Development of a Biogeochemical and Carbon Model Related to Ocean Acidification Indices with an Operational Ocean Model Product in the North Western Pacific


Abstract
:1. Introduction
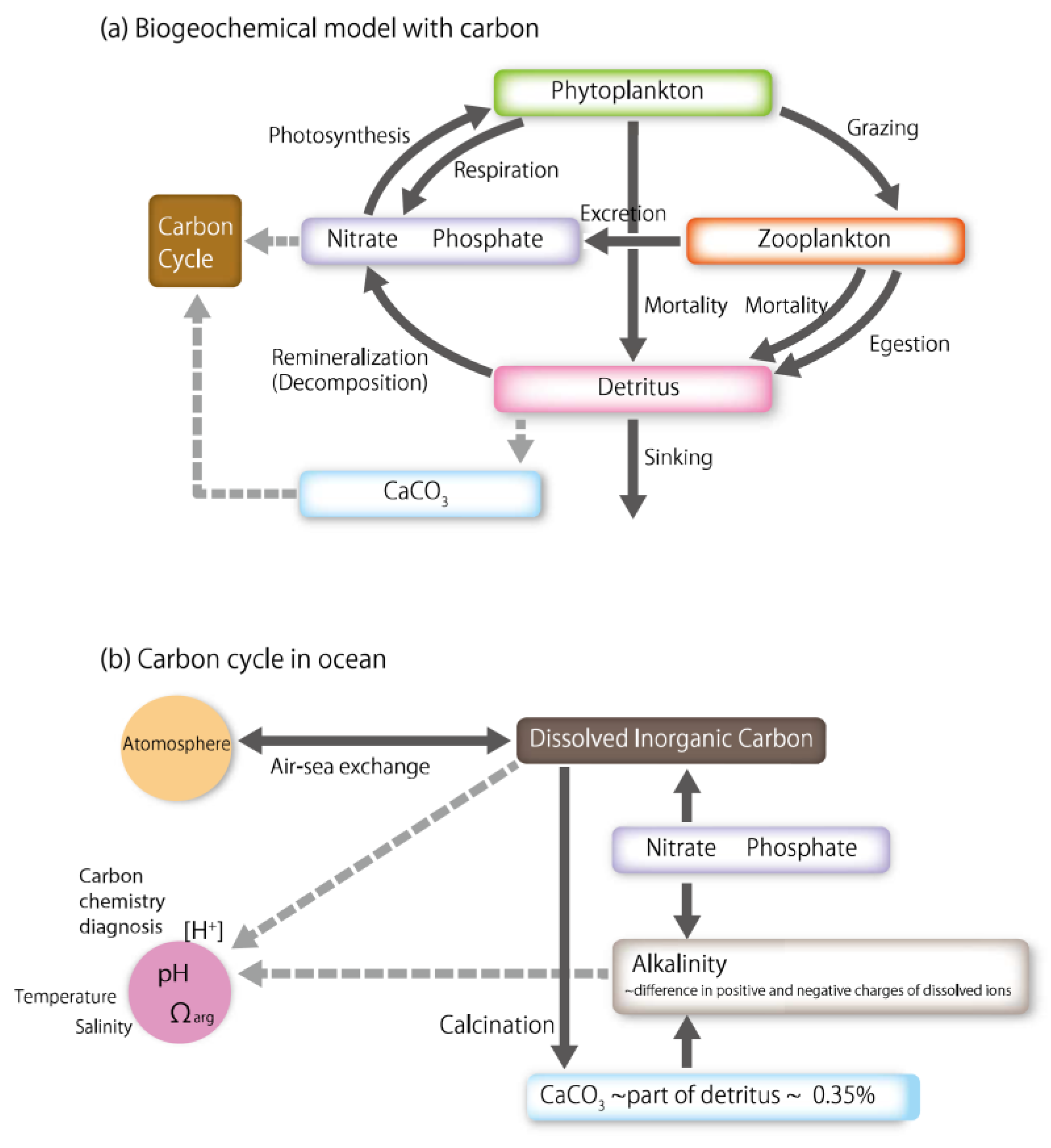
2. Model Formation
2.1. Equations Governing the Marine Ecosystem Model
2.2. Equations Governing the Carbon Cycle Model
2.3. Inorganic Carbon Cycling in Water
3. Data and Model Parameters
3.1. Climatology and Forcing Data
3.2. Biogeochemical Parameters and Observational Data for Validation
3.3. Initial and Boundary Conditions
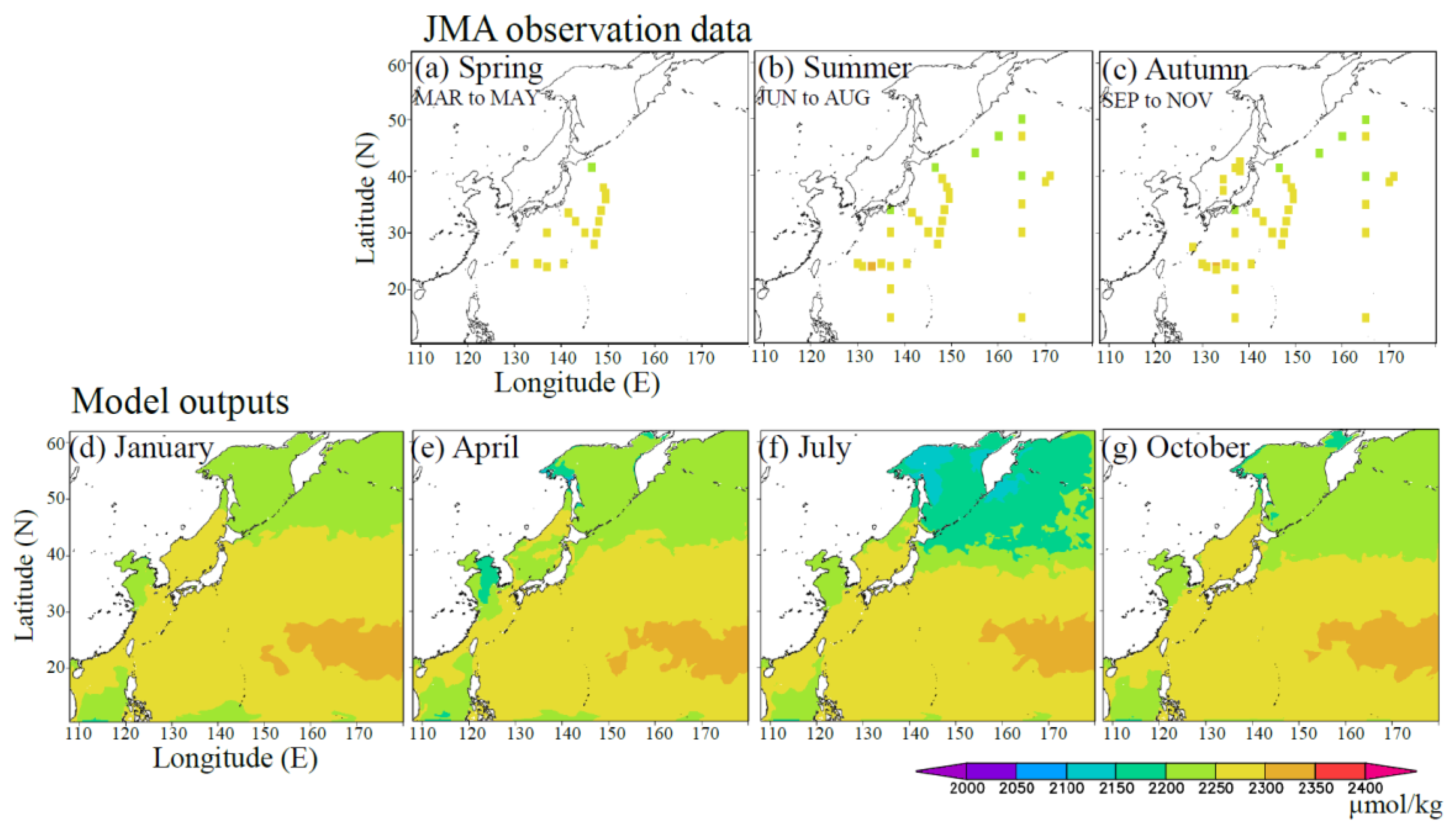
4. Results
4.1. Comparison of the Observed Data with the Simulated Outputs
4.2. Ocean Acidification Indices pH and Ωarg
4.3. Reproducibility of the Biogeochemical Variables
5. Discussion
5.1. Sensitivity Experiment for pH and Ωarg Based on Constant Temperature, Salinity, DIC and ALK
5.2. Effect of Seasonal Processes on pHinsitu
5.3. Seasonal Processes Affecting Ωarg Variations
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Baur, J.F.; Cai, W.J.; Raymond, P.A.; Bianchi, T.S.; Hopkinson, C.H.; Pierre, A.; Regnier, P. The changing carbon cycle of the coastal ocean. Nature 2013, 504, 61–70. [Google Scholar] [CrossRef]
- Bednarsek, N.; Feely, A.R.; Reum, J.C.P.; Peterson, B.; Menkel, J.; Alin, R.S.; Hales, B. Limacina helicina shell disoolution as an indicator of declining habitat suitability owing to ocean acidification in the California Current Ecosystem. Proc. Biol. Sci. 2014, 281, 20140123. [Google Scholar] [CrossRef] [PubMed]
- Feely, A.R.; Sabine, L.C.; Barne, H.R.; Millero, J.F.; Dickson, G.A.; Wanninkhof, R.; Murata, A.; Miller, A.L.; Greeley, D. Decadal changes in the aragonite and calcite saturation state of the Pacific Ocean. Glob. Biogeochem. Cycles 2012, 26, GB3001. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Zia, Y.B.; Midgley, P.M. (Eds.) Intergovernmental Panel on Climate Change (IPCC), Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 1–1535. [Google Scholar]
- Kroeker, J.K.; Kordas, L.R.; Crim, R.; Hendrik, E.I.; Ramajo, L.; Singh, S.G.; Duarte, M.C.; Gattuso, P.J. Impacts of ocean acidification on marine organisms: Qualitifying sensitivities and interaction with warming. Chang. Biol. 2013, 19, 1884–1896. [Google Scholar] [CrossRef] [PubMed]
- Feely, A.R.; Alin, R.S.; Carter, B.; Bednarsek, N.; Hales, B.; Chan, F.; Hill, M.T.; Gaylord, B.; Sanford, E.; Byrne, H.R.; et al. Chemical and biological impacts of ocean acidification along the west coast of North America. Estuar. Coast. Shelf Sci. 2016, 183, 260–270. [Google Scholar] [CrossRef]
- Wittmann, C.A.; Portner, H. Sensitivities of extant animal taxa to ocean acidification. Nat. Clim. Chang. 2013, 3, 995–1001. [Google Scholar] [CrossRef]
- IGBP; IOC; SCOR. Ocean Acidification Summary for Policymakers. In Proceedings of the Third symposium on the Ocean in a High-CO2 World International Geoshpere-Bioshere Programme, Stockholm, Sweden, 14 November 2013. [Google Scholar]
- Bender, M.; Doney, S.; Feely, R.A.; Fung, I.; Gruber, N.; Harrison, D.E.; Keeling, R.; Moore, J.K.; Sarmiento, J.; Sarachik, E.; et al. A Large–Scale CO2 Observation Plan: In Situ Oceans and Atmosphere (LSCOP); NOAA OAR Special Report; National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 2002; pp. 1–201.
- Johnson, S.K.; Plant, N.J.; Coletti, J.L.; Jannasch, W.W.; Sakamoto, M.C.; Rise, C.S.; Swift, D.D.; Williams, L.N.; Boss, E.; Haentjens, N.; et al. Biogeochemical sensor performance in the SOCCOM profiling float array. J. Geophys. Res. Ocean 2017, 122, 6416–6436. [Google Scholar] [CrossRef]
- Doney, S.C.; Lima, I.; Feely, R.A.; Glover, D.M.; Lindsay, K.; Mahowald, N.; Moore, J.K.; Wanninkhof, R. Mechanisms governing interannual variability in upper-ocean inorganic carbon system and air-sea CO2 fluxes: Physical climate and atmosphere dust. Deep-Sea Res. II 2009, 56, 640–655. [Google Scholar] [CrossRef]
- Metz, N.; Tilbrook, B.; Bakker, D.; Quere, L.; Doney, C.; Feely, R.; Hood, M.; Dargaville, R. Global changes in ocean carbon: Variability and vulnerability. EOS Trans. 2007, 88, 287. [Google Scholar]
- Le Quere, C.; Orr, J.C.; Monfray, P.; Aumont, O. Interannual variability of the oceanic sink of CO2 from 1979 through 1997. Glob. Biogeochem. Cycles 2000, 14, 1247–1265. [Google Scholar] [CrossRef]
- McKinley, G.A.; Follows, M.J.; Marshall, J. Mechanisms of air–Sea CO2 flux variability in the equatorial Pacific and the North Atlantic. Glob. Biogeochem. Cycles 2004, 18, GB2011. [Google Scholar] [CrossRef]
- McKinley, G.A.; Takahashi, T.; Butenhuis, E.; Chai, F.; Christian, J.R.; Doney, S.C.; Jiang, M.S.; Lindsay, K.; Moore, J.K.; LeQuere, C.; et al. North Pacific carbon cycle response to climate variability on seasonal to decadal timescales. J. Geophys. Res. Oceans 2006, 111, C07S06. [Google Scholar] [CrossRef]
- Obata, A.; Kitamura, Y. Interannual variability of the sea-air exchange of CO2 from 1961 to 1998 simulated with a global ocean circulation-biogeochemistry model. J. Geophys. Res. 2003, 108, 3337. [Google Scholar] [CrossRef]
- Wetzel, P.; Winguth, A.; Maier-Reimer, E. Sea–to–air CO2 flux from 1948 to 2003: A model study. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Schmittner, A.; Oschiles, A.; Matthews, H.D.; Galbraith, E.D. Future changes in climate, ocean circulation, ecosystems, and biogeochemical cycling simulated for a business-as-usual CO2 emission scenario until year 4000 AD. Glob. Biogeochem. Cycles 2008, 22, GB1013. [Google Scholar] [CrossRef]
- Kawamiya, M.; Kishi, M.; Yamanaka, Y.; Suginohara, N. An ecological-physical coupled model applied to station Papa. J. Oceanogr. 1995, 51, 635–664. [Google Scholar] [CrossRef]
- Kawamiya, M.; Kishi, M.J.; Yamanaka, Y.; Suginohara, N. Obtaining reasonable results in different oceanic regimes with the same ecological-physical coupled model. J. Oceanogr. 1997, 53, 397–402. [Google Scholar]
- Kawamiya, M.; Kishi, M.; Suginohara, N. An ecosystem model for the North Pacific embedded in a general circulation model Part I: Model description and characteristics of spatial distributions of biological variables. J. Mar. Syst. 2000, 25, 129–157. [Google Scholar] [CrossRef]
- Kishi, M.J.; Kashiwai, M.; Ware, D.M.; Megrey, B.A.; Eslinger, D.L.; Werner, F.E.; Noguchi-Aita, M.; Azumaya, T.; Fujii, M.; Hashimot, S.; et al. NEMURO—A lower trophic level model for the North Pacific marine ecosystem. Ecol. Model. 2007, 202, 12–25. [Google Scholar] [CrossRef]
- Fujii, M.; Yamanak, Y.; Nojiri, Y.; Kishi, J.M.; Chai, F. Comparison of seasonal characteristics in biogeochemistry among the subarctic North Pacific stations described with a NEMURO-based marine ecosystem model. Ecol. Model. 2007, 202, 52–67. [Google Scholar] [CrossRef]
- Yoshie, N.; Yamanaka, Y.; Rose, K.A.; Eslinger, D.L.; Ware, D.M.; Kishi, M.J. Parameter sensitivity study of the NEMURO lower trophic level marine ecosystem model. Ecol. Model. 2007, 202, 26–37. [Google Scholar] [CrossRef]
- Sasai, Y.; Yoshikawa, C.; Smith, L.S.; Hashioka, T.; Matsumoto, K.; Wakita, M.; Sasaoka, K.; Honda, C.M. Coupled 1-D physical-biological model study of phytoplankton production at two contrasting time-series stations in the western North Pacific. J. Oceanogr. 2016, 72, 509–526. [Google Scholar] [CrossRef]
- Onitsuka, G.; Yanagi, T. Difference in ecosystem dynamics between the northern and southern parts of the Japan Sea: Analyses with two ecosystem models. J. Oceanogr. 2005, 61, 415–433. [Google Scholar] [CrossRef]
- Guo, X.; Yangai, T. The role of the Taiwan Strait in an ecological model in the east china sea. Acta Oceanogr. Taiwanica 1998, 37, 139–162. [Google Scholar]
- Zhao, L.; Gao, X. Influence of cross-shelf water transport on nutrients and phytoplankton in the East China Sea: A model study. Ocean Sci. 2011, 7, 27–43. [Google Scholar] [CrossRef]
- Luo, X.; Wei, H.; Liu, Z.; Zhao, L. Seasonal variability of air-sea CO2 fluxes in the Yellow and East China Seas: A case study of continental shelf sea carbon cycle model. Cont. Shelf Res. 2015, 107, 69–78. [Google Scholar] [CrossRef]
- Bopp, L.; Resplandy, L.; Doney, S.C.; Dunner, J.P.; Gehlen, M.; Halloran, P.; Heinze, C.; Ilyinas, T.; Seferian, R.; Tjiputra, J.; et al. Multiple stressors of ocean ecosystems in the 21th century: Projections with CMIP5 models. Biogeosciences 2013, 10, 6225–6245. [Google Scholar] [CrossRef]
- Moss, R.H.; Nakicenovic, N.; O’Neill, B.C. Towards New Scenarios for Analysis of Emissions, Climate Change, Impacts, and Response Strategies. In Proceedings of the Intergovernmental Panel on Climate Change, Geneva, Switzerland, 2 September 2008; p. 132. [Google Scholar]
- Taylor, E.K.; Stouffer, J.R.; Meehl, A.G. A Summary of the CMIP5 Experiment Design. Available online: https://pcmdi.llnl.gov/mips/cmip5/experiment_design.html (accessed on 1 February 2019).
- Taylor, E.K.; Stouffer, J.; Meehl, A.G. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef]
- Watanabe, S.; Hajime, T.; Sudo, K.; Nagashima, T.; Takemura, T.; Okajima, H.; Nozawa, T.; Kawase, H.; Abe, M.; Yokohata, T.; et al. MIROC-ESM 2010: Model description and basic results of CMIP5-20c3m experiments. Geosci. Model. Dev. 2011, 4, 845–872. [Google Scholar] [CrossRef]
- Miyazawa, Y.; Zhang, R.; Guo, X.; Tamura, H.; Ambe, D.; Lee, J.S.; Yoshinari, H.; Setou, T. Water mass variability in the western north Pacific detected in a 15-year eddy resolving ocean reanalysis. J. Oceanogr. 2009, 65, 737–756. [Google Scholar] [CrossRef]
- Miyazawa, Y.; Yamashita, N.; Taniyasu, S.; Yamazaki, E.; Guo, X.; Varlamov, M.S.; Miyama, T. Ocean dispersion simulation of perfluoroalkyl substances in the Western North Pacific associated with the Great East Japan Earthquake of 2011. J. Oceanogr. 2014, 70, 535–547. [Google Scholar] [CrossRef]
- Ishizu, M.; Miyazawa, Y.; Tsunoda, T.; Guo, X. Development of a marine carbon model coupled with an operational ocean model product for ocean acidification studies. Kaiyo Mon. 2018, 50, 217–225. [Google Scholar]
- Miyazawa, Y.; Varlamov, M.S.; Miyama, T.; Guo, X.; Hihara, T.; Kiyomatsu, K.; Kachi, M.; Kurihara, Y.; Murakami, H. Assimilation of high-resolution sea surface temperature data into an operational nowcast/forecast system around Japan using a multi-scale three-dimensional variational scheme. Ocean Dyn. 2017, 67, 713–728. [Google Scholar] [CrossRef]
- Heuven, V.S.; Pierrot, D.; Rae, J.W.B.; Lewis, E.; Wallance, D.W.R. Matlab program developed for CO2 system calculation, ORNL/CDIAC-105b. In Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory; U.S. Department of Energy: Oak Ridge, TN, USA, 2011. [Google Scholar] [CrossRef]
- Lewis, E.; Wallace, D.W.R. Program Developed for CO2 System Calculations. ORNL/CDIAC-105. In Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory; U.S. Department of Energy: Oak Ridge, TN, USA, 1998. [Google Scholar]
- Orr, J.C.; Najjar, R.; Sabine, C.L.; Joos, F. Abiodic-HOWTO; Internal OCMIP Report; LSCE/CEA Saclay: Gif-sur-Yvette, France, 1999; 25p. [Google Scholar]
- Ewen, T.L.; Weaver, A.J.; Eby, M. Sensitivity of the inorganic ocean carbon cycle to future climate warming in the UVic coupled model. Atmos. Ocean 2004, 42, 23–42. [Google Scholar] [CrossRef]
- Smith, L.S.; Yamanaka, Y.; Pahlow, M.; Oschlies, A. Optimal uptake kinetics: Physiological acclimation explains the pattern of nitrate uptake by phytoplankton in the ocean. Mar. Ecol. Prog. Ser. 2009, 384, 1–12. [Google Scholar] [CrossRef]
- Ishizu, M.; Richards, J.K. Relationship between oxygen, nitrate and phosphate in the world ocean based on potential temperature. J. Geophys. Res. 2013, 118, 3586–3594. [Google Scholar] [CrossRef]
- Redfield, A.C.; Ketchum, B.H.; Richards, A.F. The influence of organisusm on the Composition of Sea–Water. In The Sea; Hill, M.N., Ed.; Interscience: New York, NY, USA, 1963; Volume 2, pp. 26–27. [Google Scholar]
- Takatani, Y.; Enyo, K.; Iida, Y.; Kojima, A.; Sasano, D.; Kosugi, N.; Midorikawa, T.; Suzuki, T.; Ishii, M. Relationships between total alkalinity in surface water and seas surface dynamic height in the Pacific Ocean. J. Geophys. Res. Oceans 2014, 119, 2806–2814. [Google Scholar] [CrossRef]
- Kantha, L.H. A general ecosystem model for applications to primary productivity and carbon cycle studies in the global oceans. Ocean Model. 2004, 6, 285–334. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Yoshie, N.; Fujii, M.; Aita, N.M.; Kishi, M. An ecosystem model coupled with nitrogen-silicon-carbon cycles applied to Station A7 in the Northwestern Pacific. J. Oceanogr. 2004, 60, 227–241. [Google Scholar] [CrossRef]
- Mussi, A. The solubility of calcite and argonate in seawater at various salinities, temperature and one atmosphere total pressure. Am. J. Sci. 1983, 283, 780–799. [Google Scholar]
- Zeebe, R.E.; Wolf-Gladrow, D. CO2 in Seawater: Equilibrium, Kinetics, Isotopes, 1st ed.; Elsevier: New York, NY, USA, 2001; Volume 65, p. 360. [Google Scholar]
- Mackenzie, F.T.; Lerman, A.; Anderson, A.J. Past and present of sediment and carbon biogeochemical cycling models. Biogeosciences 2004, 1, 11–32. [Google Scholar] [CrossRef]
- Brocker, W.S.; Peng, T.H. The role of CaCO3 compensation in the glacial to interglacial atmospheric CO2 change. Glob. Biogeochem. Cycles 1987, 1, 5–29. [Google Scholar]
- Wanninkhof, R. Relationship between wind speed and gas exchange over the ocean. J. Geophys. Res. Oceans 1992, 97, 7373–7382. [Google Scholar] [CrossRef]
- Weiss, R. The Provisions of Social Relationships. In Doing unto Others; Rubin, Z., Ed.; Prentice Hall: Englewood Cliffs, UK, 1974; pp. 17–26. [Google Scholar]
- Yasunaka, S.; Nojiri, Y.; Nakaoka, S.; Ono, T.; Whitney, F.; Telszewski, M. Mapping of sea surface nutrients in the North Pacific: Basinwide distribution and seasonal to interannual variability. J. Geophys. Res. Oceans 2014, 119, 7756–7771. [Google Scholar] [CrossRef]
- Goyet, C.; Healy, R.; Ryan, J. Global distribution of total inorganic carbon and total alkalinity below the deepest winter mixed layer depths, ORNL/CDIAC-127, NDP-076. In Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory; U.S. Department of Energy: Oak Ridge, TN, USA, 2000; pp. 1–40. [Google Scholar] [CrossRef]
- Key, M.R.; Kozyer, A.; Sabine, L.C.; Lee, K.; Wanninkhof, R.; Bullister, L.J.; Feely, A.R.; Millero, J.F.; Mordy, C.; Peng, T.H. A global ocean carbon climatology: Results from global data analysis project (GLODAP). Glob. Biogeochem. Cycles 2004, 18, GB4031. [Google Scholar] [CrossRef]
- Yasunaka, S.; Nojiri, Y.; Nakaoka, S.; Ono, T.; Mukai, H.; Usui, N. Monthly maps of sea surface dissolved inorganic carbon in the North Pacific: Basin-wide distribution and seasonal variation. J. Geophys. Res. Oceans 2013, 118, 3843–3850. [Google Scholar] [CrossRef]
- Takahashi, T.; Sutherland, C.S.; Chipman, W.D.; Goddard, G.J.; Cheng, H.; Newberger, T.; Sweeney, C.; Munro, R.D. Climatological distributions of pH, pCO2, total CO2, alkalinity, and CaCO3 saturation in the global surface ocean, and temporal changes at selected locations. Mar. Chem. 2014, 164, 95–125. [Google Scholar] [CrossRef]
- Yoshie, N.; Guo, X.; Fujii, N.; Komorita, T. Ecosystem and nutrient dynamics in the Seto Inland Sea, Japan, Interdisciplinary Studies on environmental Chemistry—Marine Environmental modeling and analysis. Model. Anal. Mar. Environ. Probl. 2011, 5, 39–49. [Google Scholar]
- Edward, F.K.; Thomas, K.M.; Klausmeier, A.C.; Litchman, E. Light and growth in marine phytoplankton: Allometric, taxonomic, and environmental variation. Limnol. Oceanogr. 2015, 60, 540–552. [Google Scholar] [CrossRef]
- Lalli, M.C.; Parsons, R.T. Biological Oceanography: An Introduction, 2nd ed.; Butterworth-Heinemann: Oxford, UK, 1997; 320p. [Google Scholar]
- Nelson, B.N.; Sigel, A.D. The global distribution and dynamics of chromophoric dissolved organic matter. Annu. Rev. Mar. Sci. 2013, 5, 447–476. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.Q.; Feely, A.R.; Carter, R.B.; Greeley, J.D.; Gledhill, K.D.; Arzayus, M.K. Climatological distribution of aragonite saturation state in the global oceans. Biogeochem. Cycles 2015, 29, 1656–1673. [Google Scholar] [CrossRef]
- Yara, Y.; Vogt, M.; Fujii, M.; Yamano, H.; Hauri, C.; Steinacher, M.; Gruber, N.; Yamanaka, Y. Ocean acidification limits temperature-induced poleward expansion of coral habitats around Japan. Biogeosciences 2012, 9, 4955–4968. [Google Scholar] [CrossRef]
- Campbell, W.J. The lognormal distribution as a model for bio-optical variability in the sea. J. Geophys. Res. 1995, 100, 13237–13254. [Google Scholar] [CrossRef]
- Tsuda, A.; Takeda, S.; Saito, H.; Nishioka, J.; Nojiri, Y.; Kudo, I.; Kiyosawa, H.; Imai, I.; Ono, T.; Shimamoto, A.; et al. A mesoscale iron enrichment in the western subarctic Pacific induces large centric diatom bloom. Science 2003, 300, 958–961. [Google Scholar] [CrossRef] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Definition | Value | Units | Reference Values Onitsuka and Yanagi [26] |
|---|---|---|---|---|
| Ecosystem Model | ||||
| For phytoplankton | ||||
| Vmax | Growth rate for phytoplankton | 0.0492 | day−1 | 0.6 |
| A | Affinity coefficient of basic cellular physiology | 6.75 | mmolN−1m−1day−1 | − |
| KDIN | Half saturation constant for DIN | 1.50 | mmolN m−1 | 1.5 |
| KDIP | Half saturation constant for DIP | 0.0940 | mmolN m−1 | 0.09 |
| MP | Phytoplankton mortality rate at 0 °C | 0.0400 | (mmolNm−1) m−3 day−1 | 0.07 |
| Pmin | Threshold of phytoplankton mortality | 0.0587 | (mmolNm−1) m−3 | − |
| R | Phytoplankton respiration rate at 0 °C | 0.0317 | day−1 | 0.3 |
| CPT | Temperature coefficient for photosynthesis | 0.0392 | °C−1 | 0.0693 |
| CRPT | Temperature coefficient for phytoplankton respiration | 0.0519 | °C−1 | 0.0519 |
| CMPT | Temperature coefficient for phytoplankton mortality | 0.0693 | °C−1 | 0.0693 |
| Iopt | Optimum light intensity for phytoplankton | * 20–120 | W m−2 | 70 |
| Iopt0 | 100.0 | − | ||
| Iopt1 | 70.0 | − | ||
| latbnd | Boundary for latitudinal differences | 45.0 | − | |
| latslp | Slope for latitudinal differences | 4.0 | − | |
| cdom | Light dissipation coefficient of seawater | * 0.015–0.045 | m−1 | 0.04 |
| cdom0 | 0.0275 | m−1 | − | |
| cdom1 | 0.0250 | m−1 | − | |
| Latslp_dom | Slope for latitudinal differences for cdom | 1.5 | − | |
| For zooplankton | ||||
| GZ | Maximum grazing rate of zooplankton at 0 °C | 0.05 | day−1 | 0.3 |
| Λ | Ivlev constant | 1.4 | (mmolN m−3)−1 | 1.4 |
| MZ | Zooplankton mortality rate at 0 °C | 0.05 | °C−1 | 0.07 |
| βz | Growth efficiency of zooplankton | 0.3 | 0.3 | |
| αz | Assimilation efficiency of zooplankton | 0.7 | 0.7 | |
| P* | Zooplankton threshold value for grazing on phytoplankton | 0.0430 | (mmolNm−1) m−3 | 0.043 |
| CGZT | Temperature coefficient for zooplankton grazing | 0.0390 | °C−1 | 0.0693 |
| CMPT | Temperature coefficient for zooplankton mortality | 0.0693 | °C−1 | 0.0693 |
| For diatoms | ||||
| WD | Singing velocity of detritus | 6.7 | m day−1 | 10 |
| VPN | Decomposition rate at 0 °C (DET→DIN) | 0.050 | day−1 | 0.05 |
| CλDT | Temperature coefficient for decomposition | 0.0693 | °C−1 | 0.0693 |
| Carbon Cycle Model | ||||
| RP:N | Stoichiometry of nitrogen to phosphorus [46] | 16.0 | − | |
| RC:P | Molar elemental ratios | 112.0 | − | |
| RCaCO3/POC | CaCO3 over nonphotosynthetical POC production ratio | 0.035 | − | |
| RALK:N | Alkalinity over non-photosynthetic N production ratio | 0.001 | − | |
| DCaCO3 | CaCO3 remineralization e-folding depth | 3500.0 | m | − |
| Chlorophyll-a | ||
|---|---|---|
| Subarctic Region | Subtropical Region | |
| Vmax | 0.692 | 0.692 |
| R | 0.02 | 0.04 |
| MP | 0.045 | 0.3 |
| GZ | 0.05 | 0.05 |
| MZ | 0.05 | 0.05 |
| VPN | 0.05 | 0.05 |
| WD | 6.79 | 6.78 |
| Parameter | Subtropical Region | Subarctic Region | Kuroshio Extension | Japan Sea |
|---|---|---|---|---|
| Chl-a | 0.57 (0.07) | 0.88 (0.07) | 0.86 (0.12) | 0.87 (0.15) |
| DIN | 0.72 (0.04) | 0.88 (0.12) | 0.87 (0.10) | 0.95 (0.11) |
| DIC | 0.74 (0.09) | 0.87 (0.20) | 0.78 (0.17) | 0.92 (0.26) |
| ALK | 0.48 (0.09) | 0.69 (0.20) | 0.48 (0.17) | 0.35 (0.26) |
| pH25 | 0.78 (0.09) | 0.86 (0.20) | 0.81 (0.17) | 0.90 (0.26) |
| Ωarg | 0.99 (0.09) | 0.98 (0.20) | 0.98 (0.17) | 0.99 (0.26) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishizu, M.; Miyazawa, Y.; Tsunoda, T.; Guo, X. Development of a Biogeochemical and Carbon Model Related to Ocean Acidification Indices with an Operational Ocean Model Product in the North Western Pacific. Sustainability 2019, 11, 2677. https://doi.org/10.3390/su11092677
Ishizu M, Miyazawa Y, Tsunoda T, Guo X. Development of a Biogeochemical and Carbon Model Related to Ocean Acidification Indices with an Operational Ocean Model Product in the North Western Pacific. Sustainability. 2019; 11(9):2677. https://doi.org/10.3390/su11092677
Chicago/Turabian StyleIshizu, Miho, Yasumasa Miyazawa, Tomohiko Tsunoda, and Xinyu Guo. 2019. "Development of a Biogeochemical and Carbon Model Related to Ocean Acidification Indices with an Operational Ocean Model Product in the North Western Pacific" Sustainability 11, no. 9: 2677. https://doi.org/10.3390/su11092677
APA StyleIshizu, M., Miyazawa, Y., Tsunoda, T., & Guo, X. (2019). Development of a Biogeochemical and Carbon Model Related to Ocean Acidification Indices with an Operational Ocean Model Product in the North Western Pacific. Sustainability, 11(9), 2677. https://doi.org/10.3390/su11092677