Quantifying the Distribution and Diversity of Fish Species Along Elevational Gradients in the Weihe River Basin, Northwest China

Abstract
:1. Introduction
2. Materials and Methods
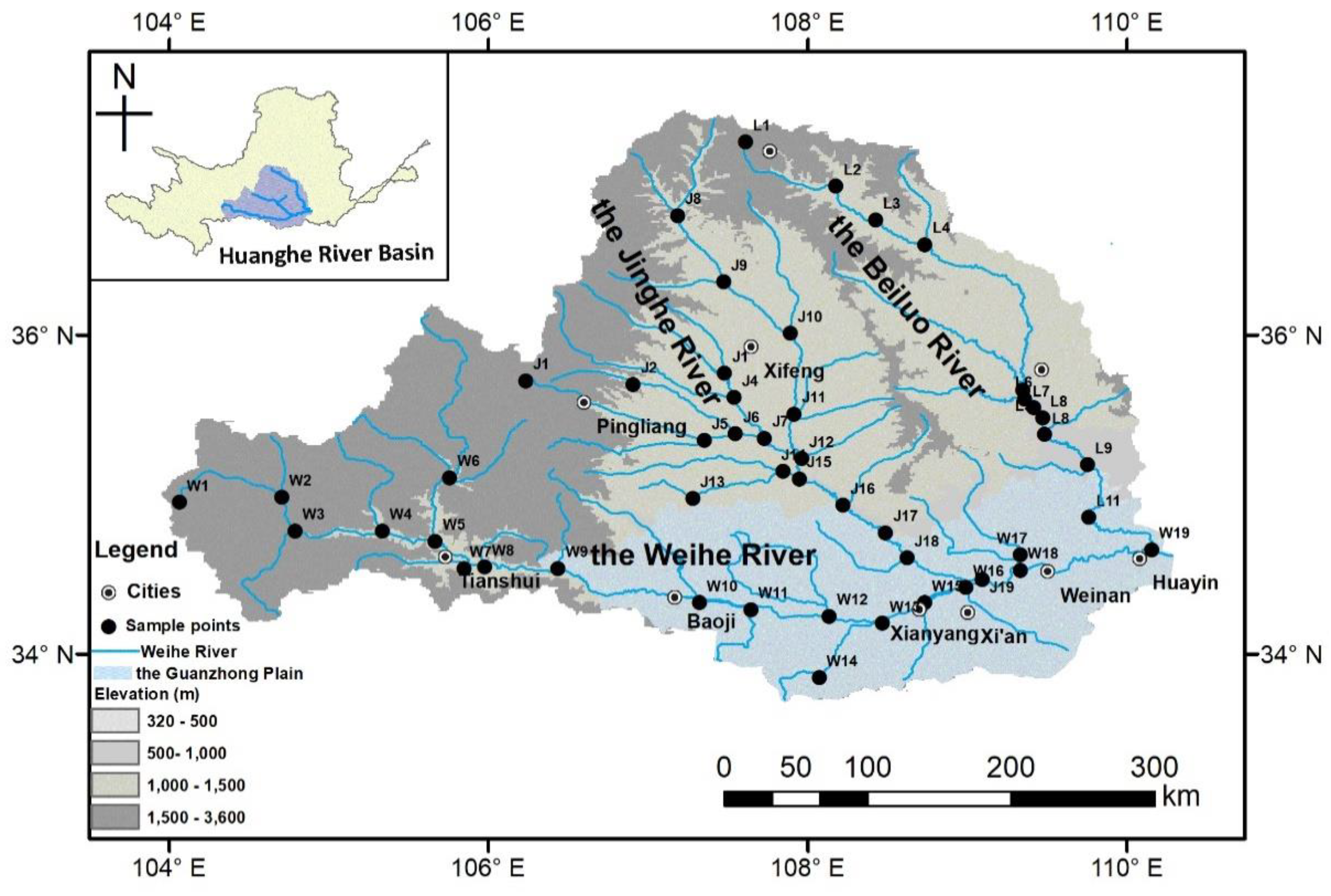
2.1. Study Area
2.2. Fish Sampling
2.3. Environmental Variables
2.4. Data Analysis
3. Results
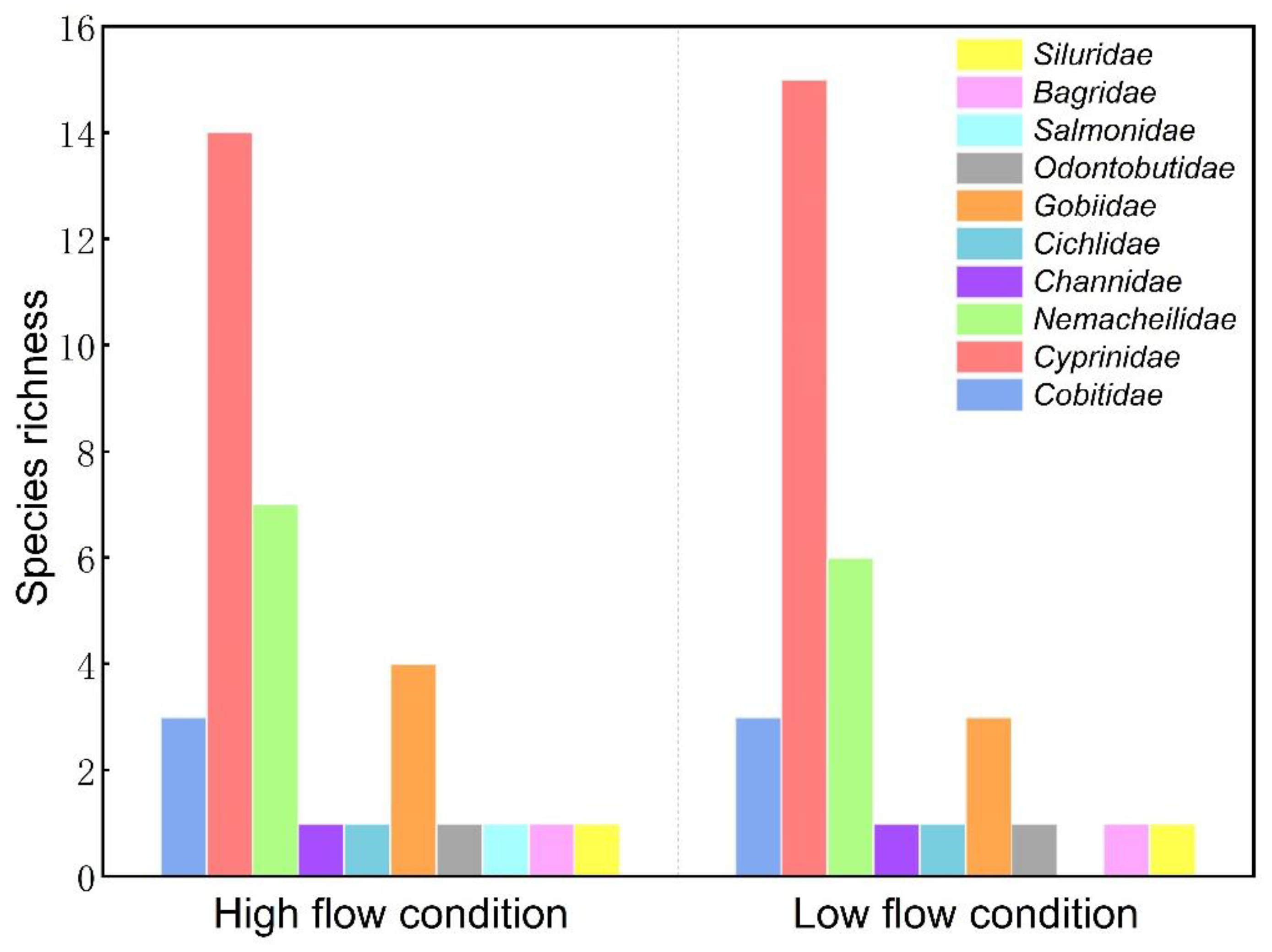
3.1. Overall Fish Species Composition in the Weihe River Basin
3.2. Patterns of Fish Species Diversity and its Distribution
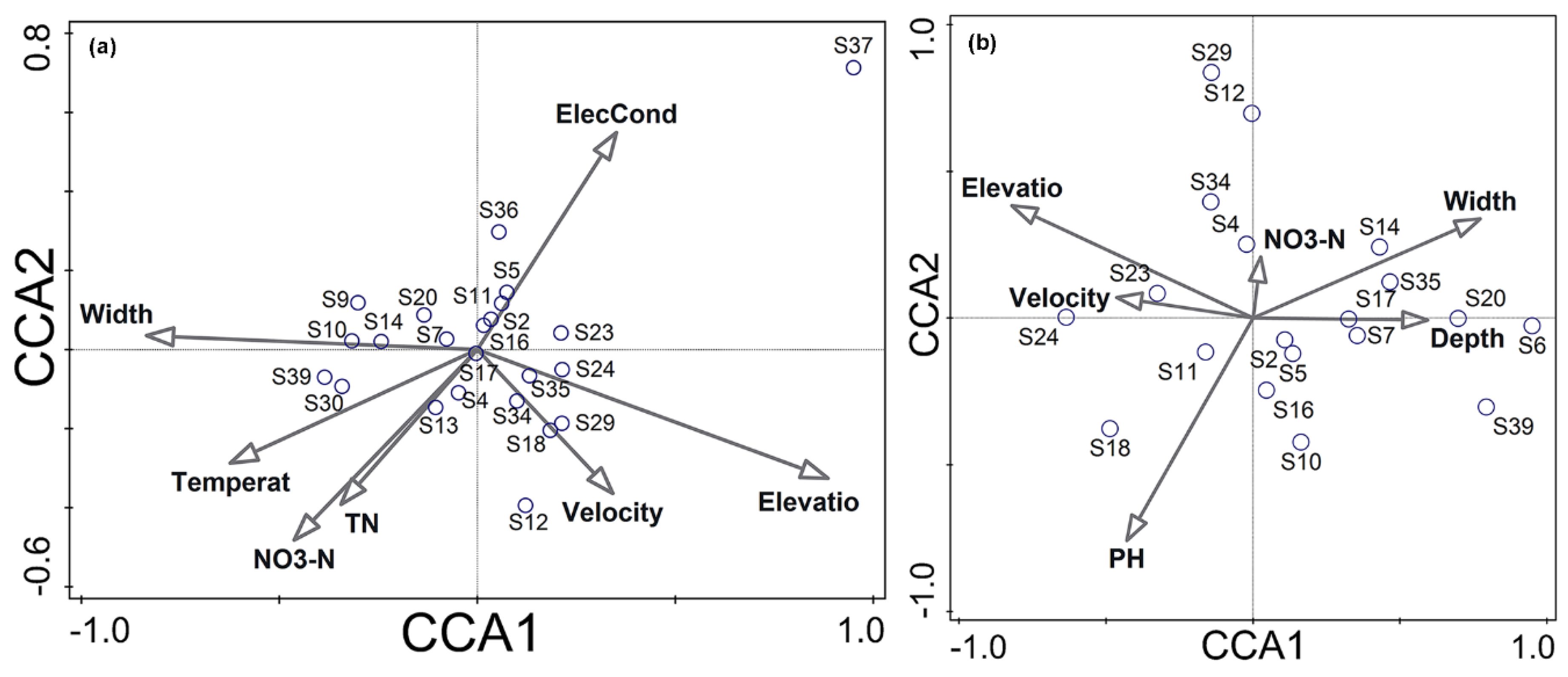
3.3. Relationships between Environmental Variables and Fish Species
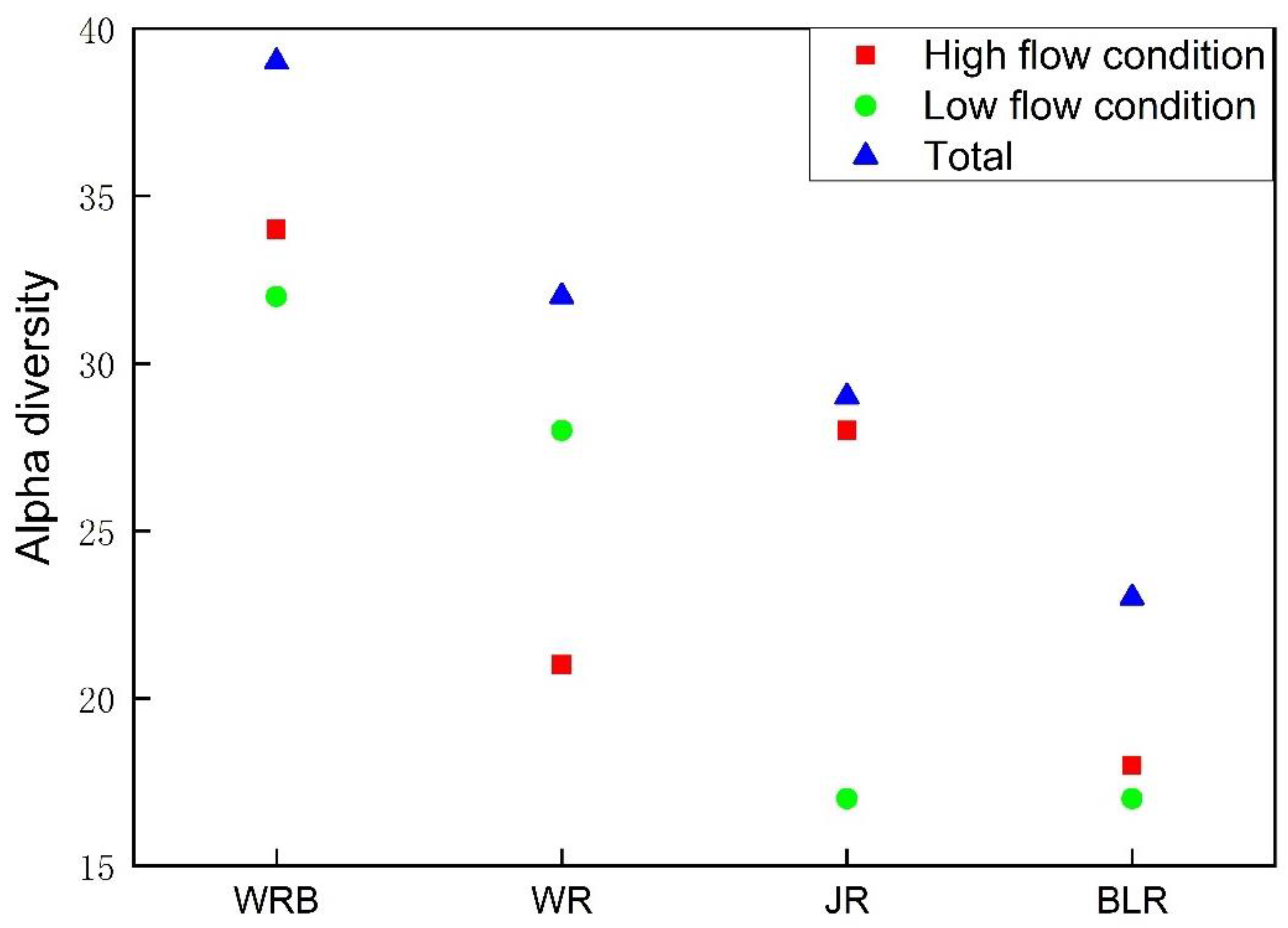
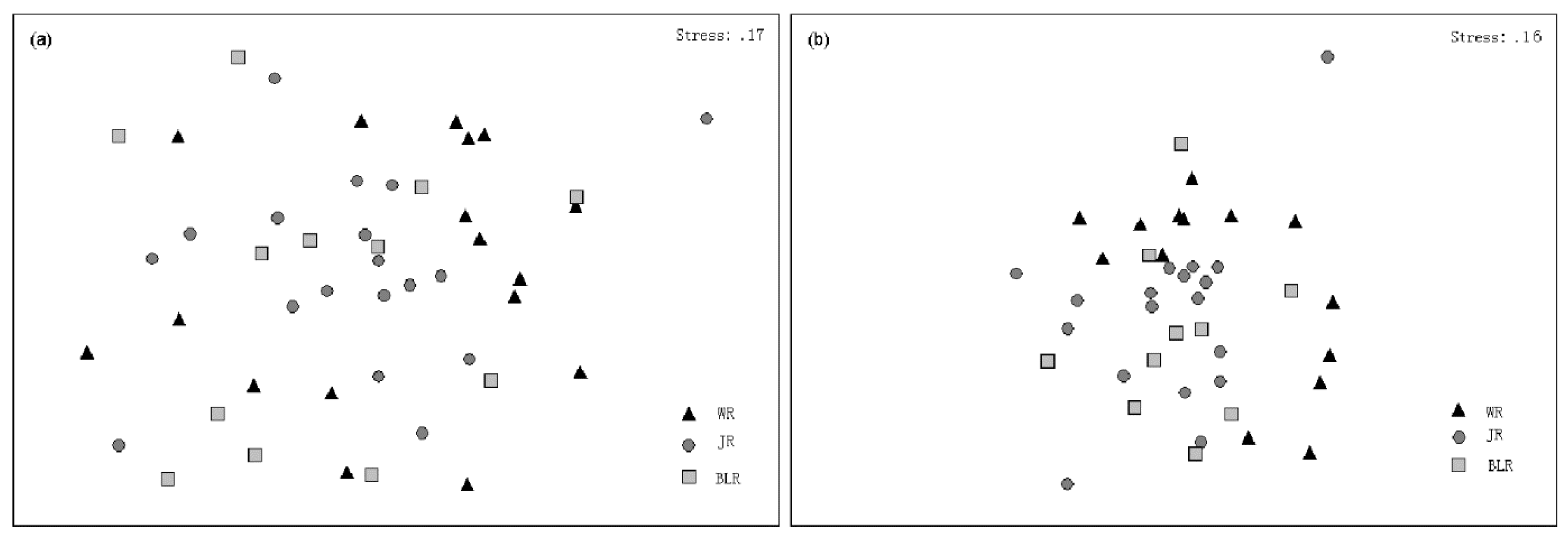
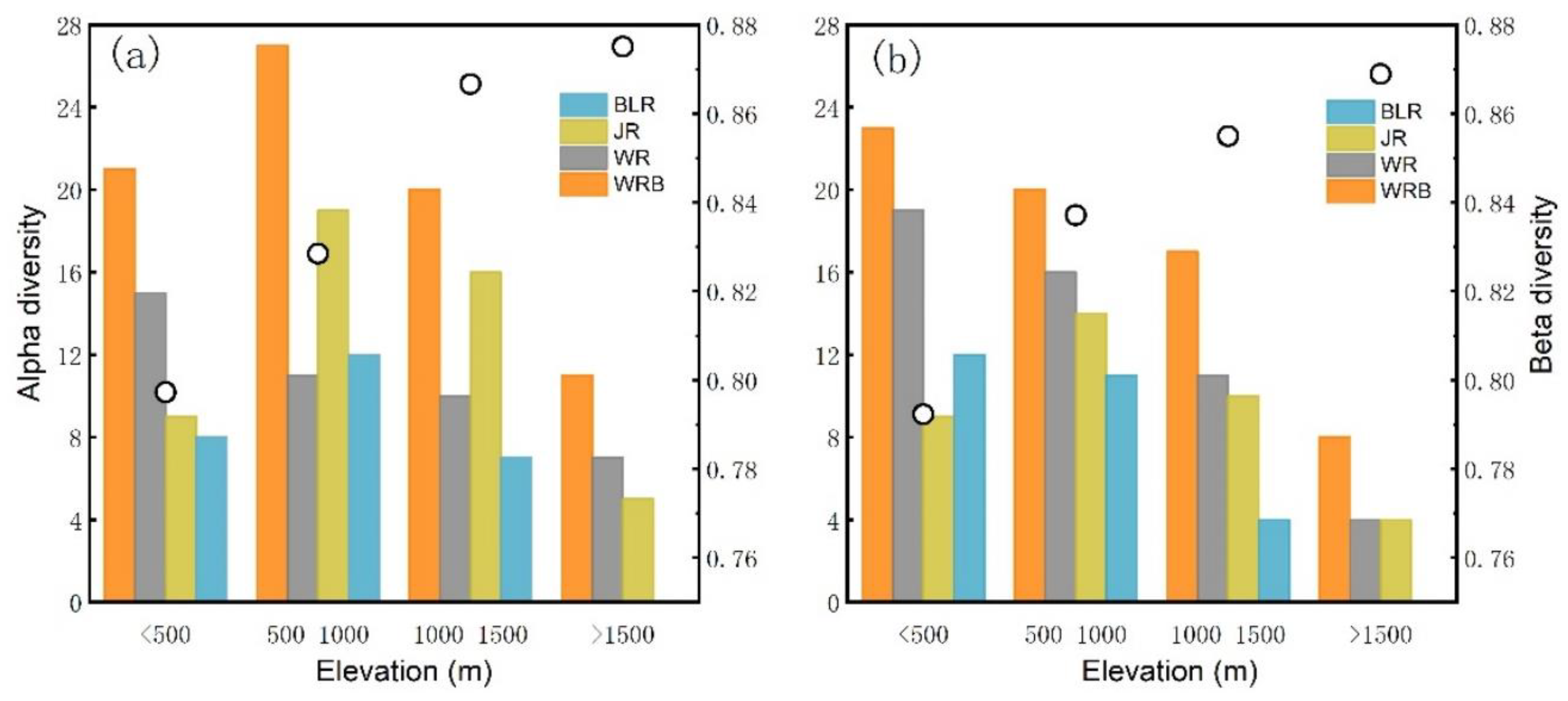
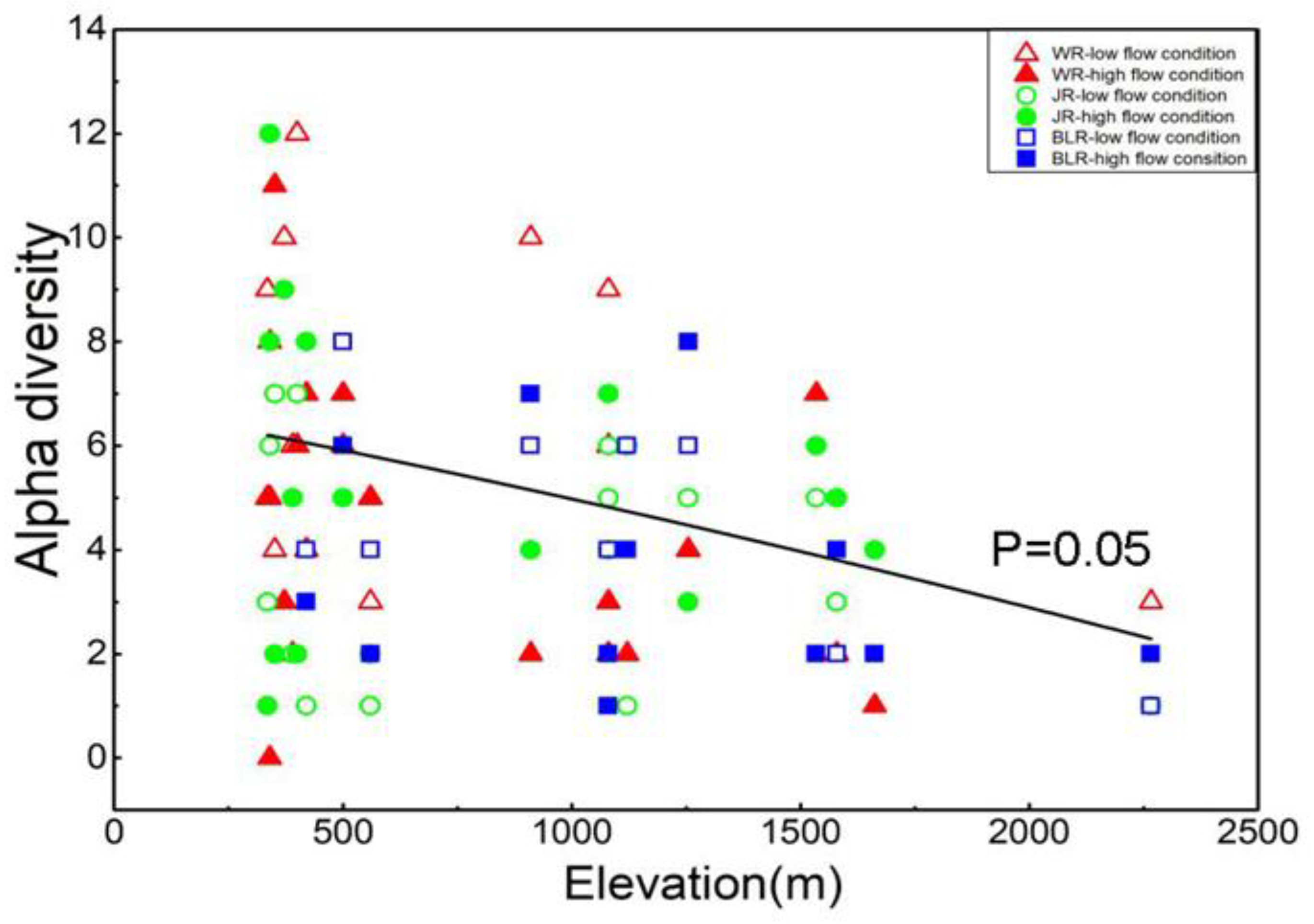
3.4. Alpha Diversity and Beta Diversity of Fish Species along Elevational Gradients
4. Discussion
4.1. Variation in Fish Species Composition and Distribution
4.2. Fish Diversity Patterns along Elevational Gradients in the Weihe River Basin
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- McNeely, J.A. Economics and Biological Diversity: Developing and Using Economic Incentives to Conserve Biological Resources; IUCN: Gland, Switzerland, 1988. [Google Scholar]
- Zainudin, M.R.Y. Assessment of fish community distribution and composition in the Perak River in order to determine biological indicators for freshwater health. Master’s Thesis, Universiti Sains Malaysia, Penang Island, Malaysia, 2005. [Google Scholar]
- Wu, W.; Xu, Z.X.; Yin, X.W.; Yu, S.Y. Fish community structure and the effect of environmental factors in the Wei River basin. Acta Sci. Circumst. 2014, 34, 1298–1308. (In Chinese) [Google Scholar]
- Lomolino, M.V. Elevation gradients of species-density: Historical and prospective views. Glob. Ecol. Biogeogr. 2001, 10, 3–13. [Google Scholar] [CrossRef]
- Bhatt, J.P.; Manish, K.; Pandit, M.K. Elevational Gradients in Fish Diversity in the Himalaya: Water Discharge Is the Key Driver of Distribution Patterns. PLoS ONE 2012, 7, e46237. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prirur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Mee, J.A.; Robins, G.L.; Post, J.R. Patterns of fish species distributions replicated across three parallel rivers suggest biotic zonation in response to a longitudinal temperature gradient. Ecol. Freshw. Fish. 2018, 27, 44–61. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.J.; Leprieur, F.; Tedesco, P.A.; Winemiller, K.O.; García-Berthou, E.; García-Berthou, E. Conservation biogeography of freshwater fishes: Recent progress and future challenges. Divers. Distrib. 2010, 16, 496–513. [Google Scholar] [CrossRef]
- Tisseuil, C.; Cornu, J.F.; Beauchard, O.; Brosse, S.; Darwall, W.; Holland, R.; Hugueny, B.; Tedesco, P.A.; Oberdorff, T. Global diversity patterns and cross-taxa convergence in freshwater systems. J. Anim. Ecol. 2013, 82, 365–376. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2004, 8, 224–239. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Leprieur, F.; Tedesco, P.A.; Hugueny, B.; Beauchard, O.; Dürr, H.H.; Brosse, S.; Oberdorff, T. Partitioning global patterns of freshwater fish beta diversity reveals contrasting signatures of past climate changes. Ecol. Lett. 2011, 14, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Sehr, M.; Keckeis, H. Habitat use of the European mudminnow Umbra krameri and association with other fish species in a disconnected Danube side arm. J. Fish Boil. 2017, 91, 1072–1093. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Bhat, A. Environmental drivers of α-diversity patterns in monsoonal tropical stream fish assemblages: A case study from tributaries of Narmada basin, India. Environ. Boil. Fishes 2017, 618, 89–761. [Google Scholar] [CrossRef]
- Benke, M.; Brändle, M.; Albrecht, C.; Wilke, T. Patterns of freshwater biodiversity in Europe: Lessons from the spring snail genus Bythinella. J. Biogeogr. 2011, 38, 2021–2032. [Google Scholar] [CrossRef]
- Hasegawa, K.; Mori, T.; Yamazaki, C. Density-dependent effects of non-native brown trout Salmo trutta on the species-area relationship in stream fish assemblages. J. Fish Biol. 2017, 90, 370–383. [Google Scholar] [CrossRef]
- Nicol, E.; Stevens, J.R.; Jobling, S. Riverine fish diversity varies according to geographical isolation and land use modification. Ecol. Evol. 2017, 7, 7872–7883. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Wu, J.; Wang, X.; Lei, G.; Chen, J. Patterns of diversity, altitudinal range and body size among freshwater fishes in the Yangtze River basin, China. Glob. Ecol. Biogeogr. 2004, 13, 543–552. [Google Scholar] [CrossRef]
- He, D.; Kang, Z.; Tao, J.; Liu, C.; Yang, J.; Chen, Y. Hydrologic connectivity driven natural stream fish assemblages in mountain streams in the Yangtze River basin: Implications for stream fish conservation in monsoonal East Asia. Hydrobiologia 2017, 785, 185–206. [Google Scholar] [CrossRef]
- Li, J.; Huang, L.; Zou, L.; Kano, Y.; Sato, T.; Yahara, T. Spatial and temporal variation of fish assemblages and their associations to habitat variables in a mountain stream of north Tiaoxi River, China. Environ. Biol. Fish. 2012, 93, 403–417. [Google Scholar] [CrossRef]
- Wu, W.; Xu, Z.; Kennard, M.J.; Yin, X.; Zuo, D. Do human disturbance variables influence more on fish community structure and function than natural variables in the Wei River basin, China? Ecol. Indic. 2016, 61, 438–446. [Google Scholar] [CrossRef]
- Bliss, S.M.; Lennox, R.J.; Midwood, J.D.; Cooke, S.J. Temporally stable and distinct fish assemblages between stream and earthen stormwater drain reaches in an urban watershed. Urban Ecosyst. 2017, 20, 1045–1055. [Google Scholar] [CrossRef]
- Peristeraki, P.; Tserpes, G.; Lampadariou, N.; Stergiou, K.I. Comparing demersal megafaunal species diversity along the depth gradient within the South Aegean and Cretan Seas (Eastern Mediterranean). PLoS ONE 2017, 12, e0184241. [Google Scholar] [CrossRef] [PubMed]
- Rolls, R.J.; Heino, J.; Ryder, D.S.; Chessman, B.C.; Growns, I.O.; Thompson, R.M.; Gido, K.B. Scaling biodiversity responses to hydrological regimes. Biol. Rev. 2018, 93, 971–995. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo-Villa, U.; Maldonado-Ocampo, J.A.; Escobar, F. Altitudinal variation in fish assemblage diversity in streams of the central Andes of Colombia. J. Fish Boil. 2010, 76, 2401–2417. [Google Scholar] [CrossRef] [PubMed]
- Cilleros, K.; Allard, L.; Vigouroux, R.; Brosse, S. Disentangling spatial and environmental determinants of fish species richness and assemblage structure in Neotropical rainforest streams. Freshw. Boil. 2017, 62, 1707–1720. [Google Scholar] [CrossRef]
- Carvajal-Quintero, J.D.; Escobar, F.; Alvarado, F.; A Villa-Navarro, F.; Jaramillo-Villa, Ú.; Maldonado-Ocampo, J.A. Variation in freshwater fish assemblages along a regional elevation gradient in the northern Andes, Colombia. Ecol. Evol. 2015, 5, 2608–2620. [Google Scholar] [CrossRef]
- Askeyev, A.; Askeyev, O.; Yanybaev, N.; Askeyev, I.; Monakhov, S.; Marić, S.; Hulsman, K. River fish assemblages along an elevation gradient in the eastern extremity of Europe. Environ. Boil. Fishes 2017, 98, 1277–1596. [Google Scholar] [CrossRef]
- Wang, J.; Soininen, J.; Zhang, Y.; Wang, B.; Yang, X.; Shen, J. Contrasting patterns in elevational diversity between microorganisms and macroorganisms. J. Biogeogr. 2011, 38, 595–603. [Google Scholar] [CrossRef]
- Harrington, R.A.; Poff, N.L.; Kondratieff, B.C. Aquatic insect β-diversity is not dependent on elevation in Southern Rocky Mountain streams. Freshwater Biol. 2016, 61, 195–205. [Google Scholar] [CrossRef]
- Li, J.; He, Q.; Hua, X.; Zhou, J.; Xu, H.; Chen, J.; Fu, C. Climate and history explain the species richness peak at mid-elevation forSchizothoraxfishes (Cypriniformes: Cyprinidae) distributed in the Tibetan Plateau and its adjacent regions. Glob. Ecol. Biogeogr. 2009, 18, 264–272. [Google Scholar] [CrossRef]
- Li, Y.; Tao, J.; Chu, L.; Yan, Y. Effects of anthropogenic disturbances on α and β diversity of fish assemblages and their longitudinal patterns in subtropical streams, China. Ecol. Freshw. Fish. 2018, 27, 433–441. [Google Scholar] [CrossRef]
- Cilleros, K.; Allard, L.; Grenouillet, G.; Brosse, S. Taxonomic and functional diversity patterns reveal different processes shaping European and Amazonian stream fish assemblages. J. Biogeogr. 2016, 43, 1832–1843. [Google Scholar] [CrossRef]
- Astorga, A.; Death, R.; Death, F.; Paavola, R.; Chakraborty, M.; Muotka, T. Habitat heterogeneity drives the geographical distribution of beta diversity: The case of New Zealand stream invertebrates. Ecol. Evol. 2014, 4, 2693–2702. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D. Connectivity and vagility determine beta diversity and nestedness in North American and European freshwater fish. J. Biogeogr. 2017, 44, 1723–1733. [Google Scholar] [CrossRef]
- Tobes, I.; Gaspar, S.; Peláez-Rodríguez, M.; Miranda, R. Spatial distribution patterns of fish assemblages relative to macroinvertebrates and environmental conditions in Andean piedmont streams of the Colombian Amazon. Inland Waters. 2016, 6, 89–104. [Google Scholar] [CrossRef]
- Liu, X.; Ao, X.; Ning, Z.; Hu, X.; Wu, X.; Ouyang, S. Diversity of fish species in suichuan river and shushui river and conservation value, China. Environ. Boil. Fishes. 2017, 100, 493–507. [Google Scholar] [CrossRef]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater Ecoregions of the World: A New Map of Biogeographic Units for Freshwater Biodiversity Conservation. Bioscience 2008, 58, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Kang, B.; Huang, X.; Wu, Y. Palaeolake isolation and biogeographical process of freshwater fishes in the Yellow River. PLoS ONE 2017, 12, e0175665. [Google Scholar] [CrossRef]
- Chang, J.; Wang, Y.; Istanbulluoglu, E.; Bai, T.; Huang, Q.; Yang, D.; Huang, S. Impact of climate change and human activities on runoff in the Weihe River Basin, China. Quatern. Int. 2015, 380–381, 169–179. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, L.; Yue, L.; Zheng, G. Petroleum hydrocarbon contamination in surface sediments of Beiluohe Basins, China. Bull Environ. Contam. Toxicol. 2008, 81, 416–421. [Google Scholar] [CrossRef]
- Song, J.; Xu, Z.; Hui, Y.; Li, H.; Li, Q. Instream flow requirements for sediment transport in the lower weihe river. Hydrol. Process. 2010, 24, 3547–3557. [Google Scholar] [CrossRef]
- Zhang, J.; Song, J.; Long, Y.; Kong, F.; Wang, L.; Zhang, Y.; Li, Q.; Wang, Y.; Hui, Y. Seasonal variability of hyporheic water exchange of the Weihe River in Shaanxi Province, China. Ecological Indic. 2018, 92, 278–287. [Google Scholar] [CrossRef]
- Ning, T.; Li, Z.; Liu, W. Vegetation dynamics and climate seasonality jointly control the interannual catchment water balance in the Loess Plateau under the Budyko framework. Hydrol Earth Syst. Sc. 2017, 21, 1515–1526. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Cheng, Q.; Zheng, B.; Li, S.; Zheng, W.; Wang, W. The Fishes of the Yellow and Bohai Sea; The Sueichan Press: Keelung, Taiwan, 1994; pp. 255–256. (In Chinese) [Google Scholar]
- Niu, S.Q.; Knouft, J.H. Hydrologic characteristics, food resource abundance, and spatial variation in stream assemblages. Ecohydrology 2017, 10, e1770. [Google Scholar] [CrossRef]
- Sommer, B.; Beger, M.; Harrison, P.L.; Babcock, R.C.; Pandolfi, J.M. Differential response to abiotic stress controls species distributions at biogeographic transition zones. Ecography 2017, 41, 478–490. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Global Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Zbinden, Z.D.; Matthews, W.J. Beta diversity of stream fish assemblages: Partitioning variation between spatial and environmental factors. Freshw. Boil. 2017, 62, 1460–1471. [Google Scholar] [CrossRef]
- Wu, W.; Xu, Z.; Yin, X.; Zuo, D. Assessment of ecosystem health based on fish assemblages in the Wei River basin, China. Environ. Monit. Assess. 2014, 186, 3701–3716. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The two-way layout with no replication. Mar. Boil. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Krumhansl, K.; Jamieson, R.; Krkosek, W. Using species traits to assess human impacts on near shore benthic ecosystems in the Canadian Arctic. Ecol. Indic. 2016, 60, 495–502. [Google Scholar] [CrossRef]
- Walker, I.R.; Levesque, A.J.; Pienitz, R.; Smol, J.P. Freshwater midges of the Yukon and adjacent Northwest Territories: A new tool for reconstructing Beringian paleoenvironments? J. N. Am. Benthol Soc. 2003, 22, 323–337. [Google Scholar] [CrossRef]
- ter Braak, C.J.F. Update Notes: CANOCO version 3.1; Agricultural Mathematics Group: Wageningen, The Netherlands, 1990. [Google Scholar]
- Zhao, C.; Yang, S.; Liu, C.; Dou, T.; Yang, Z.; Yang, Z.; Liu, X.; Xiang, H.; Nie, S.; Zhang, J.; et al. Linking hydrologic, physical and chemical habitat environments for the potential assessment of fish community rehabilitation in a developing city. J. Hydrol. 2015, 523, 384–397. [Google Scholar] [CrossRef]
- Shaanxi Institute of Zoology; Institute of Hydrobiology; Chinese Academy of Sciences; Lanzhou University Department of Biology. Qinling Fish Records; Science Press: Beijing, China, 1987. (In Chinese) [Google Scholar]
- Shaanxi Fisheries Research Institute; Department of Biology; Shaanxi Normal University. Shaanxi Fish Records; Shaanxi Science and Technology Press: Xi’an, China, 1992. (In Chinese) [Google Scholar]
- Xu, T.; Li, Z. Studies on fishes fauna of the Weihe River. J. Henan Nor. Univ. 1984, 4, 73–78. (In Chinese) [Google Scholar]
- Wang, L.; Lyons, J.; Rasmussen, P.; Seelbach, P.; Simon, T.; Wiley, M.; Kanehl, P.; Baker, E.; Niemela, S.; Stewart, P.M. Watershed, reach, and riparian influences on stream fish assemblages in the Northern Lakes and Forest Ecoregion, USA. Can. J. Fish. Aquat. Sci. 2003, 60, 491–505. [Google Scholar] [CrossRef]
- Ren, J.; Liang, G. Resource survey report of Brachymystax lenok tsinlingensis in Qianhe River Valleys of Qinling Mountains. J. Shaanxi Nor. Uni. 2004, 32, 165–168. (In Chinese) [Google Scholar]
- Zhan, C.; Qiao, C.; Xu, Z.; Yin, J. Study on ecological landscape pattern in Guanzhong part of Weihe River basin based on remote sensing. Resour Sci. 2011, 33, 2349–2355. (In Chinese) [Google Scholar]
- Zhu, D.; Chang, J. Annual variations of biotic integrity in the upper Yangtze River using an adapted index of biotic integrity (IBI). Ecol. Indic. 2008, 8, 564–572. [Google Scholar] [CrossRef]
- Da Costa, I.D.; Petry, A.C.; Mazzoni, R. Responses of fish assemblages to subtle elevations in headwater streams in southwestern Amazonia. Hydrobiologia 2017, 809, 175–184. [Google Scholar] [CrossRef]
- Cheng, S.-T.; Herricks, E.E.; Tsai, W.-P.; Chang, F.-J. Assessing the natural and anthropogenic influences on basin-wide fish species richness. Sci. Total. Environ. 2016, 572, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Kautza, A.; Sullivan, S.M.P. Relative effects of local- and landscape-scale environmental factors on stream fish assemblages: Evidence from Idaho and Ohio, USA. Fund Appl. Limnol. 2012, 180, 259–270. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Genus | Species | * Code |
|---|---|---|---|---|
| Cypriniformes | Cobitidae | Cobitis | Cobitis sinensis | S1 |
| Misgurnus | Misgurnus anguillicaudatus | S2 | ||
| Misgurnus mohoity | S3 | |||
| Paramisgurnus | Paramisgurnus dabryanus | S4 | ||
| Cyprinidae | Abbottina | Abbottina rivularis | S5 | |
| Acanthorhodeus | Acanthorhodeus macropterus | S6 | ||
| Carassius | Carassius auratus | S7 | ||
| Chanodichthys | Chanodichthys erythropterus | S8 | ||
| Ctenopharyngodon | Ctenopharyngodon idellus | S9 | ||
| Cyprinus | Cyprinus carpio | S10 | ||
| Gnathopogon | Gnathopogon imberbis | S11 | ||
| Gobio | Gobio coriparoides | S12 | ||
| Gobio rivuloides | S13 | |||
| Hemiculter | Hemiculter leucisculus | S14 | ||
| Hemiculter lucidus | S15 | |||
| Opsariichthys | Opsariichthys bidens | S16 | ||
| Pseudorasbora | Pseudorasbora parva | S17 | ||
| Rhynchocypris | Rhynchocypris lagowskii | S18 | ||
| Rhodeus | Rhodeus lighti | S19 | ||
| Rhodeus sinensis | S20 | |||
| Squaliobarbus | Squaliobarbus curriculus | S21 | ||
| Nemacheilidae | Triplophysa | Trilophysa bleekeri | S22 | |
| Triplophysa brachyptera | S23 | |||
| Triplophysa dalaica | S24 | |||
| Triplophysa kungessana | S25 | |||
| Triplophysa pappenheimi | S26 | |||
| Triplophysa robusta | S27 | |||
| Triplophysa sellaefer | S28 | |||
| Triplophysa stoliczkai | S29 | |||
| Perciformes | Channidae | Channa | Channa argus | S30 |
| Cichlidae | Oreochromis | Oreochromis mossambicus | S31 | |
| Gobiidae | Favonigobius | Favonigobius gymnauchen | S32 | |
| Rhinogobius | Rhinogobius brunneus | S33 | ||
| Rhinogobius cliffordpopei | S34 | |||
| Rhinogobius giurinus | S35 | |||
| Odontobutidae | Micropercops | Micropercops swinhonis | S36 | |
| Salmoniformes | Salmonidae | Brachymystax | Brachymystax lenok | S37 |
| Siluriformes | Bagridae | Tachysurus | Tachysurus nitidus | S38 |
| Siluridae | Silurus | Silurus asotus | S39 |
| High-Flow Condition | Low-Flow Condition | ||||||
|---|---|---|---|---|---|---|---|
| R | WR | JR | BLR | WR | JR | BLR | |
| WR | Global R | -- | 0.153 | 0.183 | -- | 0.101 | 0.111 |
| p | -- | 0.004 | 0.007 | -- | 0.020 | 0.046 | |
| JR | Global R | 0.153 | -- | 0.101 | 0.101 | -- | 0.002 |
| p | 0.004 | -- | 0.05 | 0.020 | -- | 0.460 | |
| BLR | Global R | 0.183 | 0.101 | -- | 0.111 | 0.002 | -- |
| p | 0.007 | 0.05 | -- | 0.046 | 0.460 | -- | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, D.; Zhao, X.; Song, J.; Sun, H.; Wang, S.; Bai, H.; Li, Q. Quantifying the Distribution and Diversity of Fish Species Along Elevational Gradients in the Weihe River Basin, Northwest China. Sustainability 2019, 11, 6177. https://doi.org/10.3390/su11216177
Cheng D, Zhao X, Song J, Sun H, Wang S, Bai H, Li Q. Quantifying the Distribution and Diversity of Fish Species Along Elevational Gradients in the Weihe River Basin, Northwest China. Sustainability. 2019; 11(21):6177. https://doi.org/10.3390/su11216177
Chicago/Turabian StyleCheng, Dandong, Xiaotian Zhao, Jinxi Song, Haotian Sun, Shaoqing Wang, Haifeng Bai, and Qi Li. 2019. "Quantifying the Distribution and Diversity of Fish Species Along Elevational Gradients in the Weihe River Basin, Northwest China" Sustainability 11, no. 21: 6177. https://doi.org/10.3390/su11216177
APA StyleCheng, D., Zhao, X., Song, J., Sun, H., Wang, S., Bai, H., & Li, Q. (2019). Quantifying the Distribution and Diversity of Fish Species Along Elevational Gradients in the Weihe River Basin, Northwest China. Sustainability, 11(21), 6177. https://doi.org/10.3390/su11216177