Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern Iberian Peninsula


 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
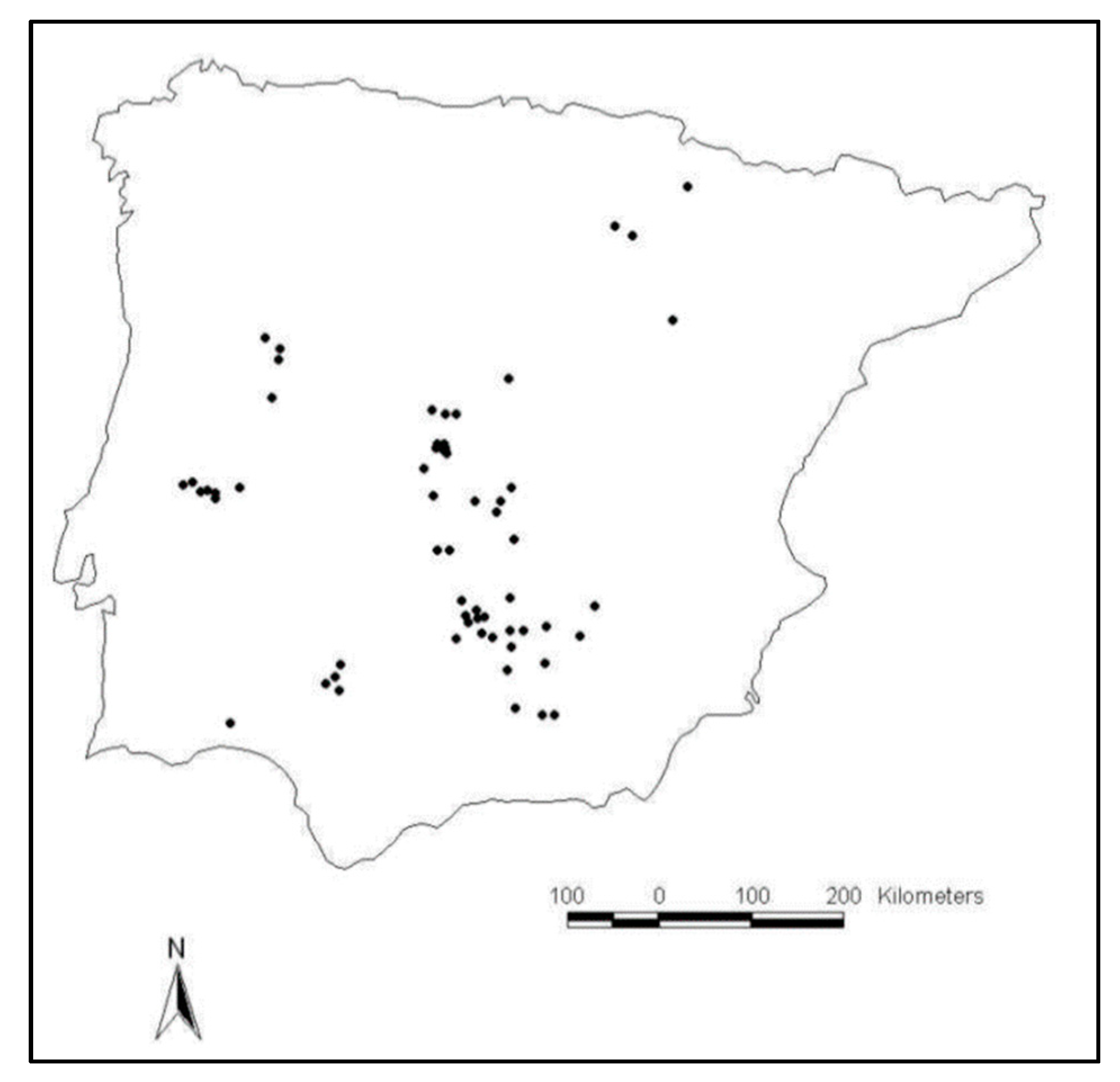
2.1. Study Area
Location, Climate, Geomorphology and Soils
2.2. Methods
3. Results
3.1. Phytosociological Classification Based on Numerical Analyses
3.2. Synthetic Vegetation Analysis
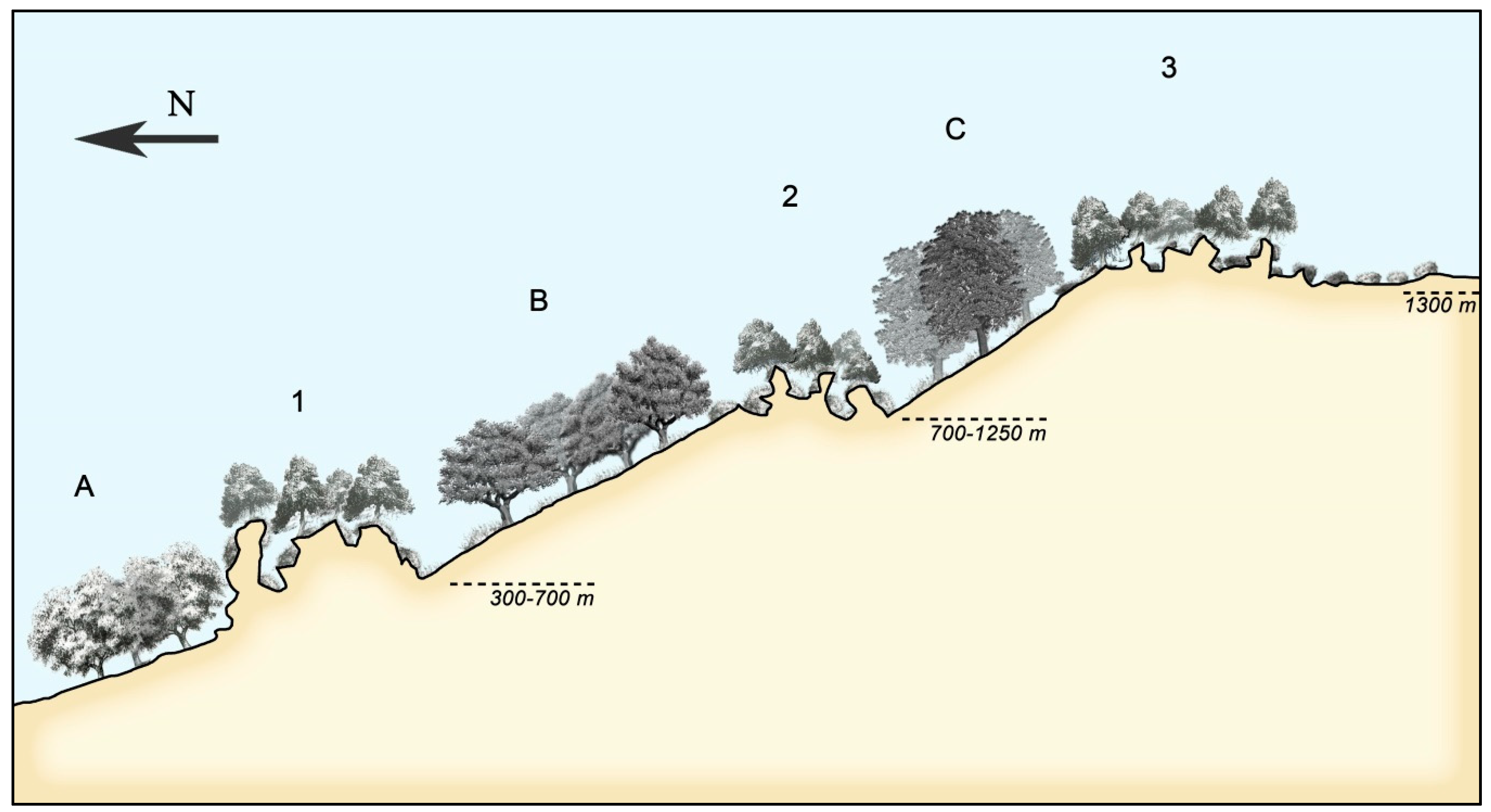
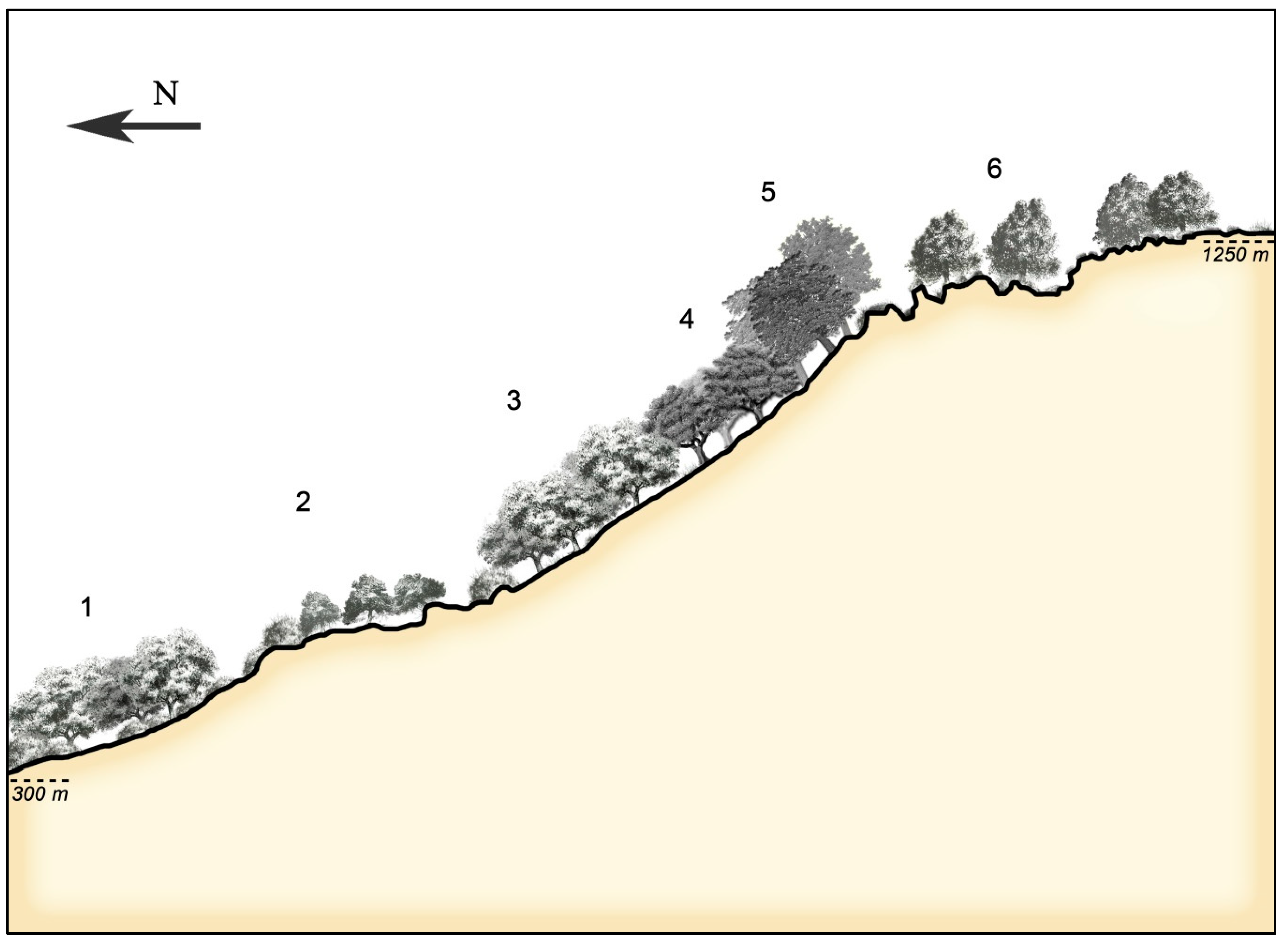
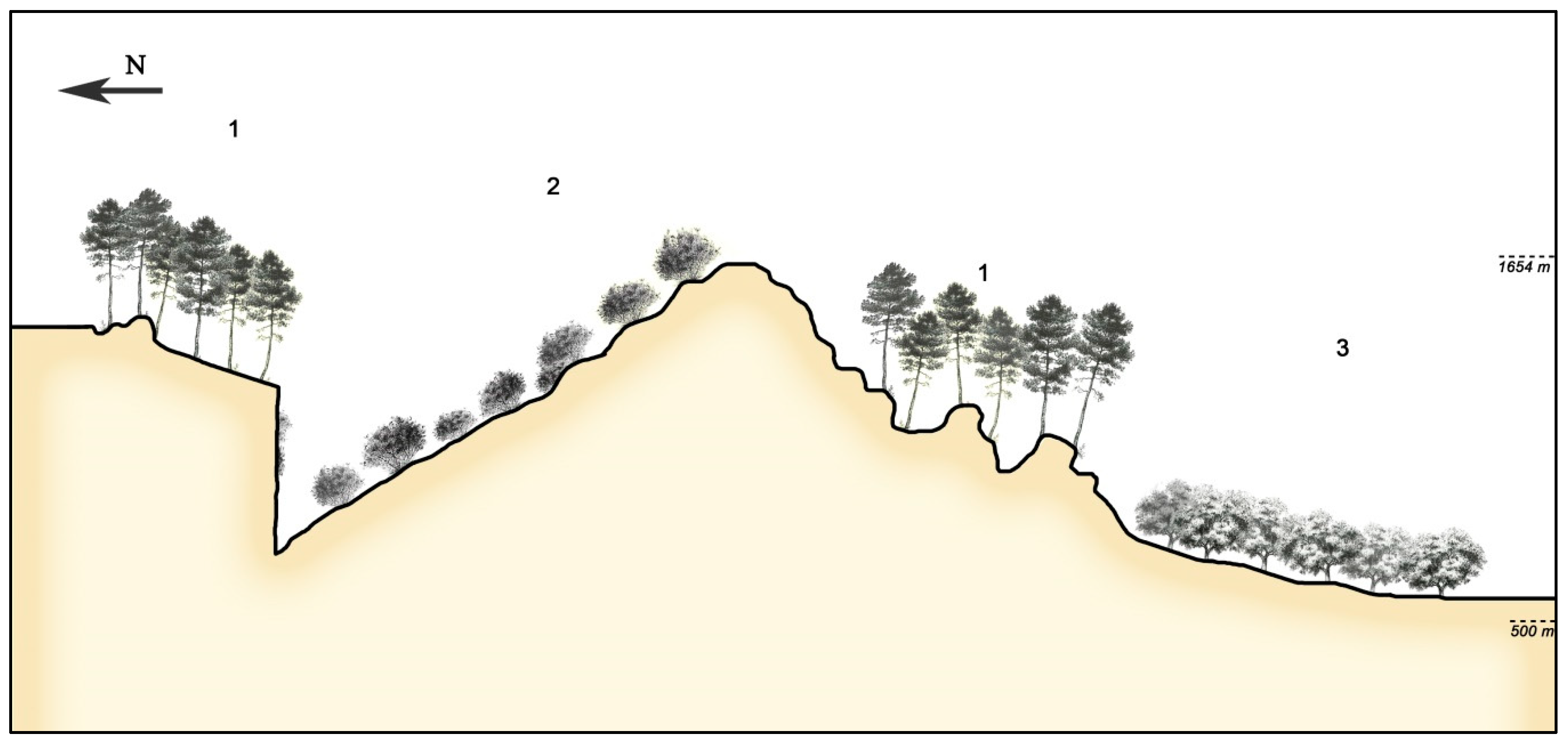
3.3. Catenal Analysis of the Landscape Evolution
4. Discussion
5. Conclusions
| QUERCETEA ILICIS Br.-Bl. ex A. O. Bolòs 1950 | |||
| Pistacio lentisci-Rhamnetalia alaterni Rivas-Martínez 1975 | |||
| Juniperion badiae Cano, Rodríguez Torres, Pinto Gomes, García Fuentes, Torres, Salazar, Ruiz, Cano-Ortiz & Montilla 2007 ex Mucina et al. 2016 nom. corr. hoc loco | |||
| Festuco merinoi-Juniperetum badiae (Rivas-Martínez & Sánchez Mata 1989) Sánchez Mata 1999 corr. Rivas-Martínez & Sánchez Mata 2011 nom. corr. hoc loco | |||
| Cytiso tribracteolati-Juniperetum oxycedri Pérez Latorre, Galán & Cabezudo in Pérez Latorre, Galán, Navas P., Gil & Cabezudo 1999 | |||
| Echinosparto iberici-Juniperetum badiae Rodríguez Torres & Cano in Cano, Rodríguez Torres, Pinto Gomes, García Fuentes, Torres, Salazar, Ruiz, Cano-Ortiz & Montilla 2007 nom. corr. hoc loco | |||
| Cytiso eriocarpi-Juniperetum badiae Pinto & Cano in Cano, Rodríguez Torres, Pinto Gomes, García Fuentes, Torres, Salazar, Ruiz, Cano-Ortiz & Montilla 2007 nom. corr. hoc loco | |||
| Pistacio terebinthi-Juniperetum badiae Cano, Rodríguez Torres, Pinto Gomes, García, Torres, Salazar, Ruiz, Cano-Ortiz & Montilla 2007 nom. corr. hoc loco | |||
| Stipo tenacissimae-Juniperetum badiae Cano, Rodríguez Torres, Pinto Gomes, García Fuentes, Torres, Salazar, Ruiz, Cano-Ortiz & Montilla 2007 nom. corr. hoc loco | |||
| Juniperetum phoeniceae-badiae ass. nova hoc loco | |||
| Teline patentis-Pistacietum terebinthi ass. nova hoc loco | |||
| Myrto communis-Juniperetum badiae ass. nova hoc loco | |||
| Genisto polyanthi-Juniperetum badiae ass. nova hoc loco | |||
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | JPB | TP | MJ | EJ | FJ | CJ | PJ | SJ | GJ |
|---|---|---|---|---|---|---|---|---|---|
| Juniperus oxycedrus subsp. badia | V | V | V | V | V | V | V | V | V |
| Quercus rotundifolia | IV | IV | III | III | V | - | V | V | V |
| Asparagus acutifolius | I | I | I | - | - | - | II | II | I |
| Pistacia lentiscus | I | III | II | - | - | III | - | - | I |
| Juniperus oxycedrus subsp. oxycedrus | IV | - | I | - | - | V | I | - | I |
| Quercus coccifera | I | III | I | - | - | - | I | III | - |
| Daphne gnidium | III | - | I | - | - | II | V | III | - |
| Thapsia villosa | I | I | I | - | - | - | III | I | - |
| Pistacia terebinthus | I | V | II | - | - | I | IV | - | - |
| Jasminum fruticans | I | II | I | - | - | - | III | - | - |
| Quercus faginea | I | I | - | - | - | - | I | - | - |
| Rubia peregrina | I | I | I | - | - | - | I | - | - |
| Phillyrea latifolia | I | I | I | - | - | I | - | - | - |
| Rhamnus alaternus | I | I | I | - | - | I | - | - | - |
| Smilax aspera | I | I | I | - | - | - | - | - | - |
| Carex hallerana | I | - | - | - | - | - | - | I | - |
| Juniperus phoenicea | V | I | - | - | - | - | - | - | - |
| Crataegus laciniata | I | I | - | - | - | - | - | - | - |
| Paeonia broteroi | I | - | - | - | - | - | - | - | - |
| Pinus halepensis | I | - | - | - | - | - | - | - | - |
| Teline patens | - | III | - | - | - | - | - | - | - |
| Rhamnus myrtifolius | - | I | - | - | - | - | - | - | - |
| Coronilla glauca | - | I | - | - | - | - | - | - | - |
| Phillyrea angustifolia | - | I | III | I | - | V | - | - | I |
| Olea europea var. sylvestris | - | I | II | - | - | II | II | I | - |
| Viburnum tinus | - | I | I | - | - | I | - | - | - |
| Arbutus unedo | - | II | II | I | - | - | - | - | - |
| Ruscus aculeatus | - | I | - | - | - | I | - | - | - |
| Pinus pinaster | - | I | - | I | - | - | - | - | - |
| Erica arborea | - | - | I | I | IV | V | - | - | I |
| Rhamnus lycioides | - | - | I | - | - | - | II | I | - |
| Osyris alba | - | - | I | - | - | - | III | I | - |
| Quercus suber | - | - | I | I | - | - | I | - | - |
| Quercus broteroi | - | - | I | - | - | - | I | - | - |
| Pyrus bourgaeana | - | - | I | - | - | - | I | - | - |
| Myrtus communis | - | - | II | - | - | III | - | - | - |
| Asparagus aphyllus | - | - | I | - | - | I | - | - | - |
| Rhamnus oleoides | - | - | I | - | - | II | - | - | - |
| Pistacia x saportae | - | - | I | - | - | I | - | - | - |
| Asparagus albus | - | - | I | - | - | - | - | - | - |
| Teucrium fruticans | - | - | I | - | - | - | - | - | - |
| Quercus marianica | - | - | I | - | - | - | - | - | - |
| Quercus canariensis | - | - | I | - | - | - | - | - | - |
| Phlomis purpurea | - | - | I | - | - | - | - | - | - |
| Crataegus monogyna | - | - | I | - | - | - | - | - | - |
| Genista polyanthos | - | - | - | - | - | - | - | - | V |
| Companions | |||||||||
| Urginea maritima | I | I | II | I | - | - | I | I | I |
| Thymus mastichina | II | I | I | - | - | - | V | II | I |
| Rosmarinus officinalis | III | III | III | - | - | - | I | - | II |
| Asphodelus albus | III | I | I | II | - | - | - | - | II |
| Cistus albidus | III | III | II | - | - | - | - | III | I |
| Dactylis hispanica | I | I | I | - | IV | - | I | IV | - |
| Retama sphaerocarpa | I | - | I | - | - | - | V | II | I |
| Asplenium ceterach | I | I | I | - | - | - | I | I | - |
| Rubus ulmifolius | I | II | III | - | - | - | I | - | - |
| Phagnalon saxatile | I | - | - | - | - | - | I | III | - |
| Geranium purpureum | I | I | - | - | - | - | - | - | I |
| Phlomis lychnitis | II | I | - | - | - | - | - | III | - |
| Lonicera periclymenum subsp. hispanica | I | I | - | - | - | - | I | - | - |
| Staehelina dubia | I | I | - | - | - | - | - | I | - |
| Santolina canescens | I | - | - | - | - | - | I | - | - |
| Halimium atriplicifolium | I | I | I | - | - | - | - | - | - |
| Melica minuta | I | I | I | - | - | - | - | - | - |
| Sanguisorba minor | I | - | I | - | - | - | - | - | - |
| Thymus zygis subsp. gracilis | I | I | - | - | - | - | - | - | - |
| Thymus orospedanus | III | II | - | - | - | - | - | - | - |
| Aphyllantes monspeliensis | I | II | - | - | - | - | - | - | - |
| Rosa canina | I | I | - | - | - | - | - | - | - |
| Brachypodium retusum | II | I | - | - | - | - | - | - | - |
| Hedera ibernica | I | I | - | - | - | - | - | - | - |
| Helleborus foetidus | I | I | - | - | - | - | - | - | - |
| Lavandula latifolia | I | - | - | - | - | - | - | - | - |
| Cistus monspeliensis | I | - | - | - | - | - | - | - | - |
| Teucrium capitatum | I | - | - | - | - | - | - | - | - |
| Teucrium rotundifolium | I | - | - | - | - | - | - | - | - |
| Helianthemum ledifolium | I | - | - | - | - | - | - | - | - |
| Clematis flammula | I | - | - | - | - | - | - | - | - |
| Linum suffruticosum | I | - | - | - | - | - | - | - | - |
| Bupleurum rigidum | I | - | - | - | - | - | - | - | - |
| Ilex aquifolium | I | - | - | - | - | - | - | - | - |
| Fumana thymifolia | I | - | - | - | - | - | - | - | - |
| Lonicera splendida | I | - | - | - | - | - | - | - | - |
| Leuzea conifera | I | - | - | - | - | - | - | - | - |
| Berberis hispanica | I | - | - | - | - | - | - | - | - |
| Buxus sempervirens | I | - | - | - | - | - | - | - | - |
| Pinus salzmannii | I | - | - | - | - | - | - | - | - |
| Helianthemum syriacum | I | - | - | - | - | - | - | - | - |
| Filipendula vulgaris | I | - | - | - | - | - | - | - | - |
| Polygala monspeliaca | I | - | - | - | - | - | - | - | - |
| Ophrys tenthrediniferfa | I | - | - | - | - | - | - | - | - |
| Helianthemum cinereum subsp. rotundifolium | I | - | - | - | - | - | - | - | - |
| Ptilostemon hispanicus | I | - | - | - | - | - | - | - | - |
| Anthyllis vulneraria subsp. maura | I | - | - | - | - | - | - | - | - |
| Arrhenatherum album | I | - | - | - | - | - | - | - | - |
| Erinacea anthyllis | I | - | - | - | - | - | - | - | - |
| Fumana paradoxa | I | - | - | - | - | - | - | - | - |
| Helictotrichon filifolium | I | - | - | - | - | - | - | - | - |
| Crupina crupinastrum | I | - | - | - | - | - | - | - | - |
| Narcissus assoanus | I | - | - | - | - | - | - | - | - |
| Medicago rigidula | I | - | - | - | - | - | - | - | - |
| Genista scorpius | I | - | - | - | - | - | - | - | - |
| Crocus serotinus subsp. salzmannii | I | - | - | - | - | - | - | - | - |
| Prunus spinosa | I | - | - | - | - | - | - | - | - |
| Cytisus scoparius subsp. reverchonii | I | - | - | - | - | - | - | - | - |
| Fumana laevipes | I | - | - | - | - | - | - | - | - |
| Genista boissieri | I | - | - | - | - | - | - | - | - |
| Amelanchier ovalis | I | - | - | - | - | - | - | - | - |
| Muscari giennense | I | - | - | - | - | - | - | - | - |
| Thymus granatensis | I | - | - | - | - | - | - | - | - |
| Biscutella sempervirens | I | - | - | - | - | - | - | - | - |
| Sedum brevifolium | - | I | I | I | - | IV | - | I | II |
| Mucizonia hispida | - | I | I | I | - | - | - | - | II |
| Tamus communis | - | II | I | - | - | - | II | - | - |
| Fraxinus angustifolia | - | I | I | - | - | - | - | - | I |
| Polygala rupestris | - | I | I | - | - | - | - | - | - |
| Sedum sediforme | - | I | I | - | - | - | - | - | - |
| Vitis vinifera subsp. sylvestris | - | I | I | - | - | - | - | - | - |
| Lithodora fruticosa | - | I | - | - | - | - | - | - | - |
| Biscutella valentina | - | I | - | - | - | - | - | - | - |
| Rubus caesius | - | I | - | - | - | - | - | - | - |
| Clematis vitalba | - | I | - | - | - | - | - | - | - |
| Euphorbia characias | - | I | - | - | - | - | - | - | - |
| Genista cinerea subsp. speciosa | - | I | - | - | - | - | - | - | - |
| Dianthus lusitanus | - | - | II | III | IV | IV | I | II | IV |
| Cistus ladanifer | - | - | III | III | - | III | I | I | III |
| Halimium umbellatum subsp. viscosum | - | I | I | - | III | I | I | I | |
| Arrhenatherum bulbosum | - | - | I | I | - | II | II | - | III |
| Halimium ocymoides | - | - | I | II | - | III | - | - | II |
| Lavandula sampaiana | - | - | II | II | - | III | IV | III | - |
| Sedum dasyphyllum | - | - | I | I | - | - | - | - | I |
| Coincya longirrostra | - | - | I | I | - | - | - | - | I |
| Digitalis mariana | - | - | I | II | - | - | - | - | I |
| Jasione mariana | - | - | I | II | - | - | - | - | III |
| Linaria saxatilis | - | - | I | II | - | - | - | - | II |
| Lavandula luisieri | - | - | I | I | - | - | - | - | I |
| Elymus caninus | - | - | I | I | - | - | - | - | I |
| Nerium oleander | - | - | I | - | - | - | - | - | I |
| Anogramma leptophylla | - | - | I | - | - | - | - | - | I |
| Adenocarpus telonensis | - | - | II | - | - | - | I | I | - |
| Cytisuss striatus subsp. eriocarpus | - | - | I | - | V | IV | I | - | - |
| Cistus salvifolius | - | - | I | - | - | III | - | III | - |
| Bryonia cretica | - | - | I | - | - | - | I | - | - |
| Flueggea tinctoria | - | - | I | - | - | - | - | - | I |
| Genista hirsuta | - | - | I | - | - | - | I | - | - |
| Teucrium gnaphalodes | - | - | I | - | - | - | - | I | - |
| Teucrium pseudochamaepytis | - | - | I | - | - | - | - | III | - |
| Cheilanthes tinaei | - | - | I | - | - | - | - | I | - |
| Calluna vulgaris | - | - | I | I | - | II | - | - | - |
| Cheilanthes hispanica | - | - | I | - | - | II | - | - | - |
| Adenocarpus argyrophyllus | - | - | I | II | - | - | - | - | - |
| Ericca scoparia | - | - | I | I | - | - | - | - | - |
| Astragalus lusitanicus | - | - | I | I | - | - | - | - | - |
| Jasione tomentosa | - | - | I | I | - | - | - | - | - |
| Cheilanthes maderensis | - | - | II | - | - | - | - | - | - |
| Arisarum simorhinum | - | - | I | - | - | - | - | - | - |
| Sedum album | - | - | I | - | - | - | - | - | - |
| Stipa capensis | - | - | I | - | - | - | - | - | - |
| Hyacinthoides hispanica | - | - | I | - | - | - | - | - | - |
| Pterocepalus diandrus | - | - | I | - | - | - | - | - | - |
| Digitalis heywoodii var. albicans | - | - | I | - | - | - | - | - | - |
| Antirrhinum graniticum subsp. onubensis | - | I | - | - | - | - | - | - | |
| Cytisus scoparius subsp. bourgaei | - | - | I | - | - | - | - | - | - |
| Aristolochia baetica | - | - | I | - | - | - | - | - | - |
| Micromeria graeca | - | - | I | - | - | - | - | - | - |
| Dianthus crassipes | - | - | I | - | - | - | - | - | - |
| Jasonia glutinosa | - | - | I | - | - | - | - | - | - |
| Helianthemum croceum | - | - | I | - | - | - | - | - | - |
| Stipa gigantea | - | - | - | II | - | - | I | II | II |
| Conopodium capillifolium | - | - | - | I | - | I | - | - | I |
| Festuca elegans | - | - | - | II | V | - | - | - | I |
| Armeria capitella | - | - | - | I | - | - | - | - | I |
| Narcissus rupicola | - | - | - | I | - | - | - | - | I |
| Digitalis thapsi | - | - | - | I | - | II | II | - | - |
| Lavandula pedunculata | - | - | - | - | IV | - | I | - | - |
| Erica australis | - | - | - | I | - | II | - | - | - |
| Gladiolus illyricus | - | - | - | I | - | I | - | - | - |
| Echinospartum ibericum | - | - | - | II | - | - | - | - | - |
| Quercus pyrenaica | - | - | - | I | - | - | - | - | - |
| Arenaria querioides | - | - | - | I | - | - | - | - | - |
| Armeria arenaria subsp. segoviensis | - | - | - | I | - | - | - | - | - |
| Erica scoparia | - | - | - | I | - | - | - | - | - |
| Leucanthemopsis flaveola | - | - | - | I | - | - | - | - | - |
| Festuca summilusitanica | - | - | - | - | V | - | - | - | - |
| Cytisus oromediterraneus | - | - | - | - | IV | - | - | - | - |
| Juniperus hemisphaerica | - | - | - | - | IV | - | - | - | - |
| Arenaria grandiflora | - | - | - | - | V | - | - | - | - |
| Thymus x bractichina | - | - | - | - | V | - | - | - | - |
| Pteridium aquilinum | - | - | - | - | V | - | - | - | - |
| Sorbus aucuparia | - | - | - | - | II | - | - | - | - |
| Jasione sessiliflora | - | - | - | - | IV | - | - | - | - |
| Genista cinerascens | - | - | - | - | I | - | - | - | - |
| Sedum hirsutum | - | - | - | - | IV | IV | - | - | - |
| Simethis planifolia | - | - | - | - | - | II | - | - | - |
| Polypodium cambricum | - | - | - | - | - | I | - | - | - |
| Adenocarpus anisochilus | - | - | - | - | - | I | - | - | - |
| Hypericum linearifolium | - | - | - | - | - | I | - | - | - |
| Thymus zygis | - | - | - | - | - | - | I | III | - |
| Helichrysum stoechas | - | - | - | - | - | - | I | IV | - |
| Cynosurus echinatus | - | - | - | - | - | - | I | I | - |
| Santolina rosmarinifolia | - | - | - | - | - | - | I | I | - |
| Hyparrhenia hirta | - | - | - | - | - | - | I | I | - |
| Cytisus scoparius subsp. scoparius | - | - | - | - | - | - | III | - | - |
| Helianthemum appeninum | - | - | - | - | - | - | I | - | - |
| Hedera helix | - | - | - | - | - | - | I | - | - |
| Lonicera implexa | - | - | - | - | - | - | I | - | - |
| Thapsia maxima | - | - | - | - | - | - | I | - | - |
| Antirrhinum graniticum | - | - | - | - | - | - | I | - | - |
| Stipa tenacissima | - | - | - | - | - | - | - | III | - |
| Cytisus multiflorus | - | - | - | - | - | - | - | I | - |
| Cistus populifolius x C. salvifolius | - | - | - | - | - | - | - | - | I |
| Festuca rothmaleri | - | - | - | - | - | - | - | - | I |
| Conopodium bourgaei | - | - | - | - | - | - | - | - | I |
| Asplenium billotii | - | - | - | - | - | - | - | - | I |
| Scrophularia canina | - | - | - | - | - | - | - | - | I |
| Genista florida | - | - | - | - | - | - | - | - | I |
References
- Adams, R.P. Taxonomy of Juniperus, section Juniperus: Sequence analysis of nrDNA and five cpDNA regions. Phytologia 2012, 94, 280–297. [Google Scholar]
- Adams, R.P. Junipers of the World: The Genus Juniperus, 4th ed.; Trafford Publishing: Vancouver, Canada, 2014. [Google Scholar]
- Adams, R.P. Morphological comparison and key to Juniperus deltoides and J. oxycedrus. Phytologia 2014, 96, 58–62. [Google Scholar]
- Amaral Franco, J. Juniperus. In Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 1); Castroviejo, S., Laínz, M., López González, G., Monserrat, P., Muñoz Garmendia, F., Paiva, J., Villar, L., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1986; pp. 181–188. [Google Scholar]
- Pignatti, S. Juniperus; Flora d’Italia 1; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Brus, R.; Idzjtic´, M.; Jarni, K. Morphologic variation in northern marginal Juniperus oxycedrus L. subsp. oxycedus population in Istria. Plant Biosyst. 2016, 150, 274–284. [Google Scholar] [CrossRef]
- Adams, R.P. Juniperus deltoides, a new species, and nomenclatural notes on Juniperus polycarpos and Juniperus turcomanica (Cupressaceae). Phytologia 2004, 86, 49–53. [Google Scholar]
- Adams, R.P.; Tashev, A.N. Geographical variation in leaf oils of Juniperus deltoides from Bulgaria, Greece, Italy and Turkey. Phytologia 2012, 94, 310–318. [Google Scholar]
- Adams, R.P.; Farzaliyev, V.; Gucel, S.; Leschner, H.V.; Mataraci, T.; Tashev, A.N.; Schwarzbach, A.E. nrDNA and petN-psbM sequencing reveals putative Juniperus oxycedrus L. from Azerbaijan, Bulgaria, Cyprus and Israel to be J. deltoides R.P. Adams. Phytologia 2015, 97, 286–290. [Google Scholar]
- Adams, R.P.; Joaquin Altarejos, J.; Fernandez, C.; Camacho, A. The leaf essential oils and taxonomy of Juniperus oxycedrus L. subsp. oxycedrus, subsp. badia (H. Gay) Debeaux, and subsp. macrocarpa (Sibth. & Sm.). Ball. J. Essent. Oil Res. 1999, 11, 167–172. [Google Scholar]
- Salido, S.; Altarejos, J.; Nogueras, M.; Sánchez, A.; Pannecouque, C.; Witvrouw, M.; De Clercq, E. Chemical studies of essential oils of Juniperus oxycedrus ssp. badia. J. Ethnopharmacol. 2002, 81, 129–134. [Google Scholar] [CrossRef]
- Adams, R.P. Systematics of Juniperus section Juniperus based on leaf essential oils and random amplified polymorphic DNAs (RAPDs). Biochem. Syst. Ecol. 2000, 28, 515–528. [Google Scholar] [CrossRef]
- Adams, R.P.; Morris, J.A.; Pandey, R.N.; Schwarzbach, A.E. Cryptic speciation between Juniperus deltoides and Juniperus oxycedrus (Cupressaceae) in the Mediterranean. Biochem. Syst. Ecol. 2005, 33, 771–787. [Google Scholar] [CrossRef]
- Roma-Marzio, F.; Najar, B.; Alessandri, J.; Pistelli, L.; Peruzzi, L. Taxonomy of prickly juniper (Juniperus oxycedrus group): A phytochemical-morphometric combined approach at the contact zone of two cryptospecies. Phytochemistry 2017, 141, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Martínez, S. Memoria del mapa de Series de Vegetación de España; ICONA. Serie Técnica. Publ. Ministerio Agricultura, Pesca y Alimentación: Madrid, Spain, 1987; pp. 1–268. [Google Scholar]
- Bolòs, O.; Vigo, J. Flora dels Països Catalans; Barcino: Barcelona, Spain, 1984. [Google Scholar]
- Rivas-Martínez, S.; Díaz, T.E.; Fernández González, F.; Izco, J.; Loidi, J.; Lousa, M.; Penas, A. Vascular plant communities of Spain and Portugal. Itinera Geobot. 2002, 15, 433–922. [Google Scholar]
- Vicioso, C. Notas sobre la flora española. An. Jardin Bot. Madrid 1946, 6, 5–89. [Google Scholar]
- Cano, E.; Rodríguez Torres, A.; Pinto Gomes, C.J.; García Fuentes, A.; Torres, J.A.; Salazar, C.; Ruiz, L.; Cano-Ortiz, A.; Montilla, R. Analysis of the Juniperus oxycedrus L. in the centre and south of the Iberian Peninsula (Spain and Portugal). Acta Bot. Gallica 2007, 154, 79–99. [Google Scholar] [CrossRef]
- Quinto-Canas, R.; Mendes, P.; Cano-Ortiz, A.; Musarella, C.M.; Pinto-Gomes, C. Forest fringe communities of the southwestern Iberian Peninsula. Rev. Chapingo Ser. Cie. 2018, 24, 415–434. [Google Scholar] [CrossRef]
- Melendo, M.; Giménez, E.; Cano, E.; Gómez Mercado, F.; Valle, F. The endemic flora in the south of the Iberian Peninsula: taxonomic composition, biological spectrum, pollination, reproductive mode and dispersal. Flora 2003, 198, 260–276. [Google Scholar] [CrossRef]
- Cano, E.; Melendo, M.; Valle, F. The plant communities of the Asplenietea trichomanis in the SW Iberian Península. Folia Geobot. 1997, 32, 361–376. [Google Scholar]
- Cano-Ortiz, A.; Pinto Gomes, C.J.; Musarella, C.M.; Cano, E. Expansion of the Juniperus genus due to anthropic activity. In Old-Growth Forest and Coniferous Forests; Weber, R.P., Ed.; Nova Science Publishers: New York, NY, USA, 2015; pp. 55–65. [Google Scholar]
- Mendes, P.; Meireles, C.; Vila-Viçosa, C.; Musarella, C.M.; Pinto-Gomes, C. Best management practices to face degraded territories occupied by Cistus ladanifer shrublands – Portugal case study. Plant Biosyst. 2015, 149, 494–502. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Loidi, J. Bioclimatology of the Iberian Peninsula. Itinera Geobot. 1999, 13, 41–47. [Google Scholar]
- Cano, E.; Torres, J.A.; Cano-Ortiz, A.; Montilla, R.J. Una nueva asociación de matorral para la alianza Lavandulo-Genistion Boissieri en el sector subbético. Lagascalia 2005, 25, 125–133. [Google Scholar]
- Castroviejo, S.; Laínz, M.; López González, G.; Monserrat, P.; Muñoz Garmendia, F.; Paiva, J.; Villar, L. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 1–2); Real Jardín Botánico, CSIC: Madrid, Spain, 1990. [Google Scholar]
- Castroviejo, S.; Aedo, C.; Cirujano, S.; Laínz, M.; Monserrat, P.; Morales, R.; Muñoz Garmendia, F.; Navarro, C.; Paiva, J.; Soriano, C. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 3); Real Jardín Botánico, CSIC: Madrid, Spain, 1993. [Google Scholar]
- Castroviejo, S.; Aedo, C.; Gómez Campo, C.; Laínz, M.; Monserrat, P.; Morales, R.; Muñoz Garmendia, F.; Nieto Feliner, G.; Rico, E.; Talavera, S.; Villar, L. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 4); Real Jardín Botánico, CSIC: Madrid, Spain, 1993. [Google Scholar]
- Castroviejo, S.; Aedo, C.; Laínz, M.; Morales, R.; Muñoz Garmendia, F.; Nieto Feliner, G.; Paiva, J. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 5); Real Jardín Botánico, CSIC: Madrid, Spain, 1997. [Google Scholar]
- Castroviejo, S.; Aedo, C.; Benedí, C.; Laínz, M.; Muñoz Garmendia, F.; Nieto Feliner, G.; Paiva, J. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 8); Real Jardín Botánico, CSIC: Madrid, Spain, 1997. [Google Scholar]
- Muñoz Garmendia, F.; Navarro, C. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 6); Real Jardín Botánico, CSIC: Madrid, Spain, 1998. [Google Scholar]
- Paiva, J.; Sales, F.; Hedge, I.C.; Aedo, C.; Aldasoro, J.J.; Castroviejo, S.; Herrero, A.; Velayos, M. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 14); Real Jardín Botánico, CSIC: Madrid, Spain, 2001. [Google Scholar]
- Talavera, S.; Aedo, C.; Castroviejo, S.; Romero Zarco, C.; Saéz, C.; Salgueiro, F.J.; Velayos, M. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol 7(1)); Real Jardín Botánico, CSIC: Madrid, Spain, 1999. [Google Scholar]
- Talavera, S.; Aedo, C.; Castroviejo, C.; Herrero, A.; Romero Zarco, C.; Salgueiro, F.J.; Velayos, S. (Eds.) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Vol. 7(2)); Real Jardín Botánico, CSIC: Madrid, Spain, 2000. [Google Scholar]
- Tutin, T.; Heywood, V.H.; Burges, D.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea (Vol. I to V); Cambrige at the University Press: Cambrige, UK, 1964. [Google Scholar]
- Valdés, B.; Talavera, S.; Fernández Galiano, E. (Eds.) Flora Vascular de Andalucía Occidental; Ketres Editora S A: Barcelona, Spain, 1987. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Tichý, L. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Sánchez Mata, D. Bioclimatología: una Ciencia Avanzada para la Caracterización del Medio Natural; Discursos de Entrada 1998; Publicaciones Institución Gran Duque de Alba: Ávila, Spain, 1999. [Google Scholar]
- Braun-Blanquet, J. Fitosociología. Bases para el Estudio de las Comunidades Vegetales; Blume: Madrid, Spain, 1979. [Google Scholar]
- Géhu, J.M.; Rivas-Martínez, S. Notions fondamentales de Phytosociologie. In Berichte der Internationalen Symposien der Internationalen Vereinigung für Vegetationskunde; Dierschk, H., Ed.; Cramer: Vaduz, Liechtenstein, 1981; pp. 5–33. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 2001, 4, 1–9. [Google Scholar]
- Montero Burgos, J.L.; González Rebollar, J.L. Diagramas Bioclimáticos; Ministerio de Agricultura, Pesca y Alimentación. ICONA: Madrid, Spain, 1983. [Google Scholar]
- Pérez Latorre, A.V.; Galán de Mera, A.; Navas, P.; Navas, D.; Gil, Y. Datos sobre la flora y vegetación del Parque Natural de los Alcornocales (Cádiz-Málaga, España). Acta Bot. Malacit. 1999, 24, 133–184. [Google Scholar]
- Pavón Núñez, M.; Hidalgo Triana, N.; Pérez Latorre, A.V. Aportaciones al conocimiento de las comunidades de Pistacia terebinthus L. y de Acer monspessulanum L. en el sur de la Península Ibérica. Lagascalia 2013, 33, 299–311. [Google Scholar]
- Rivas-Martínez, S.; Penas, A.; Díaz-González, T.E.; Ladero-Álvarez, M.; Asensi-Marfil, A.; Díez-Garretas, B.; Molero-Mesa, J.; Valle-Tendero, F.; Cano, E.; Costa-Talens, M.; et al. Mapa de series, geoseries y geopermaseries de vegetación de España. Parte I. Itinera Geobot. 2007, 17, 4–36. [Google Scholar]
- Rivas-Martínez, S.; Penas, A.; Díaz-González, T.E.; Ladero-Álvarez, M.; Asensi-Marfil, A.; Díez-Garretas, B.; Molero-Mesa, J.; Valle-Tendero, F.; Cano, E.; Costa-Talens, M.; et al. Mapa de series, geoseries y geopermaseries de vegetación de España. Parte II. Itinera Geobot. 2011, 18, 5–424. [Google Scholar]
- Rivas-Martínez, S.; Penas, A.; Díaz-González, T.E.; Ladero-Álvarez, M.; Asensi-Marfil, A.; Díez-Garretas, B.; Molero-Mesa, J.; Valle-Tendero, F.; Cano, E.; Costa-Talens, M.; et al. Mapa de series, geoseries y geopermaseries de vegetación de España. Parte II. Itinera Geobot. 2011, 18, 425–800. [Google Scholar]
- Salmerón-Sánchez, E.; Martínez-Nieto, M.I.; Martínez-Hernández, F.; Garrido-Becerra, J.A.; Mendoza-Fernández, A.J.; Gil de Carrasco, C.; Ramos-Miras, J.J.; Lozano, R.; Merlo, M.E.; Mota, J.F. Ecology, genetic diversity and phylogeography of the Iberian endemic plant Jurinea pinnata (Lag.) DC. (Compositae) on two special edaphic substrates: dolomite and gypsum. Plant Soil 2014, 374, 233–250. [Google Scholar] [CrossRef]
- Medina-Cazorla, J.M.; Garrido-Becerra, J.A.; Mendoza Fernández, A.; Pérez-García, F.J.; Salmerón, E.; Gil, C.; Mota Poveda, J.F. ’Biogeography of the Baetic ranges (SE Spain): A historical approach using cluster and parsimony analyses of endemic dolomitophytes’. Plant Biosyst. 2010, 144, 111–120. [Google Scholar] [CrossRef]
- Brković, D.L.; Tomović, G.M.; Niketić, M.S.; Lakušić, D.V. Diversity analysis of serpentine and non-serpentine flora—Or, is serpentinite inhabited by a smaller number of species compared to different rock types? Biologia 2015, 70, 61–74. [Google Scholar] [CrossRef]
- Mota, J.F.; Garrido-Becerra, J.A.; Pérez-García, F.J.; Salmerón-Sánchez, E.; Sánchez-Gómez, P.; Merlo, E. Conceptual baseline for a global checklist of gypsophytes. Lazaroa 2016, 37, 7–30. [Google Scholar]
- Del Río, S.; Herrero, L.; Fraile, R.; Penas, A. Spatial distribution of recent rainfall trends in Spain (1961–2006). Int. J. Climatol. 2011, 31, 656–667. [Google Scholar]
- Cano-Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Pinto Gomes, C.J.; Spampinato, G.; Cano, E. Taxonomy, ecology and distribution of Juniperus oxycedrus L. group in the Mediterranean Region using morphometric, phytochemical and bioclimatic approaches. BioRxiv 2018, 11, 1–23. [Google Scholar]
- Santana, N.C. Fire Recurrence and Normalized Difference Vegetation Index (NDVI) Dynamics in Brazilian Savanna. Fire 2019, 2, 1. [Google Scholar] [CrossRef]
- Spampinato, G.; Musarella, C.M.; Cano-ortiz, A.; Signorino, G. Habitat, occurrence and conservation status of the Saharo-Macaronesian and Southern-Mediterranean element Fagonia cretica L. (Zygophyllaceae) in Italy. J. Arid Land 2018, 10, 140–151. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, W.; Zhang, X.; Fang, X. Soil water storage changes within deep profiles under introduced shrubs during the growing season: evidence from semiarid Loess Plateau, China. Water 2016, 8, 475. [Google Scholar] [CrossRef]
- Liu, J.; Xiong, Y.; Tian, J.; Tan, Z. Spatiotemporal changes in evapotranspiration from an overexploited water resources basin in arid Northern China and their implications for ecosystem management. Sustainability 2019, 11, 445. [Google Scholar] [CrossRef]
- Cano, E.; Musarella, C.M.; Cano-Ortiz, A.; Piñar, J.C.; Pinto Gomes, C.J.; Rodríguez Torres, A.; Spampinato, G. A phytosociological review of siliceous sedges in C-W Spain and their state of conservation based on diversity indices. Plant Sociol. 2017, 54, 5–14. [Google Scholar]
- Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.M.; Pinto Gomes, C.J.; Spampinato, G.; Cano, E. Rupicolous habitats of interest for conservation in the central-southern Iberian Peninsula. Plant Sociol. 2017, 54, 29–42. [Google Scholar]
- Quinto-Canas, R.; Mendes, P.; Cano-Ortiz, A.; Musarella, C.M.; Pinto-Gomes, C. The Agrostion castellanae Rivas Goday 1957 corr. Rivas Goday & Rivas-Martínez 1963 alliance in the southwestern Iberian Peninsula. Plant Sociol. 2018, 55, 21–29. [Google Scholar]
- Spampinato, G.; Crisarà, R.; Cannavò, S.; Musarella, C.M. Phytotoponims of southern Calabria: A tool for the analysis of the landscape and its transformations. Atti Soc. Tosc. Sc. Nat. Mem. Serie B 2017, 124, 61–72. [Google Scholar]
- Vila-Viçosa, C.; Vázquez, F.; Mendes, P.; del Rio, S.; Musarella, C.M.; Cano-Ortiz, A.; Meireles, C. Syntaxonomic update on the relict groves of Mirbeck’s oak (Quercus canariensis Willd. and Q. marianica C. Vicioso) in southern Iberia. Plant Biosyst. 2015, 149, 512–526. [Google Scholar] [CrossRef]
- Gratani, L.; Varone, L.; Bonito, A. Carbon sequestration of four urban parks in Rome. Urban For. Urban Green. 2016, 19, 184–193. [Google Scholar] [CrossRef]
- Del Giudice, V.; Massimo, D.E.; De Paola, P.; Forte, F.; Musolino, M.; Malerba, A. Post Carbon City and Real Estate Market: Testing the Dataset of Reggio Calabria Market Using Spline Smoothing Semiparametric Method. In New Metropolitan Perspectives. ISHT 2018. Smart Innovation, Systems and Technologies; Calabrò, F., Della Spina, L., Bevilacqua, C., Eds.; Springer: Cham, Switzerland, 2019; pp. 206–214. [Google Scholar]
- De Paola, P.; Del Giudice, V.; Massimo, D.E.; Forte, F.; Musolino, M.; Malerba, A. Isovalore Maps for the Spatial Analysis of Real Estate Market: A Case Study for a Central Urban Area of Reggio Calabria, Italy. In New Metropolitan Perspectives. ISHT 2018. Smart Innovation, Systems and Technologies; Calabrò, F., Della Spina, L., Bevilacqua, C., Eds.; Springer: Cham, Switzerland, 2019; pp. 402–410. [Google Scholar]
- Malerba, A.; Massimo, D.E.; Musolino, M.; Nicoletti, F.; De Paola, P. Post Carbon City: Building Valuation and Energy Performance Simulation Programs. In New Metropolitan Perspectives. ISHT 2018. Smart Innovation, Systems and Technologies; Calabrò, F., Della Spina, L., Bevilacqua, C., Eds.; Springer: Cham, Switzerland, 2019; pp. 513–521. [Google Scholar]
- Massimo, D.E.; Del Giudice, V.; De Paola, P.; Forte, F.; Musolino, M.; Malerba, A. Geographically Weighted Regression for the Post Carbon City and Real Estate Market Analysis: A Case Study. In New Metropolitan Perspectives. ISHT 2018. Smart Innovation, Systems and Technologies; Calabrò, F., Della Spina, L., Bevilacqua, C., Eds.; Springer: Cham, Switzerland, 2019; pp. 142–149. [Google Scholar]
- Spampinato, G.; Massimo, D.E.; Musarella, C.M.; De Paola, P.; Malerba, A.; Musolino, M. Carbon sequestration by cork oak forests and raw material to built up post carbon city. In New Metropolitan Perspectives. ISHT 2018. Smart Innovation, Systems and Technologies; Calabrò, F., Della Spina, L., Bevilacqua, C., Eds.; Springer: Cham, Switzerland, 2019; pp. 663–671. [Google Scholar]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Nilsson, L.O.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. Forest. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Charro, E.; Moyano, A.; Cabezón, R. The potential of Juniperus thurifera to sequester carbon in semi-arid forest soil in spain. Forests 2017, 8, 330. [Google Scholar] [CrossRef]
- Rădulescu, C.; Toader, R.; Boca, G.; Abrudan, M.; Anghel, C.; Toader, D.C. Sustainable Development in Maramures County. Sustainability 2015, 7, 7622–7643. [Google Scholar] [CrossRef] [Green Version]
- Kurowska-Pysz, J.; Castanho, R.A.; Loures, L. Sustainable Planning of Cross-Border Cooperation: A Strategy for Alliances in Border Cities. Sustainability 2018, 10, 1416. [Google Scholar] [CrossRef]
- Castanho, R.A.; Loures, L.; Cabezas, J.; Fernández-Pozo, L. Cross-Border Cooperation (CBC) in Southern Europe—An Iberian Case Study. The Eurocity Elvas-Badajoz. Sustainability 2017, 9, 360. [Google Scholar] [CrossRef]
- Géhu, J.M.; Rivas-Martinez, S. Notions fondamentales de Phytosociologie. In Syntaxonomie; Dierschke, H., Ed.; Ber. Intern. Symposien IV–V; Cramer: Vaduz, Liechtenstein, 1981; pp. 5–33. [Google Scholar]








| Order no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| Area in m2 | 200 | 200 | 400 | 200 | 100 | 400 | 300 | 300 | 500 | 200 | 200 | ||
| Altitude in m 1 = 10 | 989 | 1243 | 423 | 1228 | 1004 | 1233 | 981 | 995 | 900 | 1021 | 1229 | ||
| Cover rate % | 15 | 55 | 75 | 60 | 30 | 60 | 25 | 20 | 60 | 25 | 25 | ||
| Orientation | - | - | E | - | - | N | NE | W | W | E | N | ||
| Slope % | - | - | 3 | - | - | 20 | 4 | 3 | 20 | 50 | 5 | ||
| Average veg. height (m.) | 1.2 | 0.5 | 0.9 | 1.5 | 1.3 | 1.8 | 1.4 | 1.5 | 1.5 | 1.3 | 1.5 | ||
| Cluster no. | GJ1 | GJ2 | GJ3 | GJ4 | GJ5 | GJ6 | GJ7 | GJ8 | GJ9 | GJ10 | GJ11 | ||
| Characteristics of Association and Higher Units | P | ||||||||||||
| Juniperus oxycedrus subsp. badia | 2 | 3 | 3 | 2 | 2 | 2 | 2 | 2 | 3 | 2 | 2 | 11 | |
| Genista polyanthos | 2 | 3 | 3 | 3 | 2 | 2 | 2 | 2 | 1 | 2 | 2 | 11 | |
| Quercus rotundifolia | + | 1 | - | 1 | + | + | + | + | 1 | + | 1 | 10 | |
| Phillyrea angustifolia | - | - | + | - | - | - | - | + | + | - | - | 3 | |
| Erica arborea | - | - | - | - | - | + | - | - | - | - | + | 2 | |
| Juniperus oxycedrus subsp. oxycedrus | + | - | - | - | - | - | - | - | - | - | - | 1 | |
| Pistacia lentiscus | - | - | 1 | - | - | - | - | - | - | - | - | 1 | |
| Asparagus acutifolius | - | - | + | - | - | - | - | - | - | - | - | 1 | |
| Companions | |||||||||||||
| Dianthus lusitanus | - | + | - | - | 1 | 1 | + | 1 | 1 | 1 | 1 | 8 | |
| Cistus ladanifer | + | 1 | + | - | - | - | + | + | + | + | 1 | 8 | |
| Jasione mariana | - | - | - | - | 1 | 1 | 2 | 1 | - | + | + | 6 | |
| Arrhenatherum bulbosum | - | - | - | 1 | + | - | + | + | - | 1 | + | 6 | |
| Linaria saxatilis | - | - | - | + | 1 | 1 | - | + | - | 1 | - | 5 | |
| Sedum brevifolium | - | - | - | - | + | - | 1 | 1 | + | + | - | 5 | |
| Halimium ocymoides | + | + | - | - | - | + | 1 | - | - | - | - | 4 | |
| Rosmarinus officinalis | + | - | + | - | - | - | - | + | + | - | - | 4 | |
| Asphodelus albus | - | - | - | 1 | - | + | + | + | - | - | - | 4 | |
| Rumex angiocarpus | - | - | - | + | + | - | + | - | - | - | - | 3 | |
| Thymus mastichina | - | - | + | + | - | - | - | - | - | - | - | 2 | |
| Urginea maritima | - | - | - | - | - | - | - | + | + | - | - | 2 | |
| Umbilicus rupestris | - | - | - | - | + | - | - | - | - | + | - | 2 | |
| Dactylis lusitanica | - | - | + | + | - | - | - | - | - | - | - | 2 | |
| Elymus caninus | - | - | - | - | - | - | + | - | - | + | - | 2 | |
| Digitalis mariana | - | - | - | - | - | - | - | - | - | + | 2 | 2 | |
| Other species:Cistus populifolius x C. salvifolius GJ1(1), Cistus albidus GJ3(+), Retama sphaerocarpa GJ3(+), Nerium oleander GJ3(1), Flueggea tinctoria GJ3(+), Corrigiola telephiifolia GJ3(+), Scirpus holoschoenus GJ3(+), Scrophularia canina GJ3(+), Fraxinus angustifolia GJ3(+), Festuca elegans GJ4(1), Elymus hispanicus GJ4(+), Armeria capitella GJ4(1), Sedum forsteranum GJ4(+), Petrorhagia nanteuillii GJ4(+), Halimium umbellatum subsp. viscosum GJ5(+), Poa bulbosa GJ6(+), Vulpia myuros GJ7(+), Lamarckia aurea GJ7(+), Mucizonia hispida GJ8(+), Dipcadi serotinum GJ8(+), Lavandula luisieri GJ8(+), Conopodium bourgaei GJ9(+), Asplenium billotii GJ9(+), Sedum dasyphyllum GJ10(+), Geranium purpureum GJ10(+), Anogramma leptophylla GJ10(+), Stipa gigantea GJ10(+), Conopodium capillifolium GJ10(+), Narcissus rupicola GJ10(+), Festuca rothmaleri GJ10(+), Coincya longirrostra GJ11(+), Genista florida GJ11(1). | |||||||||||||
| P = number of Presences; - = absent. Localities: GJ1. Mount Manzano. Near Torre Castañarejo (30S0449835/4256709); GJ2. Collado Grande. Torre Vigilancia (30S0443300/4257119); GJ3. Mouth of the Montoro River (30S0417604/4253109); GJ4. Pico Estrella. Finca Ruichoto (30S0448476/4250771); GJ5. Mount Manzano. Peñón de Atilano (30S0447322/4255301); GJ6. Abulagoso (30S0385725/4258301); GJ7. Cañada Real (30S0450280/4256964); GJ8. Piedrallana (30S0450483/4256122); GJ9. Mount Manzano (30S0450025/4257173); GJ10. Umbría Monroi. (30S0447629/4254779); GJ11. Collado Grande. Torre Vigilancia (30S0443228/4257178). | |||||||||||||
| Order no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | ||
| Area in m2 | 500 | 500 | 400 | 300 | 400 | 400 | 150 | 500 | 500 | 400 | 400 | 300 | 400 | 200 | 500 | 500 | 500 | 400 | 400 | 600 | 500 | 500 | 500 | 600 | ||
| Altitude in m 1 = 10 | 600 | 820 | 1019 | 1006 | 568 | 988 | 240 | 820 | 780 | 912 | 832 | 766 | 842 | 1141 | 300 | 451 | 480 | 650 | 503 | 391 | 280 | 300 | 420 | 600 | ||
| Cover rate % | 70 | 100 | 40 | 30 | 60 | 55 | 80 | 95 | 95 | 60 | 100 | 60 | 60 | 30 | 60 | 75 | 65 | 80 | 75 | 60 | 60 | 60 | 75 | 65 | ||
| Orientation | W | NE | S | SW | S | SW | NE | SW | SW | W | NW | S | S | S | SW | E | E | SW | E | E | NW | SW | NE | NE | ||
| Slope % | 40 | 20 | 25 | 75 | 60 | 60 | 10 | 25 | 30 | 15 | 10 | 12 | 20 | 90 | 25 | 30 | 35 | 20 | 95 | 40 | 40 | 25 | 25 | 15 | ||
| Average veg. height (m.) | 3.5 | 5 | 4 | 2.5 | 2.5 | 2.5 | 3 | 5 | 5 | 4 | 2.5 | 4 | 3 | 1.5 | 3.5 | 7.0 | 7.0 | 5 | 4.5 | 3:5 | 3 | 3 | 7.5 | 8.0 | ||
| Cluster no. | MJ1 | MJ2 | MJ3 | MJ4 | MJ5 | MJ6 | MJ7 | MJ8 | MJ9 | MJ10 | MJ11 | MJ12 | MJ13 | MJ14 | MJ15 | MJ16 | MJ17 | MJ18 | MJ19 | MJ20 | MJ21 | MJ22 | MJ23 | MJ24 | ||
| Characteristics of Association and Higher Units | P | |||||||||||||||||||||||||
| Juniperus oxycedrus subsp. badia | 3 | 4 | 2 | 2 | 3 | 3 | 3 | 5 | 4 | 3 | 5 | 3 | 3 | 4 | 3 | 4 | 3 | 3 | 2 | 1 | 4 | 4 | 3 | 4 | 24 | |
| Quercus rotundifolia | 1 | 2 | + | 1 | + | + | - | - | 1 | - | + | 1 | 1 | + | + | + | + | 1 | 1 | 1 | - | - | - | - | 17 | |
| Phillyrea angustifolia | 1 | 1 | - | + | 1 | 1 | 1 | 1 | - | - | + | 1 | 1 | - | - | 1 | 1 | 1 | + | + | - | - | - | - | 15 | |
| Pistacia terebinthus | + | + | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 2 | 1 | - | - | + | 1 | 2 | + | + | 11 | |
| Myrtus communis | - | - | - | - | - | - | - | - | - | + | 1 | 3 | 2 | 1 | - | 1 | - | 2 | - | - | 1 | 1 | 1 | 1 | 11 | |
| Pistacia lentiscus | 1 | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 1 | - | - | - | 1 | 1 | 1 | 2 | 2 | 9 | |
| Arbutus unedo | - | - | - | - | - | 1 | + | - | - | + | 1 | - | + | 1 | - | 1 | + | - | - | - | - | - | - | - | 8 | |
| Quercus coccifera | 1 | + | - | - | - | - | - | + | + | - | - | - | - | 1 | 1 | + | - | - | - | - | - | - | - | - | 7 | |
| Olea europea var. sylvestris | + | - | - | - | - | - | - | - | - | - | - | - | - | + | - | 2 | 2 | 3 | 4 | 3 | - | - | - | - | 7 | |
| Rhamnus oleoides | + | + | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | 2 | 2 | 2 | 2 | 7 | |
| Jasminum fruticans | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | + | 1 | 1 | 1 | 1 | 6 | |
| Erica arborea | - | - | - | - | - | - | - | - | - | + | + | + | 1 | - | - | + | - | - | - | - | - | - | - | - | 5 | |
| Asparagus albus | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | 1 | 2 | 1 | 1 | - | - | 5 | |
| Daphne gnidium | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | + | 1 | - | - | - | + | - | - | - | 4 | |
| Smilax aspera | - | - | - | - | - | - | - | - | - | - | - | + | - | + | - | - | - | - | - | + | - | - | + | - | 4 | |
| Phlomis purpurea | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | - | - | - | - | 1 | 2 | - | - | 4 | |
| Osyris alba | - | + | - | - | - | + | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | |
| Juniperus oxycedrus subsp. oxycedrus | + | - | - | - | 2 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | |
| Rhamnus alaternus | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | - | - | - | 1 | - | - | - | 3 | |
| Quercus broteroi | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + | - | - | - | - | - | - | - | 3 | |
| Asparagus acutifolius | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | 2 | |
| Thapsia villosa | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | 2 | |
| Phillyrea latifolia | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | 1 | - | - | - | 2 | |
| Asparagus aphyllus | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | - | - | 2 | |
| Rubia peregrina | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | 1 | |
| Viburnum tinus | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Crataegus monogyna | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | 1 | |
| Pyrus bourgaeana | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | 1 | |
| Teucrium fruticans | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | 1 | |
| Pistacia x saportae | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | 1 | |
| Quercus suber | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Quercus marianica | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Quercus canariensis | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Companions | ||||||||||||||||||||||||||
| Cistus ladanifer | - | - | - | + | + | + | 2 | - | 2 | 1 | + | 1 | 1 | 1 | 1 | - | 1 | + | + | + | 1 | - | - | - | 16 | |
| Rosmarinus officinalis | - | - | - | - | - | - | - | + | 2 | 1 | + | 2 | + | - | 1 | - | + | + | + | + | + | + | - | - | 13 | |
| Cistus albidus | + | - | - | - | - | + | - | - | - | - | - | - | - | 1 | 1 | + | 1 | - | - | + | + | + | + | + | 11 | |
| Cheilanthes maderensis | - | + | 1 | + | 1 | - | - | - | - | + | - | - | 1 | - | - | - | - | + | 1 | - | - | - | + | + | 10 | |
| Urginea maritima | - | + | - | - | - | - | - | - | 1 | + | + | + | + | + | - | - | - | + | - | + | - | - | - | - | 9 | |
| Dianthus lusitanus | - | - | 1 | + | - | + | - | - | + | - | - | - | - | 1 | 1 | - | - | - | 2 | + | - | - | - | - | 8 | |
| Lavandula sampaiana | - | - | - | - | + | + | + | + | + | - | - | - | - | - | - | 1 | - | - | 1 | - | - | + | - | - | 8 | |
| Umbilicus rupestris | - | + | 1 | - | + | + | - | - | - | + | - | - | + | - | - | - | - | + | + | - | - | - | - | - | 8 | |
| Sedum brevifolium | - | - | 1 | - | - | + | - | - | - | + | - | - | + | - | - | - | - | + | - | - | - | - | - | - | 5 | |
| Mucizonia hispida | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | - | - | + | + | - | - | - | - | - | 5 | |
| Asphodelus albus | - | - | - | - | - | + | 1 | + | + | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 5 | |
| Bryonia cretica | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + | - | - | - | - | + | + | 5 | |
| Astragalus lusitanicus | - | - | - | - | - | - | + | - | - | + | + | 1 | + | - | - | - | - | - | - | - | - | - | - | - | 5 | |
| Dactylis hispanica | - | + | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | 4 | |
| Jasione mariana | - | - | - | + | - | + | - | - | - | - | - | - | 1 | - | - | - | - | - | - | + | - | - | - | - | 4 | |
| Asplenium ceterach | - | + | 1 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | - | - | - | - | - | 4 | |
| Arrhenatherum bulbosum | - | - | - | - | - | + | - | - | + | - | - | + | + | - | - | - | - | - | - | - | - | - | - | - | 4 | |
| Halimium atriplicifolium | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | + | + | + | - | - | - | - | - | - | - | 4 | |
| Digitalis heywoodii var. albicans | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | + | + | - | - | - | - | - | - | - | 4 | |
| Sedum dasyphyllum | + | - | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | |
| Thymus mastichina | - | - | - | - | - | - | - | - | + | - | - | - | - | - | 1 | - | - | - | - | - | + | - | - | - | 3 | |
| Sanguisorba minor | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | + | + | 3 | |
| Tamus communis | - | - | + | - | - | - | - | - | - | - | + | - | - | - | - | - | 1 | - | - | - | - | - | - | - | 3 | |
| Nerium oleander | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | + | - | - | - | 1 | - | - | - | - | 3 | |
| Anogramma leptophylla | - | - | - | + | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | - | - | - | - | - | 3 | |
| Selaginella denticulata | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | + | + | 3 | |
| Cytisus scoparius subsp. bourgaei | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | 2 | - | - | 3 | |
| Micromeria graeca | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | + | - | + | 3 | |
| Halimium umbellatum subsp. viscosum | - | - | 1 | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | 3 | |
| Genista hirsuta | - | 1 | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | + | - | - | - | - | - | - | 3 | |
| Lavandula luisieri | - | - | - | - | - | - | - | + | - | - | - | + | + | - | - | - | - | - | - | - | - | - | - | - | 3 | |
| Cistus salvifolius | - | 1 | - | - | - | - | - | - | - | - | + | + | - | - | - | - | - | - | - | - | - | - | - | - | 3 | |
| Other species:Coincya longirrostra MJ7, MJ15(+), Sedum sediforme MJ1, MJ9(+), Retama sphaerocarpa MJ9, MJ20(+), Vitis vinifera subsp. sylvestris MJ14, MJ16(+), Adenocarpus argyrophyllus MJ4, MJ5(+), Erica scoparia MJ7(2), MJ8(+), Halimium ocymoides MJ7(1), MJ13(+), Cytisus striatus subsp. eriocarpus MJ2(1), MJ19(2), Adenocarpus telonensis MJ2(1), MJ8(+), Rubus ulmifolius MJ1(+), MJ20(+), Digitalis mariana MJ20, MJ21(+), Aristolochia baetica MJ21, MJ22(1), Jasonia glutinosa MJ1(+), Polygala rupestris MJ1(+), Helianthemum croceum MJ1(+), Fraxinus angustifolia MJ15(+), Antirrhinum graniticum subsp. onubensis MJ16(+), Flueggea tinctoria MJ21(+), Dianthus crassipes MJ21(1), Linaria saxatilis MJ6(+), Calluna vulgaris MJ13(1), Cheilanthes hispanica MJ4(+), Helichrysum serotinum MJ9(+), Teucrium gnaphalodes MJ2(+), Teucrium pseudochamaepitys MJ9(+), Cheilanthes tinaei MJ2(+), Stipa capensis MJ10(+), Hyacinthoides hispanica MJ3(+), Pterocepalus diandrus MJ13(+). | ||||||||||||||||||||||||||
| P = number of Presences; - = absent. Localities: MJ1. Mora de Toledo (Toledo) [19], (Stipo tenacissimae-Juniperetum lagunae); MJ2. Mora de Toledo (Toledo) [19], (Stipo tenacissimae-Juniperetum lagunae); MJ3. Collado Sierra de la Solana (30S0404372/4259526); MJ4. Puerto Viejo (30S0382806/4254598); MJ5. Crestones cuerda sierra Chillón (30S0334946/4289958); MJ6. Sierra de Solana (30S0404372/4259526); MJ7. Aldea Cerezo al Yeguas (Cardeña); MJ8. Puerto Lapice (C.Real) [19], (Stipo tenacissimae-Juniperetum lagunae); MJ9. Marjaliza (Toledo) [19], (Stipo tenacissimae-Juniperetum lagunae); MJ10. Sierrra Madrona (30S04067709/4250513); MJ11. Sierra Quintana (oeste) (30S0391176/4250840); MJ12. Swamp of the Garganta (30S0374625/4260280); MJ13. Camino Peña Escrita-Finca Valmayor (30S0387688/4255453); MJ14. Puerto Viejo (30S0382806/4254598); MJ15. Aldea Cerezo al Yeguas (Cardeña); MJ16. Aldea Cerezo al Yeguas (Cardeña); MJ17. Aldea Cerezo al Yeguas (Cardeña); MJ18. Near San Benito (30S0352613/4271155); MJ19. Easternmost Sierra Almadén (30S0346378/4291566); MJ20. Mouth of the Montoro River (30S0417854/4253846); MJ21. Garganta del Río Viar (Sevilla) [19], (Community of Phlomis purpurea); MJ22. Garganta del Río Viar (Sevilla) [19], (Community of Phlomis purpurea); MJ23. Garganta del Río Viar (Sevilla) [19], (Community of Phlomis purpurea); MJ24. Garganta del Río Viar (Sevilla) [19], (Community of Phlomis purpurea). | ||||||||||||||||||||||||||
| Order no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | |
| Area in m2 | 150 | 250 | 150 | 250 | 250 | 250 | 200 | 400 | 150 | 200 | 400 | 400 | 500 | 150 | 500 | 250 | 250 | 200 | 600 | 500 | 600 | |
| Altitude in m 1=10 | 50 | 105 | 95 | 69 | 105 | 95 | 115 | 135 | 140 | 115 | 115 | 120 | 105 | 110 | 105 | 113 | 120 | 157 | 130 | 100 | 95 | |
| Cover rate % | 40 | 70 | 80 | 65 | 60 | 60 | 40 | 65 | 50 | 75 | 65 | 65 | 70 | 45 | 65 | 70 | 75 | 60 | 60 | 60 | 60 | |
| Orientation | N | W | SW | S | N | S | N | E | NW | E | SE | SE | S | SW | S | SW | SW | S | S | SW | SW | |
| Slope % | 40 | 15 | 25 | 15 | 15 | 25 | 25 | 20 | 15 | 15 | 14 | 10 | 20 | 25 | 20 | 15 | 15 | 20 | 45 | 20 | 15 | |
| Average veg. height (m.) | 2.0 | 2.5 | 2.5 | 2.0 | 2.0 | 2.5 | 3.0 | 2.5 | 1.8 | 4.0 | 2.5 | 4.0 | 2.5 | 2.0 | 2.5 | 2.5 | 2.0 | 2.0 | 4.0 | 2.0 | 1.8 | |
| Cluster no. | JPB1 | JPB2 | JPB3 | JPB4 | JPB5 | JPB6 | JPB7 | JPB8 | JPB9 | JPB10 | JPB11 | JPB12 | JPB13 | JPB14 | JPB15 | JPB16 | JPB17 | JPB18 | JPB19 | JPB20 | JPB21 | |
| Characteristics of Association and Higher Units | P | |||||||||||||||||||||
| Juniperus phoenicea | 1 | - | - | 1 | 1 | 2 | 2 | + | 3 | 4 | 3 | 3 | 3 | 1 | 2 | 3 | 3 | 3 | 2 | 3 | 1 | 19 |
| Juniperus oxycedrus subsp. badia | + | 3 | 4 | 3 | 3 | 3 | 3 | 3 | 1 | 1 | + | 2 | 3 | 2 | 3 | 3 | 3 | 2 | 3 | 3 | 3 | 21 |
| Juniperus oxycedrus subsp. oxycedrus | - | 1 | 1 | - | - | - | - | 2 | 2 | 1 | 1 | 1 | 2 | 2 | 3 | 2 | 2 | + | - | 1 | 3 | 15 |
| Quercus rotundifolia | - | + | - | - | + | 1 | + | + | + | + | 1 | + | - | - | - | 1 | + | + | + | + | + | 15 |
| Pistacia terebinthus | - | - | + | - | - | - | - | - | - | + | - | + | - | + | - | - | - | - | + | - | - | 5 |
| Pistacia lentiscus | 1 | - | - | 1 | - | - | - | - | - | - | - | - | 1 | + | - | - | - | - | - | - | - | 4 |
| Quercus coccifera | 1 | - | - | + | 1 | 1 | - | - | - | - | + | - | 1 | + | - | - | - | - | - | - | - | 7 |
| Asparagus acutifolius | + | - | - | + | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | 3 |
| Jasminum fruticans | + | - | - | - | - | + | - | - | - | - | - | - | - | 1 | 1 | - | - | - | - | - | - | 4 |
| Daphne gnidium | - | + | - | 1 | + | - | 1 | - | - | - | 1 | - | + | - | + | + | - | - | 1 | + | + | 11 |
| Quercus faginea | - | + | - | 1 | - | - | + | - | - | - | + | - | - | - | - | - | - | - | + | - | - | 5 |
| Thapsia villosa | - | + | - | - | + | - | + | + | + | - | + | + | - | - | - | + | - | + | - | - | - | 9 |
| Carex hallerana | - | - | 1 | - | - | - | - | - | - | - | - | - | + | - | + | - | - | - | - | 1 | - | 4 |
| Rhamnus myrtifolius | - | - | 1 | - | - | - | - | 1 | - | - | - | + | - | - | - | - | - | 2 | - | - | - | 4 |
| Pinus halepensis | - | - | - | + | - | - | - | - | - | - | - | + | - | + | - | - | - | - | - | - | - | 3 |
| Phillyrea latifolia | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | 1 |
| Pinus salzmannii | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | + | - | 2 |
| Lonicera periclymenum subsp. hispanica | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | 1 |
| Ptilostemon hispanicus | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | + | - | - | 2 |
| Hedera ibernica | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | - | - | - | 2 |
| Rubia peregrina | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | 2 |
| Bupleurum rigidum | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | 1 |
| Ilex aquifolium | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | 1 |
| Clematis flammula | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | 1 |
| Rhamnus alaternus | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Smilax aspera | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Teline patens | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Melica minuta | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | 1 |
| Paeonia broteroi | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Companions | ||||||||||||||||||||||
| Rosmarinus officinalis | + | 2 | 2 | - | 1 | 1 | - | - | - | - | - | - | + | + | - | 1 | 1 | - | 1 | + | - | 11 |
| Thymus orospedanus | - | 1 | 1 | - | - | + | 2 | 2 | - | - | - | + | - | - | - | + | + | - | + | - | + | 10 |
| Asphodelus albus | - | + | + | - | - | - | 1 | + | + | + | 1 | + | - | - | - | + | - | - | - | - | - | 9 |
| Phlomis lychnitis | - | - | - | - | - | - | + | 1 | + | + | + | 1 | - | + | + | - | - | - | - | - | + | 9 |
| Brachypodium retusum | - | - | - | - | + | + | - | + | - | + | - | 1 | 1 | - | + | - | - | - | - | + | + | 9 |
| Thymus mastichina | - | - | - | + | - | - | 1 | - | + | + | 2 | 1 | + | - | - | - | 1 | - | - | - | + | 9 |
| Cistus albidus | - | - | - | - | - | 1 | - | 2 | + | - | - | + | + | + | - | 1 | - | - | - | - | - | 7 |
| Dactylis hispanica | - | - | - | - | + | + | - | + | + | - | - | - | + | + | - | - | - | - | - | - | - | 6 |
| Lavandula latifolia | - | 1 | + | - | 1 | + | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | 5 |
| Thymus zygis subsp. gracilis | - | 1 | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 1 | - | - | - | - | + | - | 5 |
| Cistus monspeliensis | - | + | - | 2 | - | + | - | - | - | - | - | - | + | + | - | - | - | - | - | - | - | 5 |
| Teucrium capitatum | - | + | - | - | - | - | - | + | 1 | - | - | - | - | - | - | - | + | - | - | - | - | 4 |
| Crataegus laciniata | + | - | - | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | 4 |
| Aphyllantes monspeliensis | - | - | - | + | - | - | - | - | + | - | 1 | - | - | - | - | - | - | - | + | - | - | 4 |
| Urginea maritima | - | - | - | - | - | - | - | + | - | - | - | + | - | + | + | - | - | - | - | - | - | 4 |
| Halimium atriplicifolium | - | 1 | - | + | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 |
| Eryngium campestre | - | - | - | - | - | - | + | + | + | - | - | - | - | - | - | - | - | - | - | - | - | 3 |
| Teucrium rotundifolium | - | - | + | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | 2 |
| Rosa canina | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | 2 |
| Berberis hispanica | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | 1 | - | - | - | 2 |
| Retama sphaerocarpa | - | - | - | + | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Helianthemum syriacum | - | - | - | - | - | - | 2 | + | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Helianthemum cinereum subsp. rotundifolium | - | - | - | - | - | - | - | - | + | + | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Erinacea anthyllis | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | + | - | 2 |
| Helictotrichon filifolium | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | + | - | - | - | 2 |
| Genista scorpius | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | 1 | 2 |
| Crocus serotinus subsp. salzmannii | - | - | - | - | - | - | - | - | - | - | - | - | + | - | + | - | - | - | - | - | - | 2 |
| Genista boissieri | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | + | - | 2 |
| Santolina canescens | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | + | 2 |
| Other species:Rubus ulmifolius JPB1(+), Phagnalon saxatile JPB1(+), Linum tenue JPB3(1), Helianthemum ledifolium JPB3(+), Buxus sempervirens JPB9(2), Filipendula vulgaris JPB7(+), Plantago lanceolata JPB7(+), Ornithogalum narbonense JPB7(+), Polygala monspeliaca JPB7(+), Ophrys tenthrediniferfa JPB7(+), Anthyllis vulneraria subsp. maura JPB2(+), Arrhenatherum album JPB8(+), Fumana paradoxa JPB9(1), Lotus corniculatus JPB9(1), Sanguisorba minor JPB9(+), Crupina crupinastrum JPB10(+), Helleborus foetidus JPB10(+), Geranium purpureum JPB11(+), Narcissus assoanus JPB11(+), Scorpiurus muricatus JPB12(+), Sherardia arvensis JPB12(+), Medicago rigidula JPB12(+), Crepis vesicaria subsp. haenseleri JPB12(+), Bellis perennis JPB12(+), Echinaria capitata JPB12(+), Poa annua JPB12(+), Rumex bucephalophorus JPB12(+), Verbuscum giganteum JPB14(+), Prunus spinosa JPB14(+), Asplenium ceterach JPB15(+), Cytisus scoparius subsp. reverchonii JPB16(+), Fumana laevipes JPB16(+), Paronychia argentea JPB16(+), Amelanchier ovalis JPB16(1), Muscari giennense JPB17(+), Rhagadiolus stellatus JPB17(+), Thymus granatensis JPB18(+), Biscutella sempervirens JPB18(+), Staehelina dubia JPB20(+), Linum suffruticosum JPB19(+), Fumana thymifolia JPB20(+), Lonicera splendida JPB20(+), Leuzea conifera JPB20(+). | ||||||||||||||||||||||
| P = number of Presences; - = absent. Localities: JPB1.—Casilla de los Rajones. JPB2 and JPB5.—Alto de las Muelas. JPB3.—Los Yegüerizos. JPB4.—Bujaraiza. JPB6.—Los Palancares. JPB7.—La Fresnedilla. JPB8.—Collado de la Traviesa. JPB9 and JPB18.—Lancha de la Escalera. JPB10.—La Morra. JPB11.—Fuente del Milano. JPB12.—Hoya de Miguel Barba. JPB13 and JPB17.—Vilches. JPB14.—La Canaleja. JPB15.—Aguascebas Reservoir. JPB16.—San Antón. JPB19.—Rise to the birth of the Guadalquivir. JPB20 and JPB21.—Highway Quesada-Pozo Alcón. | ||||||||||||||||||||||
| Order no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Area in m2 | 300 | 250 | 200 | 300 | 200 | 250 | 250 | 300 | 200 | 250 | 300 | |
| Altitude in m 1 = 10 | 60 | 57 | 65 | 95 | 74 | 85 | 95 | 103 | 105 | 105 | 130 | |
| Cover rate % | 60 | 60 | 70 | 100 | 80 | 100 | 100 | 70 | 70 | 60 | 70 | |
| Orientation | N | NE | N | SE | N | N | NW | N | S | S | S | |
| Slope % | 5 | 15 | 5 | 5 | 30 | 10 | 20 | 5 | 2 | 25 | 40 | |
| Average veg. height (m.) | 3.0 | 2.0 | 2.0 | 8.0 | 2.5 | 4.0 | 4.5 | 3.5 | 2.0 | 3.0 | 3.0 | |
| Cluster no. | TP1 | TP2 | TP3 | TP4 | TP5 | TP6 | TP7 | TP8 | TP9 | TP10 | TP11 | |
| Characteristics of Association and Higher Units | P | |||||||||||
| Pistacia terebinthus | 3 | 3 | 3 | 5 | 4 | 5 | 5 | 3 | 4 | 3 | 3 | 11 |
| Juniperus oxycedrus subsp. badia | + | 1 | - | - | + | - | 1 | 1 | 1 | 2 | 1 | 8 |
| Quercus coccifera | + | + | 1 | 1 | 2 | + | + | - | 1 | - | - | 8 |
| Quercus rotundifolia | + | - | + | + | 1 | 1 | 1 | 1 | + | + | - | 9 |
| Jasminum fruticans | 1 | - | + | - | + | + | + | - | - | - | 1 | 6 |
| Pistacia lentiscus | + | + | 1 | 2 | + | - | - | + | - | - | - | 6 |
| Teline patens | - | 2 | + | - | + | 1 | 2 | - | + | - | - | 6 |
| Asparagus acutifolius | + | + | - | 1 | - | - | - | - | - | - | - | 3 |
| Rhamnus alaternus | 1 | 1 | - | - | - | - | + | - | - | - | - | 3 |
| Quercus faginea | - | - | - | - | - | - | - | + | 1 | + | - | 3 |
| Thapsia villosa | - | - | - | + | - | - | - | - | + | - | - | 2 |
| Smilax aspera | - | + | - | 1 | - | 1 | - | - | - | - | - | 3 |
| Tamus communis | + | 1 | 1 | - | - | - | - | - | - | - | + | 4 |
| Ruscus aculeatus | - | + | + | - | - | - | - | + | - | - | - | 3 |
| Arbutus unedo | - | - | + | - | + | 1 | - | + | - | - | - | 4 |
| Juniperus phoenicea | + | 2 | - | - | - | - | - | - | - | - | - | 2 |
| Hedera ibernica | + | - | - | - | - | 2 | - | - | - | - | - | 2 |
| Rubia peregrina | - | - | + | - | - | + | + | - | - | - | - | 3 |
| Phillyrea latifolia | - | + | 1 | - | - | - | - | - | - | - | - | 2 |
| Lonicera periclymenum subsp. hispanica | - | - | - | - | 1 | 1 | - | - | - | - | - | 2 |
| Phillyrea angustifolia | - | - | - | - | - | - | - | + | - | - | - | 1 |
| Olea europea var. sylvestris | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Viburnum tinus | - | - | - | - | - | 1 | - | - | - | - | - | 1 |
| Pinus pinaster | - | - | - | - | - | - | + | - | - | - | - | 1 |
| Coronilla glauca | 2 | - | - | - | - | - | - | - | - | - | - | 1 |
| Rhamnus myrtifolius | - | - | - | - | - | - | - | - | - | - | + | 1 |
| Companions | ||||||||||||
| Rosmarinus officinalis | - | + | 1 | 1 | 1 | - | 1 | - | 1 | 1 | - | 7 |
| Cistus albidus | + | + | - | + | 1 | - | - | 1 | + | 1 | - | 7 |
| Rubus ulmifolius | + | 1 | + | - | - | + | + | - | - | - | - | 5 |
| Thymus orospedanus | + | - | - | - | + | - | - | 1 | + | + | - | 5 |
| Aphyllantes monspeliensis | - | - | - | + | - | - | - | + | + | + | - | 4 |
| Brachypodium retusum | - | - | - | - | + | - | - | - | - | + | 1 | 3 |
| Clematis vitalba | + | - | - | - | - | - | 1 | + | - | - | - | 3 |
| Halimium atriplicifolium | - | - | - | - | - | - | - | - | 2 | 1 | - | 2 |
| Thymus zygis subsp. gracilis | - | + | 1 | - | - | - | - | - | - | - | - | 2 |
| Asphodelus albus | - | - | - | - | - | - | - | + | + | - | - | 2 |
| Thymus mastichina | - | - | - | - | - | - | - | 1 | - | - | 1 | 2 |
| Rosa canina | - | - | - | - | - | 1 | + | - | - | - | - | 2 |
| Lotus corniculatus | - | - | - | - | - | - | - | - | + | + | - | 2 |
| Euphorbia characias | + | - | + | - | - | - | - | - | - | - | - | 2 |
| Staehelina dubia | - | + | - | + | - | - | - | - | - | - | - | 2 |
| Lithodora fruticosa | - | - | - | - | - | - | - | - | + | + | - | 2 |
| Other species:Polygala rupestris TP11(+), Melica minuta TP11(1), Sedum sediforme TP11(+), Dactylis hispanica TP8(+), Crataegus laciniata TP3(+), Phlomis lychnitis TP4(+), Urginea maritima TP11(+), Helleborus foetidus TP11(+), Geranium purpureum TP3(+), Asplenium ceterach TP11(+), Vitis vinifera subsp. sylvestris TP2(+), Ficus carica TP2(+), Cardamine hirsuta TP3(+), Vincetoxicum nigrum TP3(+), Genista cinerea subsp. speciosa TP4(1), Melica magnolii TP5(+), Fraxinus angustifolia TP6(+), Dorycnium pentaphyllum TP7(+), Linum narbonense TP10(+), Sedum brevifolium TP11(+), Mucizonia hispida TP11(+), Asplenium trichomanes TP11(+), Biscutella valentina TP11(1), Rubus caesius TP11(+), Ruta chalepensis TP11(+). | ||||||||||||
| P = number of Presences; - = absent. Localities: 1 and 2.—Near Presa del Tranco. 3.—Puntal del Poyo Gonzalo. 4.—Cerro de los Cabezones. 5.—Ravine of the Obispo. 6.—Los Blancores. 7.—Near Finca de Mihí. 8.—Aguascebas Reservoir. 9.—Mount of Solegas Anchas. 10.—Cañada de los Caballeros. 11.—Rise to the birth of the Guadalquivir. | ||||||||||||
| Weather Station | PP | TP | IO | Ombrotype | ETP | e | Ioex1 | Ioex2 * | Ioex3 | Ombroclimatic Behaviour of the Locality |
|---|---|---|---|---|---|---|---|---|---|---|
| Almadén-Minas (CR) | 625.2 | 194.4 | 3.21 | dry | 808.54 | 161.7 | 0.59 | 1.19 | 1.78 | Semiarid |
| Cabezas Rubias (H) | 993.4 | 177.6 | 5.59 | subhumid | 702.14 | 140.42 | 1.2 | 2.4 | 3.6 | dry |
| Aracena (H) | 1025.8 | 175.2 | 5.85 | subhumid | 703.46 | 140.69 | 1.26 | 2.52 | 3.78 | dry |
| Santiago Pontones (J) | 1148.7 | 164.4 | 6.98 | subhumid | 675.23 | 135.04 | 1.54 | 3.08 | 4.62 | dry |
| Vadillo Castril (J) | 1182.2 | 140.4 | 8.42 | humid | 488.88 | 97.72 | 1.93 | 3.86 | 5.79 | subhumid |
| Grazalema (Ca) | 1962.2 | 183.6 | 10.7 | humid | 726.22 | 145.24 | 2.47 | 4.94 | 7.42 | subhumid |
| Montoro (Co) | 522.4 | 210 | 2.48 | dry | 903.15 | 180.63 | 0.4 | 0.81 | 1.22 | arid |
| Pozoblanco (Co) | 514.4 | 193.2 | 2.66 | dry | 805.45 | 161.09 | 0.45 | 0.91 | 1.37 | arid |
| Villanueva del Arzobispo (J) | 698.2 | 196.8 | 3.54 | dry | 915.7 | 183.14 | 0.65 | 1.3 | 1.96 | semiarid |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano, E.; Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Rodríguez Torres, A.; Del Río González, S.; Pinto Gomes, C.J.; Quinto-Canas, R.; Spampinato, G. Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern Iberian Peninsula. Sustainability 2019, 11, 1111. https://doi.org/10.3390/su11041111
Cano E, Musarella CM, Cano-Ortiz A, Piñar Fuentes JC, Rodríguez Torres A, Del Río González S, Pinto Gomes CJ, Quinto-Canas R, Spampinato G. Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern Iberian Peninsula. Sustainability. 2019; 11(4):1111. https://doi.org/10.3390/su11041111
Chicago/Turabian StyleCano, Eusebio, Carmelo M. Musarella, Ana Cano-Ortiz, José C. Piñar Fuentes, Alfonso Rodríguez Torres, Sara Del Río González, Carlos J. Pinto Gomes, Ricardo Quinto-Canas, and Giovanni Spampinato. 2019. "Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern Iberian Peninsula" Sustainability 11, no. 4: 1111. https://doi.org/10.3390/su11041111
APA StyleCano, E., Musarella, C. M., Cano-Ortiz, A., Piñar Fuentes, J. C., Rodríguez Torres, A., Del Río González, S., Pinto Gomes, C. J., Quinto-Canas, R., & Spampinato, G. (2019). Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern Iberian Peninsula. Sustainability, 11(4), 1111. https://doi.org/10.3390/su11041111