---|---|---|---|---|---|---|---|---|
| GY | - | −0.65 ** | 0.00 | 0.97 *** | −0.18 | 0.09 | −0.01 | −0.43 | −0.44 |
| EVi | - | 0.43 | −0.65 * | 0.64 ** | 0.30 | 0.35 | 0.11 | −0.17 | |
| W2 | - | −0.08 | 0.17 | 0.85 *** | 0.95 *** | 0.47 | −0.10 | ||
| Ii | - | −0.22 | 0.09 | −0.07 | −0.51 | −0.48 | |||
| bi | - | −0.14 | 0.06 | −0.04 | −0.14 | ||||
| MSE | - | 0.84 *** | 0.23 | −0.30 | |||||
| ASV | - | 0.55 * | −0.08 | ||||||
| YSi | - | 0.72 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weedon, O.D.; Finckh, M.R. Heterogeneous Winter Wheat Populations Differ in Yield Stability Depending on their Genetic Background and Management System. Sustainability 2019, 11, 6172. https://doi.org/10.3390/su11216172
Weedon OD, Finckh MR. Heterogeneous Winter Wheat Populations Differ in Yield Stability Depending on their Genetic Background and Management System. Sustainability. 2019; 11(21):6172. https://doi.org/10.3390/su11216172
Chicago/Turabian StyleWeedon, Odette D., and Maria R. Finckh. 2019. "Heterogeneous Winter Wheat Populations Differ in Yield Stability Depending on their Genetic Background and Management System" Sustainability 11, no. 21: 6172. https://doi.org/10.3390/su11216172
APA StyleWeedon, O. D., & Finckh, M. R. (2019). Heterogeneous Winter Wheat Populations Differ in Yield Stability Depending on their Genetic Background and Management System. Sustainability, 11(21), 6172. https://doi.org/10.3390/su11216172