Successional Variation in the Soil Microbial Community in Odaesan National Park, Korea


 , and
, and 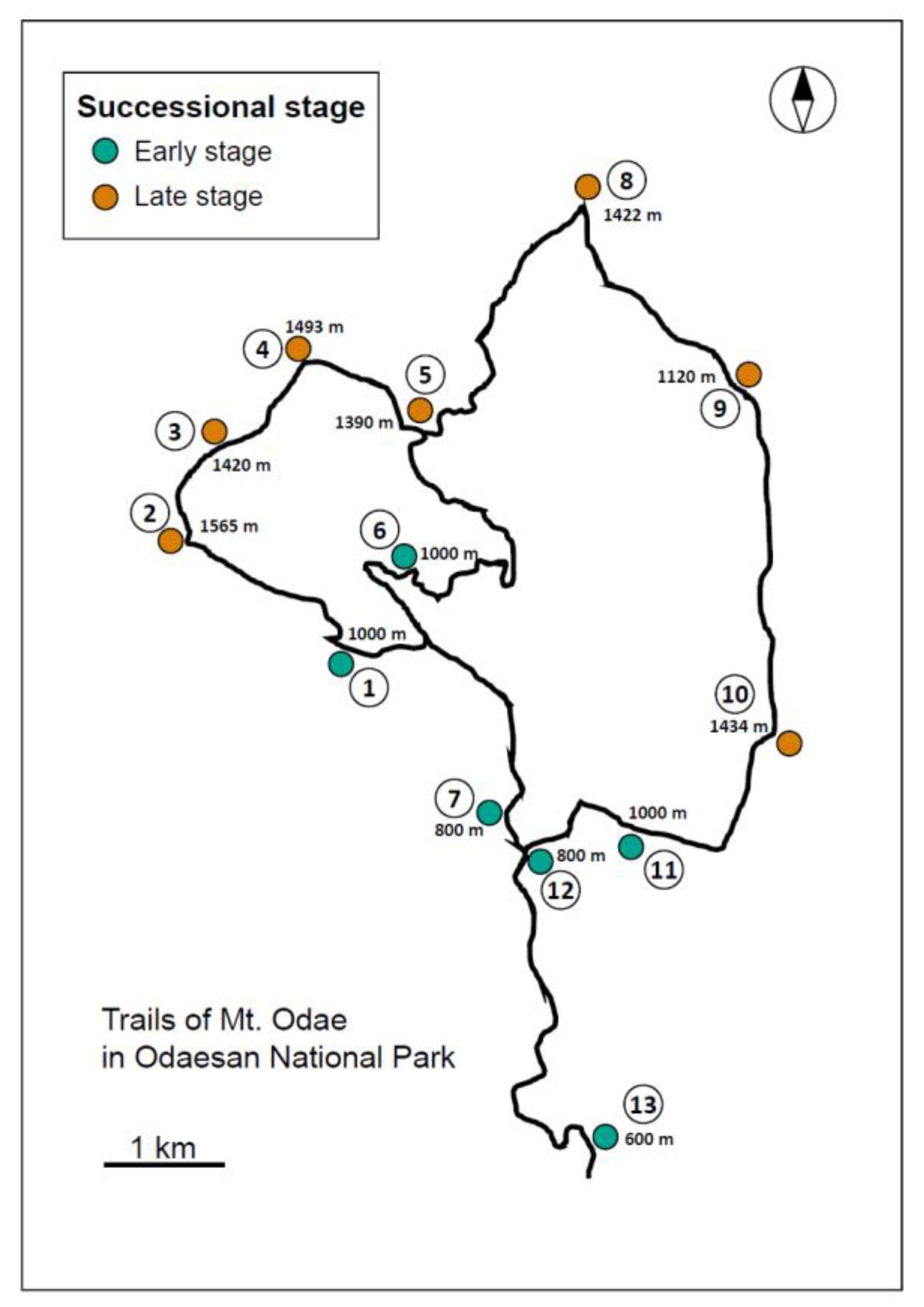{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Study Sites
2.2. Soil Sampling
2.3. Analysis of Soil Properties
2.4. DNA Library Preparation and 454 Pyrosequencing
2.5. Bioinformatics Analysis
3. Results
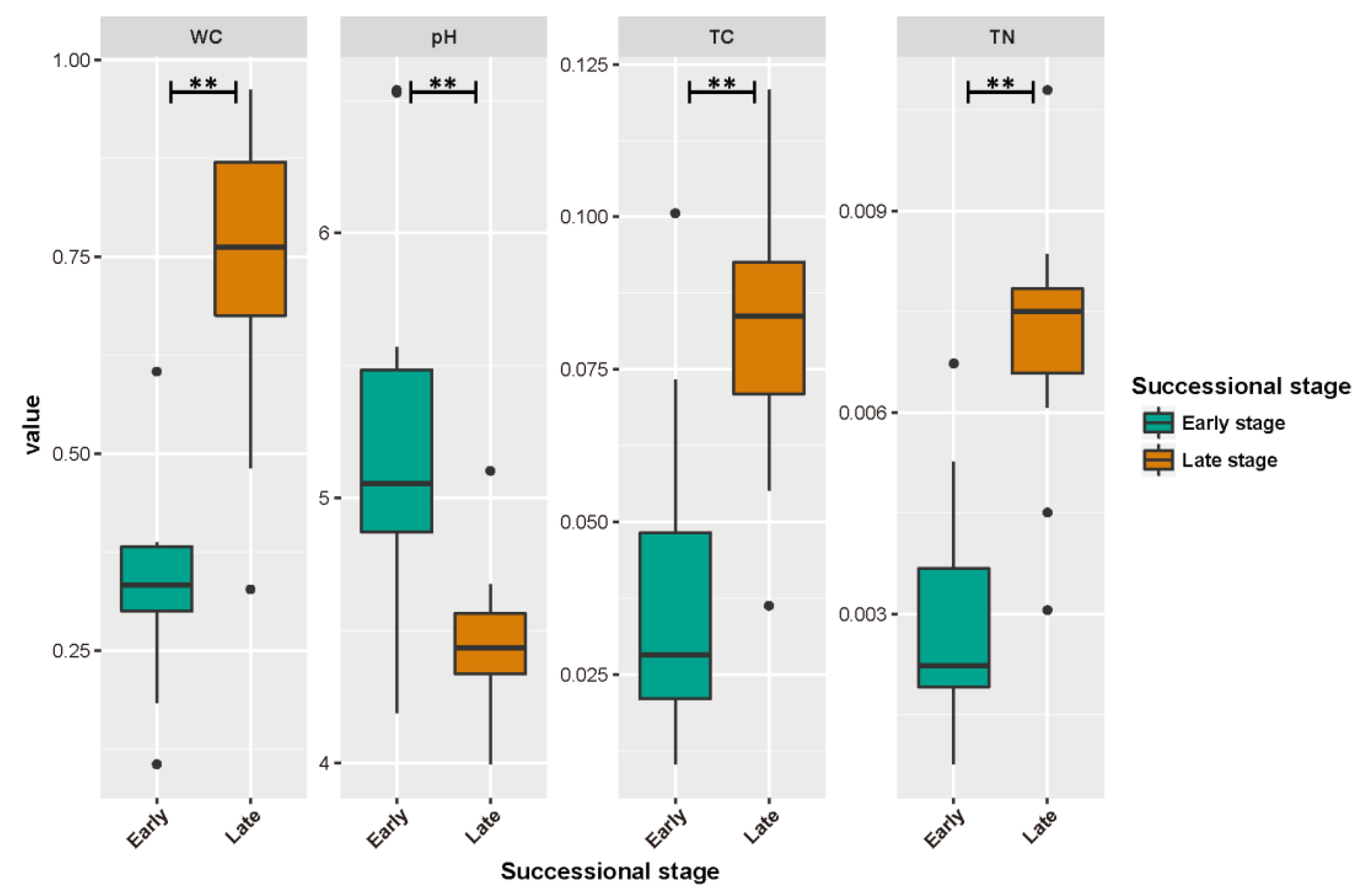
3.1. Soil Properties
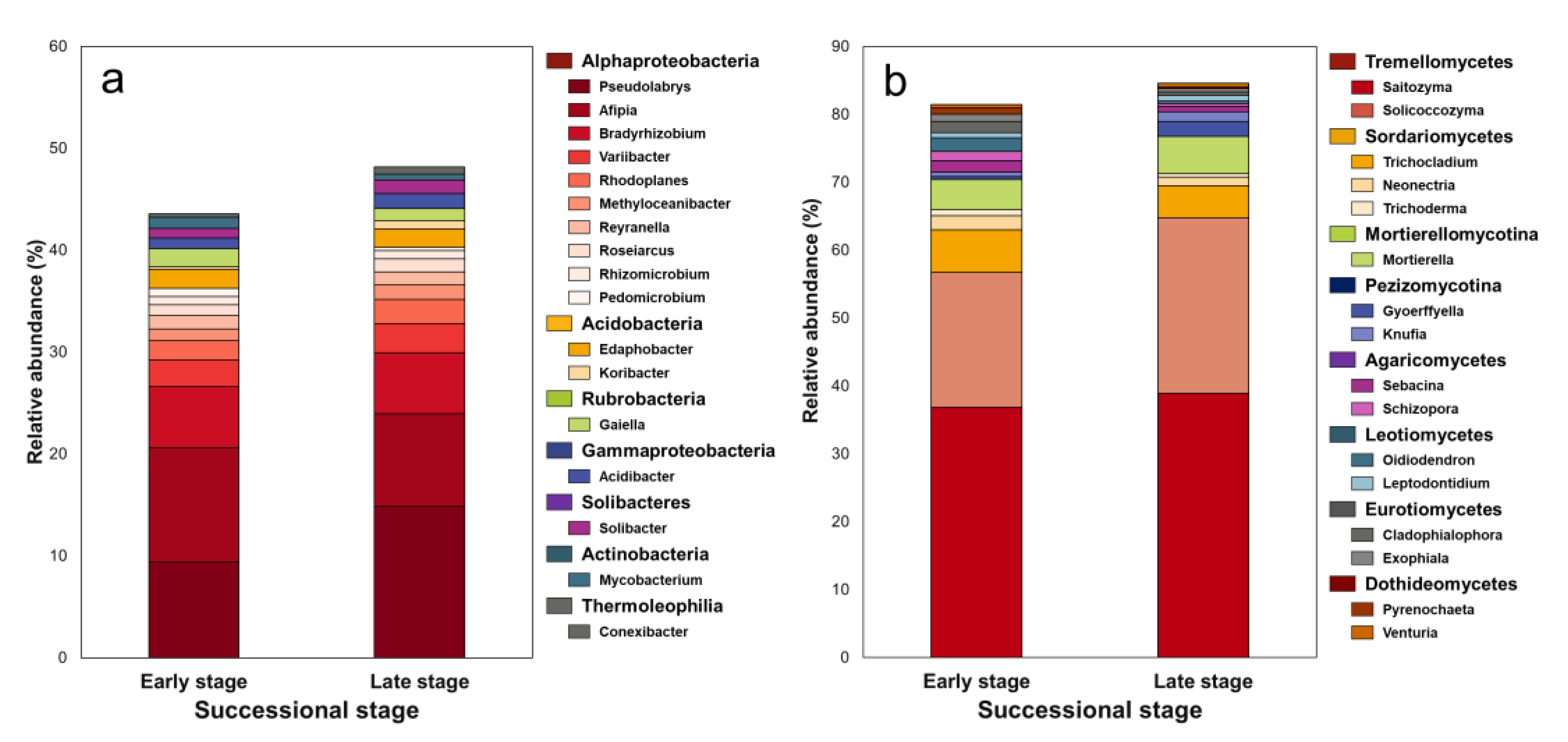
3.2. Microbial Composition
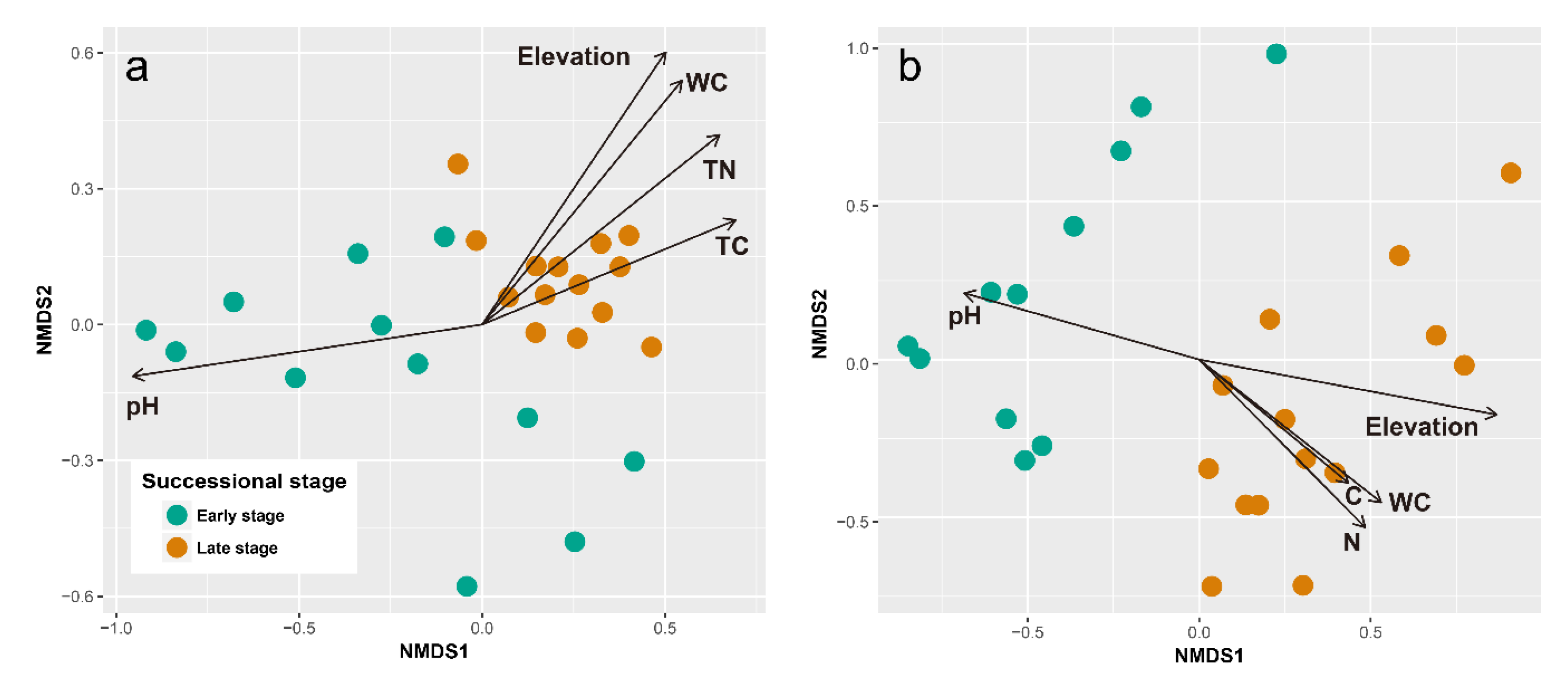
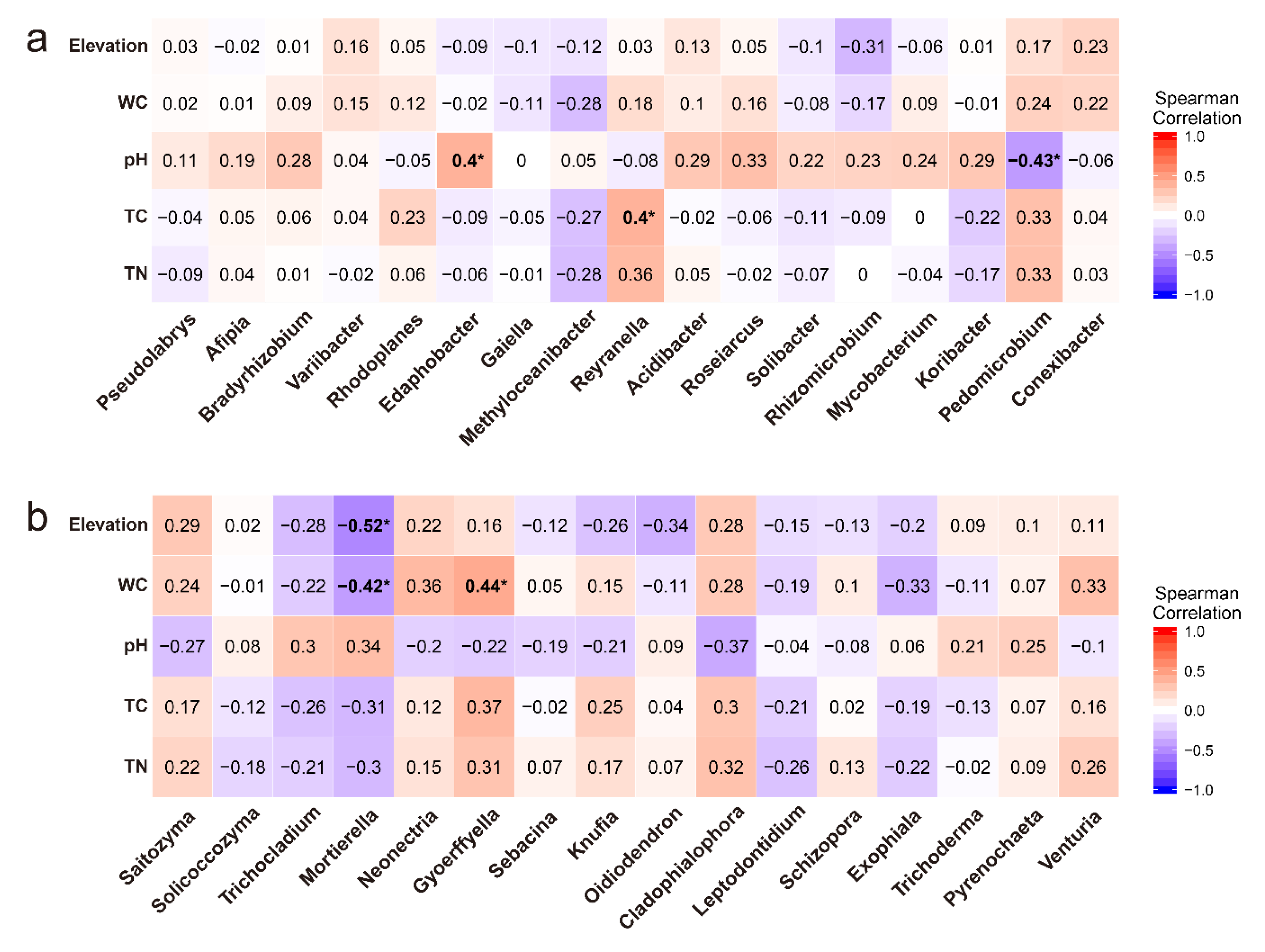
3.3. Microbial Diversity and Its Relationships with Environmental Variables
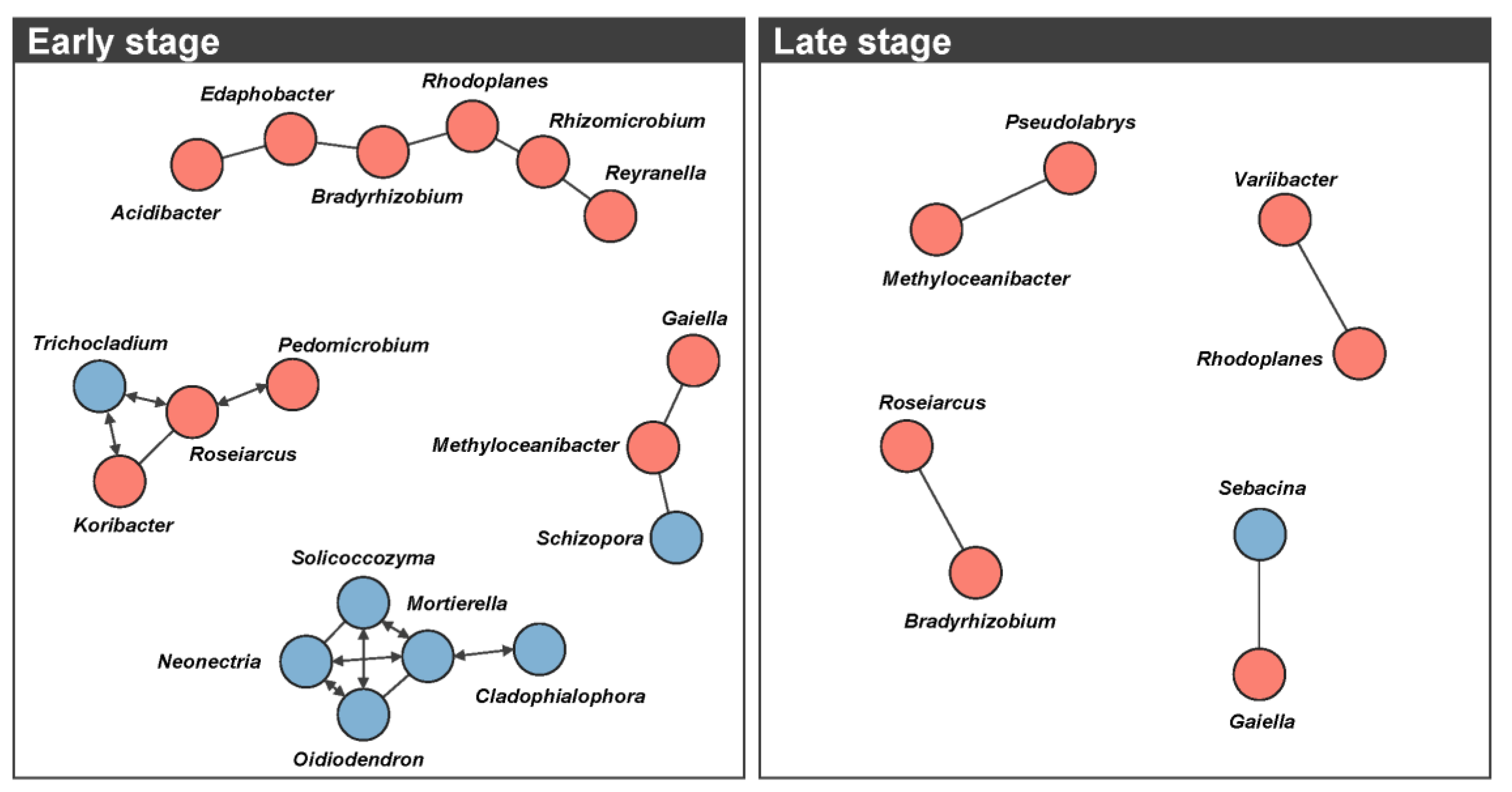
3.4. Microbial Networks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wooley, J.C.; Ye, Y.Z. Metagenomics: Facts and artifacts, and computational challenges. J. Comput. Sci. Technol. 2010, 25, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Lee, D.W.; Kwon, S.L.; Heo, Y.M.; Jang, S.; Kwon, B.-O.; Khim, J.S.; Kim, G.-H.; Kim, J.-J. Importance of functional diversity in assessing the recovery of the microbial community after the Hebei Spirit oil spill in Korea. Environ. Int. 2019, 128, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Igalavithana, A.D.; Lee, S.S.; Niazi, N.K.; Lee, Y.-H.; Kim, K.H.; Park, J.-H.; Moon, D.H.; Ok, Y.S. Assessment of soil health in urban agriculture: Soil enzymes and microbial properties. Sustainability Basel 2017, 9, 310. [Google Scholar] [CrossRef] [Green Version]
- Birgander, J.; Rousk, J.; Olsson, P.A. Comparison of fertility and seasonal effects on grassland microbial communities. Soil Biol. Biochem. 2014, 76, 80–89. [Google Scholar] [CrossRef]
- Heo, Y.M.; Lee, H.; Kim, K.; Kwon, S.L.; Park, M.Y.; Kang, J.E.; Kim, G.-H.; Kim, B.S.; Kim, J.-J. Fungal diversity in intertidal mudflats and abandoned solar salterns as a source for biological resources. Mar. Drugs 2019, 17, 601. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Rui, J.P.; Mao, Y.J.; Yannarell, A.; Mackie, R. Dynamics of the bacterial community structure in the rhizosphere of a maize cultivar. Soil Biol. Biochem. 2014, 68, 392–401. [Google Scholar] [CrossRef]
- Puissant, J.; Cecillon, L.; Mills, R.T.E.; Robroek, B.J.M.; Gavazov, K.; De Danieli, S.; Spiegelberger, T.; Buttler, A.; Brun, J.J. Seasonal influence of climate manipulation on microbial community structure and function in mountain soils. Soil Biol. Biochem. 2015, 80, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Tas, N.; Prestat, E.; McFarland, J.W.; Wickland, K.P.; Knight, R.; Berhe, A.A.; Jorgenson, T.; Waldrop, M.P.; Jansson, J.K. Impact of fire on active layer and permafrost microbial communities and metagenomes in an upland Alaskan boreal forest. ISME J. 2014, 8, 1904–1919. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.F.; Xu, S.J.; Li, C.M.; Zhao, L.; Feng, H.Y.; Yue, G.Y.; Ren, Z.W.; Cheng, G.D. The soil carbon/nitrogen ratio and moisture affect microbial community structures in alkaline permafrost-affected soils with different vegetation types on the Tibetan plateau. Res. Microbiol. 2014, 165, 128–139. [Google Scholar] [CrossRef]
- Luyssaert, S.; Jammet, M.; Stoy, P.C.; Estel, S.; Pongratz, J.; Ceschia, E.; Churkina, G.; Don, A.; Erb, K.; Ferlicoq, M.; et al. Land management and land-cover change have impacts of similar magnitude on surface temperature. Nat. Clim. Chang. 2014, 4, 389–393. [Google Scholar] [CrossRef] [Green Version]
- Schuur, E.A.G.; McGuire, A.D.; Schadel, C.; Grosse, G.; Harden, J.W.; Hayes, D.J.; Hugelius, G.; Koven, C.D.; Kuhry, P.; Lawrence, D.M.; et al. Climate change and the permafrost carbon feedback. Nature 2015, 520, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Verges, A.; Steinberg, P.D.; Hay, M.E.; Poore, A.G.B.; Campbell, A.H.; Ballesteros, E.; Heck, K.L.; Booth, D.J.; Coleman, M.A.; Feary, D.A.; et al. The tropicalization of temperate marine ecosystems: Climate-mediated changes in herbivory and community phase shifts. Proc. R. Soc. B Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Cortez, J.; Billes, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Pastor, J.; Post, W.M. Influence of climate, soil-moisture, and succession on forest carbon and nitrogen cycles. Biogeochemistry 1986, 2, 3–27. [Google Scholar] [CrossRef]
- Turner, M.G.; Baker, W.L.; Peterson, C.J.; Peet, R.K. Factors influencing succession: Lessons from large, infrequent natural disturbances. Ecosystems 1998, 1, 511–523. [Google Scholar] [CrossRef]
- Elliott, K.J.; Boring, L.R.; Swank, W.T.; Haines, B.R. Successional changes in plant species diversity and composition after clearcutting a Southern Appalachian watershed. For. Ecol. Manag. 1997, 92, 67–85. [Google Scholar] [CrossRef]
- Reynolds, H.L.; Packer, A.; Bever, J.D.; Clay, K. Grassroots ecology: Plant-microbe-soil interactions as drivers of plant community structure and dynamics. Ecology 2003, 84, 2281–2291. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.C.; Dong, Q.; Wu, C.; Zhou, X.H.; Shi, H.C. Study of the succession of microbial communities for sulfur cycle response to ecological factors change in sediment of sewage system. Environ. Sci. Pollut. Res. 2015, 22, 9250–9259. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Hortal, S.; Armas, C.; Pugnaire, F.I. Interactions among soil, plants, and microorganisms drive secondary succession in a dry environment. Soil Biol. Biochem. 2014, 78, 298–306. [Google Scholar] [CrossRef]
- Scheu, S. Changes in microbial nutrient status during secondary succession and its modification by earthworms. Oecologia 1990, 84, 351–358. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, C.K.; Jiang, L.F.; Luo, Y.Q. Trends in soil microbial communities during secondary succession. Soil Biol. Biochem. 2017, 115, 92–99. [Google Scholar] [CrossRef]
- Kardol, P.; Todd, D.E.; Hanson, P.J.; Mulholland, P.J. Long-term successional forest dynamics: Species and community responses to climatic variability. J. Veg. Sci. 2010, 21, 627–642. [Google Scholar] [CrossRef]
- Cong, J.; Liu, X.D.; Lu, H.; Xu, H.; Li, Y.D.; Deng, Y.; Li, D.Q.; Zhang, Y.G. Available nitrogen is the key factor influencing soil microbial functional gene diversity in tropical rainforest. BMC Microbiol. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Shi, N.N.; Liu, Y.X.; Zheng, Y.; Ding, Q.; Mi, X.C.; Ma, K.P.; Wubet, T.; Buscot, F.; Guo, L.D. Host plant richness explains diversity of ectomycorrhizal fungi: Response to the comment of Tedersoo et al. (2014). Mol. Ecol. 2014, 23, 996–999. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.; Jang, S.; Lee, J.; Lee, H.; Lim, Y.W.; Kim, C.; Kim, J.J. Diversity of Wood-Inhabiting Polyporoid and Corticioid Fungi in Odaesan National Park, Korea. Mycobiology 2016, 44, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kim, S.; Shin, Y.; Choung, Y. Spatial pattern and association of tree species in a mixed Abies holophylla-broadleaved deciduous forest in Odaesan National Park. J. Plant Biol. 2012, 55, 242–250. [Google Scholar] [CrossRef]
- Park, Y.; Kim, J.; Jang, T.; Chae, H.; Takami, Y. Local climate mediates spatial and temporal variation in carabid beetle communities in three forests in Mount Odaesan, Korea. Ecol. Entomol. 2017, 42, 184–194. [Google Scholar] [CrossRef]
- Byun, D.; Lee, H.; Kim, C. Vegetation pattern and successional sere in the forest of Mt. Odae. Korean J. Ecololgy (Korea Repub.) 1998, 21, 283–290. [Google Scholar]
- Mosandl, R.; Kleinert, A. Development of oaks (Quercus petraea (Matt.) Liebl.) emerged from bird-dispersed seeds under old-growth pine (Pinus silvestris L.) stands. For. Ecol. Manag. 1998, 106, 35–44. [Google Scholar] [CrossRef]
- González-Espinosa, M.; Ramírez-Marcial, N.; Galindo-Jaimes, L. Secondary succession in montane pine-oak forests of Chiapas, Mexico. In Ecology and Conservation of Neotropical Montane Oak Forests; Springer: Berlin/Heidelberg, Germany, 2006; pp. 209–221. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Sagova-Mareckova, M.; Cermak, L.; Novotna, J.; Plhackova, K.; Forstova, J.; Kopecky, J. Innovative methods for soil DNA purification tested in soils with widely differing characteristics. Appl. Environ. Microbiol. 2008, 74, 2902–2907. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.; Jang, S.; Min, M.; Hong, J.-H.; Lee, H.; Lee, H.; Lim, Y.W.; Kim, J.-J. Comparison of the diversity of basidiomycetes from dead wood of the Manchurian fir (Abies holophylla) as evaluated by fruiting body collection, mycelial isolation, and 454 sequencing. Microb. Ecol. 2015, 70, 634–645. [Google Scholar] [CrossRef]
- Sundberg, C.; Al-Soud, W.A.; Larsson, M.; Alm, E.; Yekta, S.S.; Svensson, B.H.; Sørensen, S.J.; Karlsson, A. 454 pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 2013, 85, 612–626. [Google Scholar] [CrossRef] [Green Version]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PloS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Kõljalg, U.; Larsson, K.H.; Abarenkov, K.; Nilsson, R.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E. UNITE: A database providing web-based methods for the molecular identification of ectomycorrhizal fungi. New Phytol. 2005, 166, 1063–1068. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJournal Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Findlay, R.H.; Trexler, M.B.; Guckert, J.B.; White, D.C. Laboratory study of disturbance in marine-sediments: Response of a microbial community. Mar. Ecol. Prog. Ser. 1990, 62, 121–133. [Google Scholar] [CrossRef]
- Lee, H.; Heo, Y.M.; Kwon, S.L.; Yoo, Y.; Lee, A.H.; Kwon, B.-O.; Kim, G.-H.; Khim, J.S.; Kim, J.-J. Recovery of the benthic bacterial community in coastal abandoned saltern requires over 35 years: A comparative case study in the Yellow Sea. Environ. Int. 2020, 135, 105412. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.S.; Webb, R.I.; Ridd, M.J.; Hill, R.T.; Negri, A.P. The effects of copper on the microbial community of a coral reef sponge. Environ. Microbiol. 2001, 3, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Iturbe, I.; Isham, V.; Cox, D.R.; Manfreda, S.; Porporato, A. Space-time modeling of soil moisture: Stochastic rainfall forcing with heterogeneous vegetation. Water Resour. Res. 2006, 42. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wei, W.; Chen, L.D.; Chen, W.L.; Wang, J.L. Response of temporal variation of soil moisture to vegetation restoration in semi-arid Loess Plateau, China. Catena 2014, 115, 123–133. [Google Scholar] [CrossRef]
- Gamboa, A.M.; Hidalgo, C.; De Leon, F.; Etchevers, J.D.; Gallardo, J.F.; Campo, J. Nutrient addition differentially affects soil carbon sequestration in secondary tropical dry forests: Early- versus late-succession stages. Restor. Ecol. 2010, 18, 252–260. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Bader, N.E.; Johnson, D.W.; Cheng, W.X. Does accelerated soil organic matter decomposition in the presence of plants increase plant N availability? Soil Biol. Biochem. 2009, 41, 1080–1087. [Google Scholar] [CrossRef]
- Aponte, C.; Maranon, T.; Garcia, L.V. Microbial C, N and P in soils of Mediterranean oak forests: Influence of season, canopy cover and soil depth. Biogeochemistry 2010, 101, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt-Romermann, M.; Kudernatsch, T.; Pfadenhauer, J.; Kirchner, M.; Jakobi, G.; Fischer, A. Long-term effects of nitrogen deposition on vegetation in a deciduous forest near Munich, Germany. Appl. Veg. Sci. 2007, 10, 399–406. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Scow, K.M. Soil moisture and plant residue addition interact in their effect on extracellular enzyme activity. Pedobiologia 2011, 54, 71–78. [Google Scholar] [CrossRef]
- Castaño, C.; Lindahl, B.D.; Alday, J.G.; Hagenbo, A.; Martínez de Aragón, J.; Parladé, J.; Pera, J.; Bonet, J.A. Soil microclimate changes affect soil fungal communities in a Mediterranean pine forest. New Phytol. 2018, 220, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Steinweg, J.M.; Dukes, J.S.; Wallenstein, M.D. Modeling the effects of temperature and moisture on soil enzyme activity: Linking laboratory assays to continuous field data. Soil Biol. Biochem. 2012, 55, 85–92. [Google Scholar] [CrossRef]
- Aulakh, M.S.; Aulakh, N.S. Interactions of nitrogen with other nutrients and water: Effect on crop yield and quality, nutrient use efficiency, carbon sequestration, and environmental pollution. Adv. Agron. 2005, 86, 341–409. [Google Scholar] [CrossRef]
- Lafleur, B.; Hooper-Bui, L.M.; Mumma, E.P.; Geaghan, J.P. Soil fertility and plant growth in soils from pine forests and plantations: Effect of invasive red imported fire ants Solenopsis invicta (Buren). Pedobiologia 2005, 49, 415–423. [Google Scholar] [CrossRef]
- Noh, N.J.; Son, Y.; Lee, S.K.; Seo, K.W.; Heo, S.J.; Yi, M.J.; Park, P.S.; Kim, R.H.; Son, Y.M.; Lee, K.H. Carbon and nitrogen storage in an age-sequence of Pinus densiflora stands in Korea. Sci. China Life Sci. 2010, 53, 822–830. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil Microbial Community Responses to Multiple Experimental Climate Change Drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Bezemer, T.M.; Lawson, C.S.; Hedlund, K.; Edwards, A.R.; Brook, A.J.; Igual, J.M.; Mortimer, S.R.; Van der Putten, W.H. Plant species and functional group effects on abiotic and microbial soil properties and plant-soil feedback responses in two grasslands. J. Ecol. 2006, 94, 893–904. [Google Scholar] [CrossRef]
- Kämpfer, P.; Young, C.-C.; Arun, A.; Shen, F.-T.; Jäckel, U.; Rossello-Mora, R.; Lai, W.-A.; Rekha, P. Pseudolabrys taiwanensis gen. nov., sp. nov., an alphaproteobacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2006, 56, 2469–2472. [Google Scholar]
- Liu, X.-Z.; Wang, Q.-M.; Göker, M.; Groenewald, M.; Kachalkin, A.; Lumbsch, H.T.; Millanes, A.; Wedin, M.; Yurkov, A.; Boekhout, T. Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [Green Version]
- Cong, J.; Yang, Y.F.; Liu, X.D.; Lu, H.; Liu, X.; Zhou, J.Z.; Li, D.Q.; Yin, H.Q.; Ding, J.J.; Zhang, Y.G. Analyses of soil microbial community compositions and functional genes reveal potential consequences of natural forest succession. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Bonfante, P.; Anca, I.A. Plants, Mycorrhizal Fungi, and Bacteria: A Network of Interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppe, B.; Kahl, T.; Karasch, P.; Wubet, T.; Bauhus, J.; Buscot, F.; Kruger, D. Network analysis reveals ecological links between N-fixing bacteria and wood-decaying fungi. PloS ONE 2014, 9, e88141. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Oh, S.-Y.; Lee, Y.M.; Jang, Y.; Jang, S.; Kim, C.; Lim, Y.W.; Kim, J.-J. Successional Variation in the Soil Microbial Community in Odaesan National Park, Korea. Sustainability 2020, 12, 4795. https://doi.org/10.3390/su12114795
Lee H, Oh S-Y, Lee YM, Jang Y, Jang S, Kim C, Lim YW, Kim J-J. Successional Variation in the Soil Microbial Community in Odaesan National Park, Korea. Sustainability. 2020; 12(11):4795. https://doi.org/10.3390/su12114795
Chicago/Turabian StyleLee, Hanbyul, Seung-Yoon Oh, Young Min Lee, Yeongseon Jang, Seokyoon Jang, Changmu Kim, Young Woon Lim, and Jae-Jin Kim. 2020. "Successional Variation in the Soil Microbial Community in Odaesan National Park, Korea" Sustainability 12, no. 11: 4795. https://doi.org/10.3390/su12114795
APA StyleLee, H., Oh, S. -Y., Lee, Y. M., Jang, Y., Jang, S., Kim, C., Lim, Y. W., & Kim, J. -J. (2020). Successional Variation in the Soil Microbial Community in Odaesan National Park, Korea. Sustainability, 12(11), 4795. https://doi.org/10.3390/su12114795