Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics




Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the Rice Endophytes
2.2. Identification of Bacteria
2.3. Phylogenic Tree
2.4. Indole Acetic Acid (IAA) Production
2.5. Potential for Nitrogen-Fixing and Phosphate-Solubilizing Ability
2.6. Fungicides on Bacteria Growth
2.7. Endophytic Colonization and the Effect on Rice Growth
2.8. IAA Production in MN1 Culture Medium with Different Tryptophan Concentration
2.9. The Effect of Fungicides on IAA Production by MN1
2.10. Statistical Analysis
3. Results and Discussion
3.1. Endophytes’ Isolation with Application of Fungicides
3.2. Fungicide Tolerance and Rice Inoculation Test
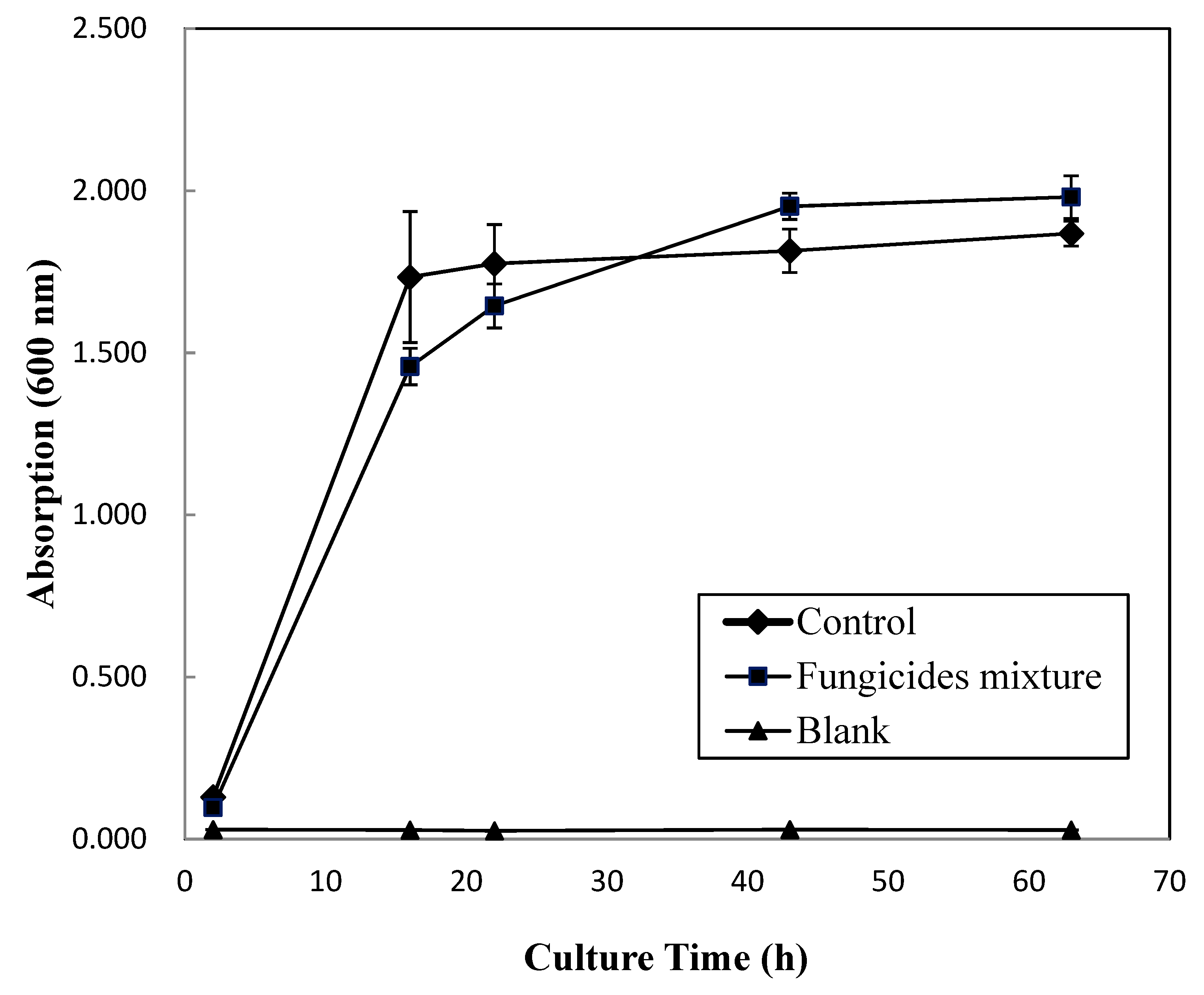
3.3. The Growth Curve of MN1 under Fungicides Application
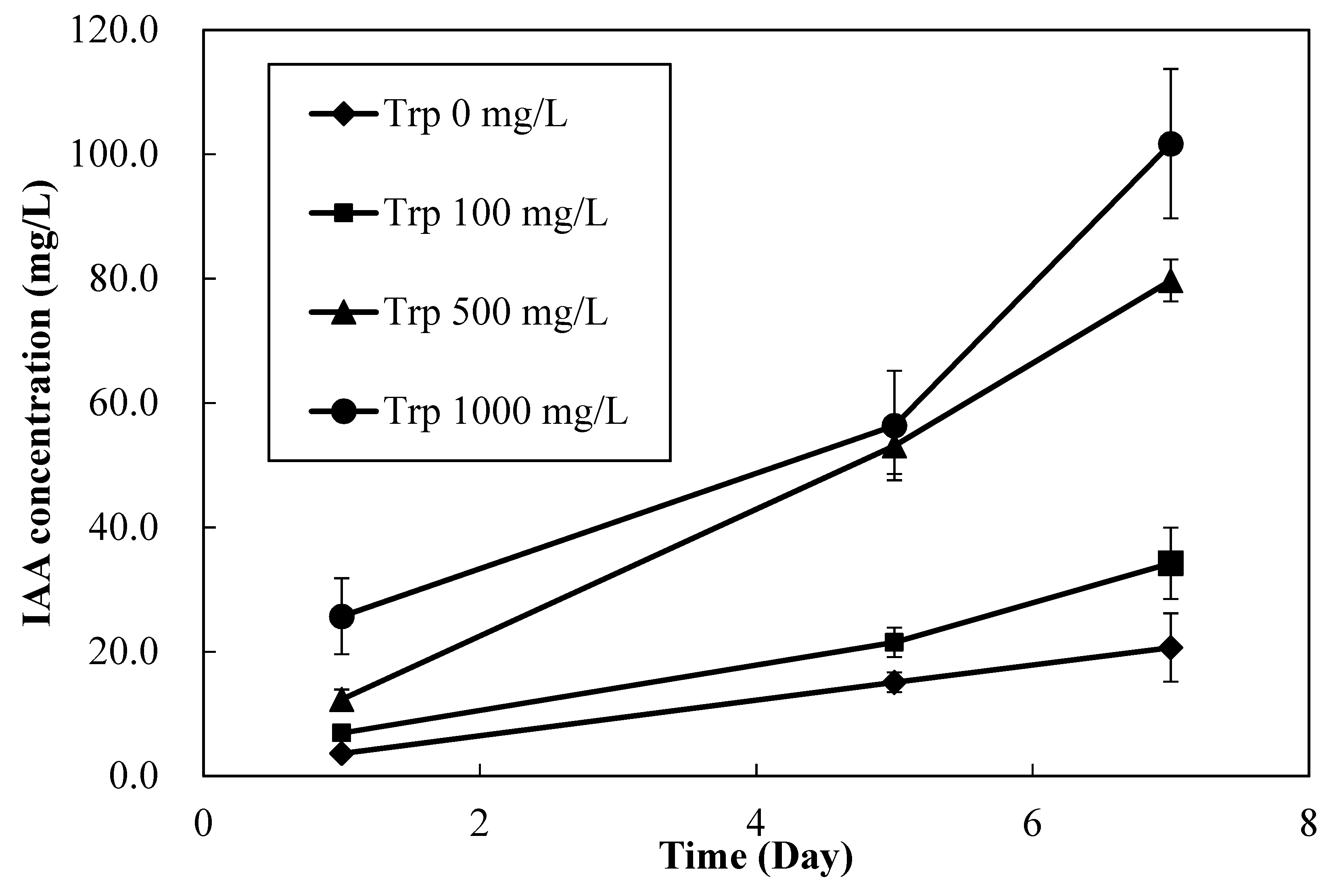
3.4. Tryptophan Dose-Dependent IAA Production Rates of MN1
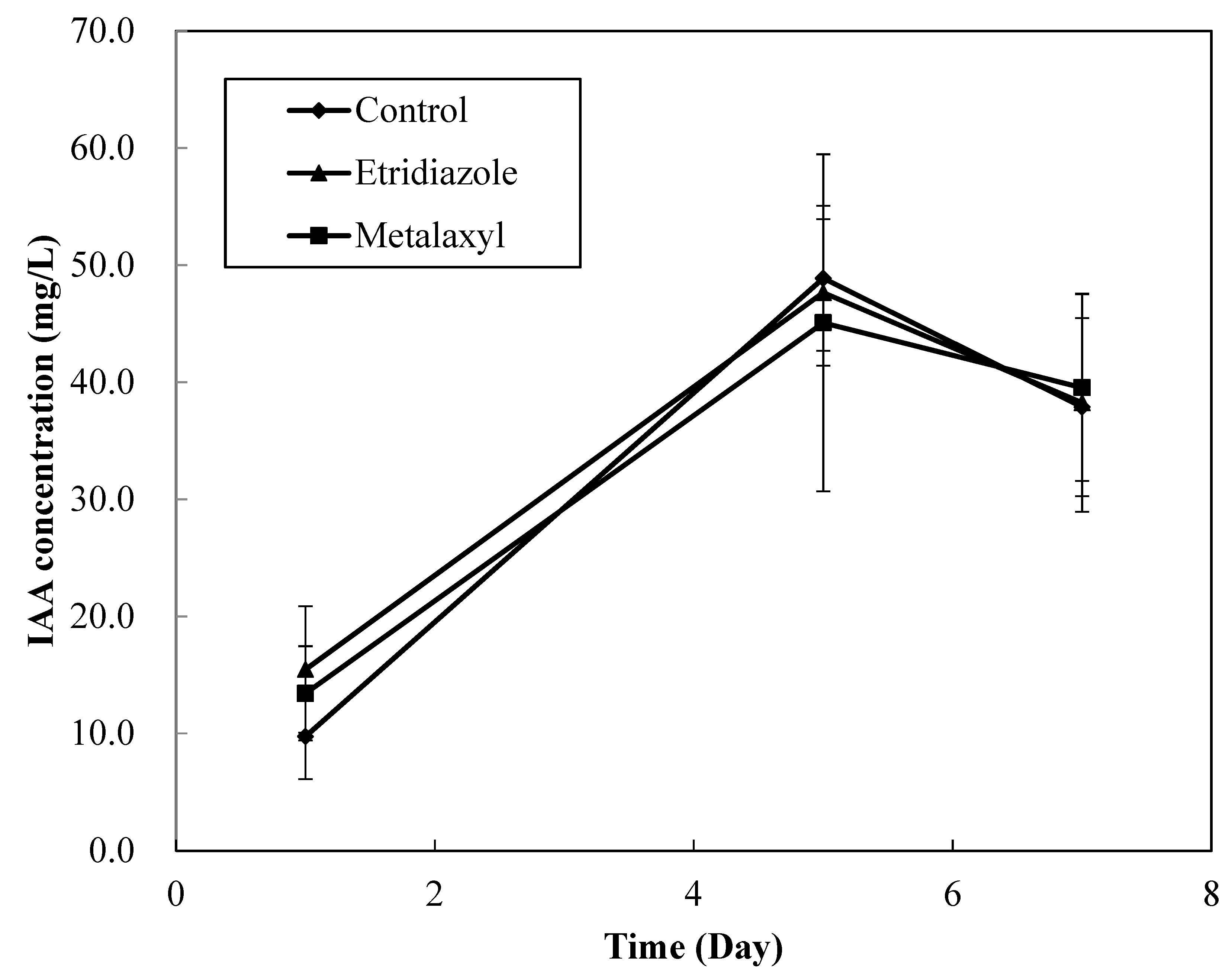
3.5. Effect of Fungicides on IAA Production of MN1
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Peng, S.; Tang, Q.; Zou, Y. Current Status and Challenges of Rice Production in China. Plant Prod. Sci. 2009, 12, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–687. [Google Scholar] [CrossRef] [PubMed]
- Pillay, V.K.; Nowak, J. Inoculum density, temperature, and genotype effects on in vitro growth promotion and epiphytic and endophytic colonization of tomato (Lycopersicon esculentum L.) seedlings inoculated with a pseudomonad bacterium. Can. J. Microbiol. 1997, 43, 354–361. [Google Scholar] [CrossRef]
- Hurek, T.; Reinhold-Hurek, B. Azoarcus sp. strain BH72 as a model for nitrogen-fixing grass endophytes. J. Biotechnol. 2003, 106, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Balandreau, J.; Kwon, S.W.; Weon, H.Y.; Lakshminarasimhan, C. Effects of the inoculation of Burkholderia vietnamensis and related endophytic diazotrophic bacteria on grain yield of rice. Microb. Ecol. 2008, 55, 21–37. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Hardoim, C.C.P.; van Overbeek, L.S.; van Elsas, J.D. Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS ONE 2012, 7, e30438. [Google Scholar] [CrossRef]
- Etesami, H.; Hosseini, H.M.; Alikhani, H.A. Bacterial biosynthesis of 1-aminocyclopropane-1-caboxylate (ACC) deaminase, a useful trait to elongation and endophytic colonization of the roots of rice under constant flooded conditions. Physiol. Mol. Biol. Plants 2014, 20, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Ahemad, M.; Khan, M.S. Effect of fungicides on plant growth promoting activities of phosphate solubilizing Pseudomonas putida isolated from mustard (Brassica compestris) rhizosphere. Chemosphere 2012, 86, 945–950. [Google Scholar] [CrossRef]
- Radzuhn, B.; Lyr, H. On the mode of action of the fungicide etridiazole. Pestic. Biochem. Physiol. 1984, 22, 14–23. [Google Scholar] [CrossRef]
- Yang, C.; Hame, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of action and possible impact on nontarget microorganisms. ISRN Ecol. 2011, 2011, 1–8. [Google Scholar] [CrossRef]
- Fisher, D.J.; Hayes, A.L. Mode of action of the systemic fungicides furalaxyl, metalaxyl, and ofurace. Pestic. Sci. 1982, 13, 330–339. [Google Scholar] [CrossRef]
- Krieger, M.S.; Cook, W.L.; Kennard, L.M. Extraction of tricyclazole from soil and sediment with subcritical water. J. Agric. Food Chem. 2000, 48, 2178–2183. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.F.; Gossen, B.D.; Chang, K.F.; Turnbull, G.D.; Howard, R.J. Effect of seed damage and metalaxyl seed treatment on pythium seedling blight and seed yield of field pea. Can. J. Plant Sci. 2001, 81, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Sultan, N.S.; Raipat, B.S.; Sinha, M.P. Survival of plant growth promoting rhizobacteria (PGPR) in soil under pesticide stress. ISESCO J. Sci. Technol. 2013, 9, 82–88. [Google Scholar]
- Najam-ul-Sehar Ahmad, M.; Akhtar, M.F.Z.; Jamil, M.; Latif, M.; Ahmad, I. Pesticide tolerant plant growth promoting rhizobacteria isolated from rhizosphere of okra. Soil Environ. 2015, 34, 111–118. [Google Scholar]
- Council of Agriculture Executive Yuan, R.O.C. (Taiwan). Plant Protection Manual. 2018. Available online: https://otserv2.tactri.gov.tw/ppm/ (accessed on 15 November 2018).
- Martinez, C.; Levesque, C.A.; Belanger, R.R.; Tweddell, R.J. Evaluation of fungicides for the control of carrot cavity spot. Pest Manag. Sci. 2005, 61, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Kunova, A.; Pizzatti, C.; Cortesi, P. Impact of tricyclazole and azoxystrobin on growth, sporulation and secondary infection of the rice blast fungus, Magnaporthe oryzae. Pest Manag. Sci. 2013, 69, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Zhan, F.; He, Y.; Yang, Y.; Li, Y.; Li, T.; Zhao, Z. Effects of tricyclazole on cadmium tolerance and accumulation characteristics of a dark septate endophyte (DSE), Exophiala pisciphila. Bull. Environ. Contam. Toxicol. 2016, 96, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Felsenstein, J. Confidence-limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.; Weber, R.P. The colorimetric estimation of IAA. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef]
- Brick, J.M.; Bostock, R.M.; Silversone, S.E. Rapid in situ assay for indole acetic acid production by bacteria immobilized on nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar]
- Eckford, R.; Cook, F.D.; Saul, D.; Aislabie, J.; Foght, J. Free-living heterotrophic nitrogen-fixing bacteria isolated from fuel-contaminated Antarctic soils. Appl. Environ. Microbiol. 2012, 68, 5181–5185. [Google Scholar] [CrossRef]
- Ladha, J.K.; Barraquio, W.L.; Watanabe, I. Isolation and identification of nitrogen fixing Enterobacter cloacae and Klebsiella planticola associated with rice plants. Can. J. Microbiol. 1983, 29, 1301–1308. [Google Scholar] [CrossRef]
- Colnaghi, R.; Green, A.; He, L.; Rudnick, P.; Kennedy, C. Strategies for increased ammonium production in free-living or plant associated nitrogen-fixing bacteria. Plant Soil 1997, 194, 145–154. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Microbiologiya 1948, 17, 362–370. [Google Scholar]
- Szkop, M.; Bielawski, W. A simple method for simultaneous RP-HPLC determination of indolic compounds related to bacterial biosynthesis of indole-3-acetic acid. Antonie Van Leeuwenhoek 2013, 103, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, G.A.; Ashworth, J. Bacteriostatic action of nitrification inhibitors. Can. J. Microbiol. 1982, 28, 1093–1100. [Google Scholar] [CrossRef]
- Scheldeman, P.; Rodríguez-Díaz, M.; Goris, J.; Pil, A.; De Clerck, E.; Herman, L.; De Vos, P.; Logan, N.A.; Heyndrickx, M. Bacillus farraginis sp. nov., Bacillus fortis sp. nov. and Bacillus fordii sp. nov., isolated at dairy farms. Int. J. Syst. Evol. Microbiol. 2004, 54, 1355–1364. [Google Scholar] [CrossRef]
- Shivaji, S.; Chaturvedi, P.; Begum, Z.; Pindi, P.K.; Manorama, R.; Padmanaban, D.A.; Shouche, Y.S.; Pawar, S.; Vaishampayan, P.; Dutt, C.B.; et al. Janibacter hoylei sp. nov., Bacillus isronensis sp. nov. and Bacillus aryabhattai sp. nov., isolated from cryotubes used for collecting air from the upper atmosphere. Int. J. Syst. Evol. Microbiol. 2009, 59, 2977–2986. [Google Scholar] [CrossRef]
- Silva, M.P.; Pereira, C.A.; Junqueira, J.C.; Jorge, A.O.C. Methods of destroying bacterial spores. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 490–496. [Google Scholar]
- Boricha, H.; Fulekar, M.H. Pseudomonas plecoglossicida as a novel organism for the bioremediation of cypermethrin. Biol. Med. 2009, 1, 1–10. [Google Scholar]
- Rahmoune, B.; Morsli, A.; Khelifi-Slaoui, M.; Khelifi, L.; Strueh, A.; Erban, A.; Kopka, J.; Prell, J.; van Dongen, J.T. Isolation and characterization of three new PGPR and their effects on the growth of Arabidopsis and Datura plants. J. Plant Interact. 2017, 12, 1–6. [Google Scholar] [CrossRef]
- Pascual, J.; García-López, M.; Bills, G.F.; Genilloud, O. Pseudomonas granadensis sp. nov., a new bacterial species isolated from the Tejeda, Almijara and Alhama Natural Park, Granada, Spain. Int. J. Syst. Evol. Microbiol. 2015, 65, 625–632. [Google Scholar] [CrossRef]
- Rana, S.; Jindal, V.; Mandal, K.; Kaur, G.; Gupta, V.K. Thiamethoxam degradation by Pseudomonas and Bacillus strains isolated from agricultural soils. Environ. Monit. Assess. 2015, 187, 300. [Google Scholar] [CrossRef]
- Kämpfer, P.; Thummes, K.; Chu, H.I.; Tan, C.C.; Arun, A.B.; Chen, W.M.; Lai, W.A.; Shen, F.T.; Rekha, P.D.; Young, C.C. Pseudacidovorax intermedius gen. nov., sp. nov., a novel nitrogen-fixing betaproteobacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2008, 58, 491–495. [Google Scholar] [CrossRef]
- Liaqat, F.; Eltem, R. Identification and characterization of endophytic bacteria isolated from in vitro cultures of peach and pear rootstocks. 3 Biotech 2016, 6, 120. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, J.; Liu, Y.; Chen, S. Isolation and identification of nitrogen-fixing bacilli from plant rhizospheres in Beijing region. J. Appl. Microbiol. 2005, 99, 1271–1281. [Google Scholar] [CrossRef] [Green Version]
- Nongkhlaw, F.M.; Joshi, S.R. Epiphytic and endophytic bacteria that promote growth of ethnomedicinal plants in the subtropical forests of Meghalaya, India. Rev. Biol. Trop. 2014, 62, 1295–1308. [Google Scholar] [CrossRef]
- Ambardar, S.; Vakhlu, J. Plant growth promoting bacteria from Crocus sativus rhizosphere. World J. Microbiol. Biotechnol. 2013, 29, 2271–2279. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A.; Hosseini, H.M. Indole-3-acetic acid (IAA) production trait, a useful screening to select endophytic and rhizosphere competent bacteria for rice growth promoting agents. MethodsX 2015, 2, 72–78. [Google Scholar] [CrossRef]
- García-Jaramillo, M.; Redondo-Gómez, S.; Barcia-Piedras, J.M.; Aguilar, M.; Jurado, V.; Hermosín, M.C.; Cox, L. Dissipation and effects of tricyclazole on soil microbial communities and rice growth as affected by amendment with alperujo compost. Sci. Total Environ. 2016, 550, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Moulas, C.; Petsoulas, C.; Rousidou, K.; Perruchon, C.; Karas, P.; Karpouzas, D.G. Effects of systemic pesticides imidacloprid and metalaxyl on the phyllosphere of pepper plants. Biomed. Res. Int. 2013, 2013, 969750. [Google Scholar] [CrossRef]
- Singh, N.; Gupta, S.; Marwa, N.; Pandey, V.; Verma, P.C.; Rathaur, S.; Singh, N. Arsenic mediated modifications in Bacillus aryabhattai and their biotechnological applications for arsenic bioremediation. Chemosphere 2016, 164, 524–534. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Liu, M.; Liu, X.; Cheng, B.S.; Ma, X.L.; Lyu, X.T.; Zhao, X.F.; Ju, Y.L.; Min, Z.; Fang, Y.L. Selection and evaluation of phosphate-solubilizing bacteria from grapevine rhizospheres for use as biofertilizers. Span. J. Agric. Res. 2016, 14, e1106. [Google Scholar] [CrossRef] [Green Version]
- Pailan, S.; Gupta, D.; Apte, S.; Krishnamurthi, S.; Saha, P. Degradation of organophosphate insecticide by a novel Bacillus aryabhattai strain SanPS1, isolated from soil of agricultural field in Burdwan, West Bengal, India. Int. Biodeterior. Biodegrad. 2015, 103, 191–195. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda Mdel, C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Min) | Eluent A (%) | Eluent B (%) |
|---|---|---|
| 0.1% acetic acid (v/v, aqueous) | 80:20 Acetonitrile: H2O (v/v) | |
| 0 | 80 | 20 |
| 13 | 50 | 50 |
| 18 | 0 | 100 |
| 22 | 80 | 20 |
| 26 | Stop |
| Treatment † | Strain | Closest Identificaton | Similarity (%) | Nitrogen Fixation | Phosphate Solubilization | IAA ‡ (mg/L) |
|---|---|---|---|---|---|---|
| 10× Etridiazole | E1 | Pseudacidovorax intermedius | 100 | + | - | <1 |
| 10× Etridiazole | E2 | Rhizobium larrymoorei | 99.78 | + | - | 4 |
| 10× Etridiazole | E4 | Sphingomonas aquatilis | 100 | + | + | <1 |
| 10× Etridiazole | E5 | Flavobacterium ginsengiterrae | 97.1 | - | - | <1 |
| 10× Etridiazole | E6 | Brevundimonas vesicularis | 99.63 | - | - | <1 |
| 10× Etridiazole | E7 | Bacillus aryabhattai | 100 | + | - | 7.9 |
| 10× Etridiazole | E9 | Mycobacterium sphagni | 99.42 | + | + | <1 |
| 10× Etridiazole | E10 | Mycobacterium vanbaalenii | 99.71 | + | - | <1 |
| 10× Etridiazole | EN1 | Mycobacterium aromaticivorans | 99.06 | + | + | <1 |
| 10× Metalaxyl | M1 | Bacillus sporothermodurans | 98.6 | + | - | <1 |
| 5× Metalaxyl | M7 | Bacillus fortis | 98.17 | - | - | <1 |
| 1× Metalaxyl | MN1 | Bacillus aryabhattai | 100 | + | - | 8.7 |
| 5× Tricyclazole | T6 | Pseudomonas granadensis | 99.49 | + | + | 2.0 |
| 5× Tricyclazole | T9 | Bacillus fortis | 98.38 | + | - | 1.1 |
| 5× Tricyclazole | TN1 | Pseudomonas baetica | 99.57 | + | + | <1 |
| 5× Tricyclazole | TN3 | Pseudomonas plecoglossicida | 99.01 | + | + | <1 |
| 1× Tricyclazole | T3 | Bacillus marisflavi | 99.72 | + | - | <1 |
| Bacterial Strain | Fungicide Inhibition | Inoculation Test | ||||
|---|---|---|---|---|---|---|
| Etridiazole 7 mg/Kg | Metalaxy 19 mg/Kg | Tricyclazole 160 mg/Kg | Fungicides Mixture | Shoots Growth Promoting | Roots Growth Promoting | |
| Rhizobium larrymoorei E2 | − | − | + | + | + | + |
| Bacillus aryabhattai E7 | − | − | + | N/A | = | + |
| Bacillus aryabhattai MN1 | − | − | − | − | = | + |
| Pseudomonas granadensis T6 | − | − | + | N/A | + | + |
| Bacillus fortis T9 | + | − | N/A | N/A | N/A | N/A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, F.-T.; Yen, J.-H.; Liao, C.-S.; Chen, W.-C.; Chao, Y.-T. Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics. Sustainability 2019, 11, 1133. https://doi.org/10.3390/su11041133
Shen F-T, Yen J-H, Liao C-S, Chen W-C, Chao Y-T. Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics. Sustainability. 2019; 11(4):1133. https://doi.org/10.3390/su11041133
Chicago/Turabian StyleShen, Fo-Ting, Jui-Hung Yen, Chien-Sen Liao, Wen-Ching Chen, and Yi-Ting Chao. 2019. "Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics" Sustainability 11, no. 4: 1133. https://doi.org/10.3390/su11041133
APA StyleShen, F. -T., Yen, J. -H., Liao, C. -S., Chen, W. -C., & Chao, Y. -T. (2019). Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics. Sustainability, 11(4), 1133. https://doi.org/10.3390/su11041133