Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Collection
2.3. Medium
2.4. Preparation of Enzyme Extract-Containing Microcapsules (MC)
2.5. Experimental Setting
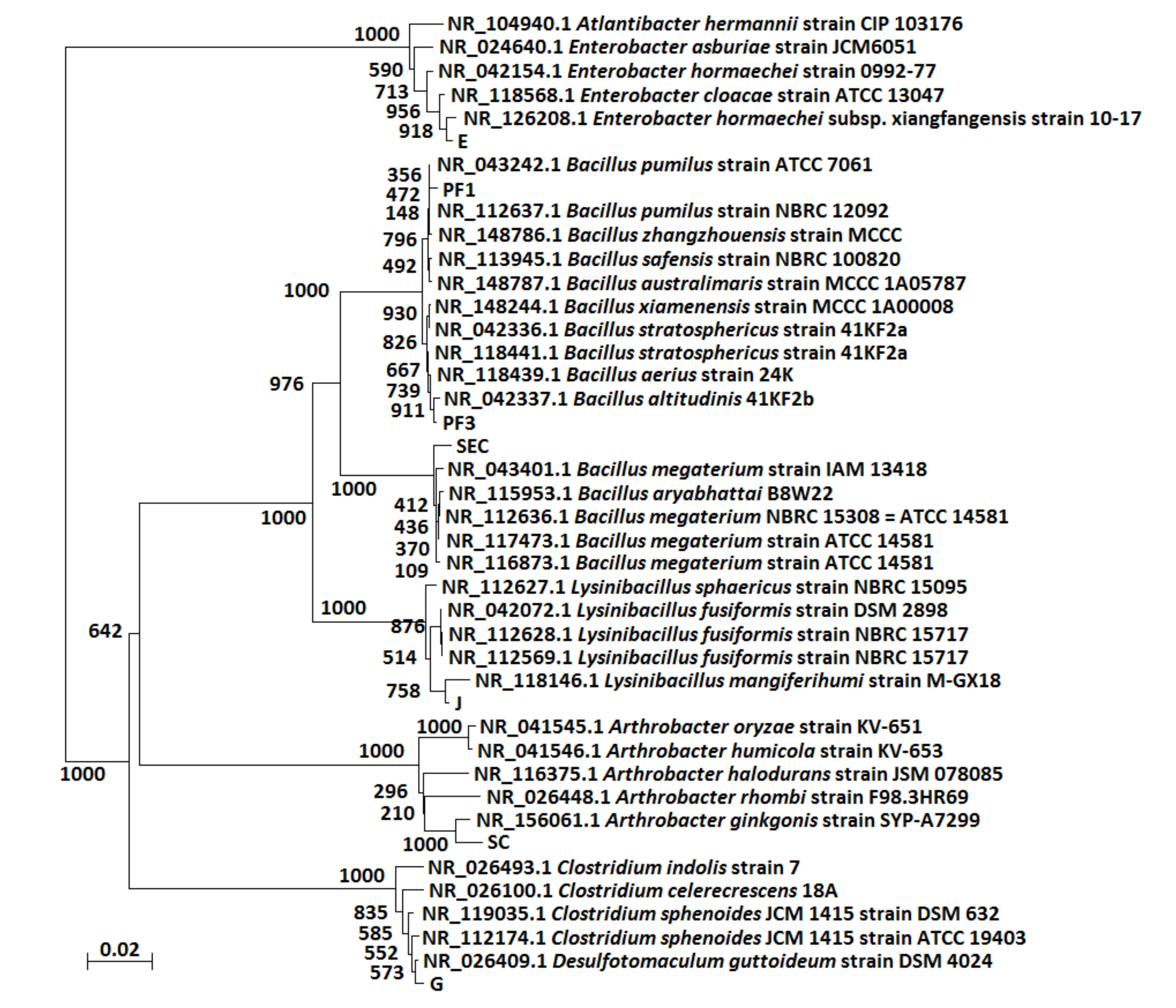
2.6. Isolation, Identification and Tests of the ACE-Degrading Bacteria
2.7. Analytical Methods
2.8. Microbial Community Analysis
3. Results and Discussion
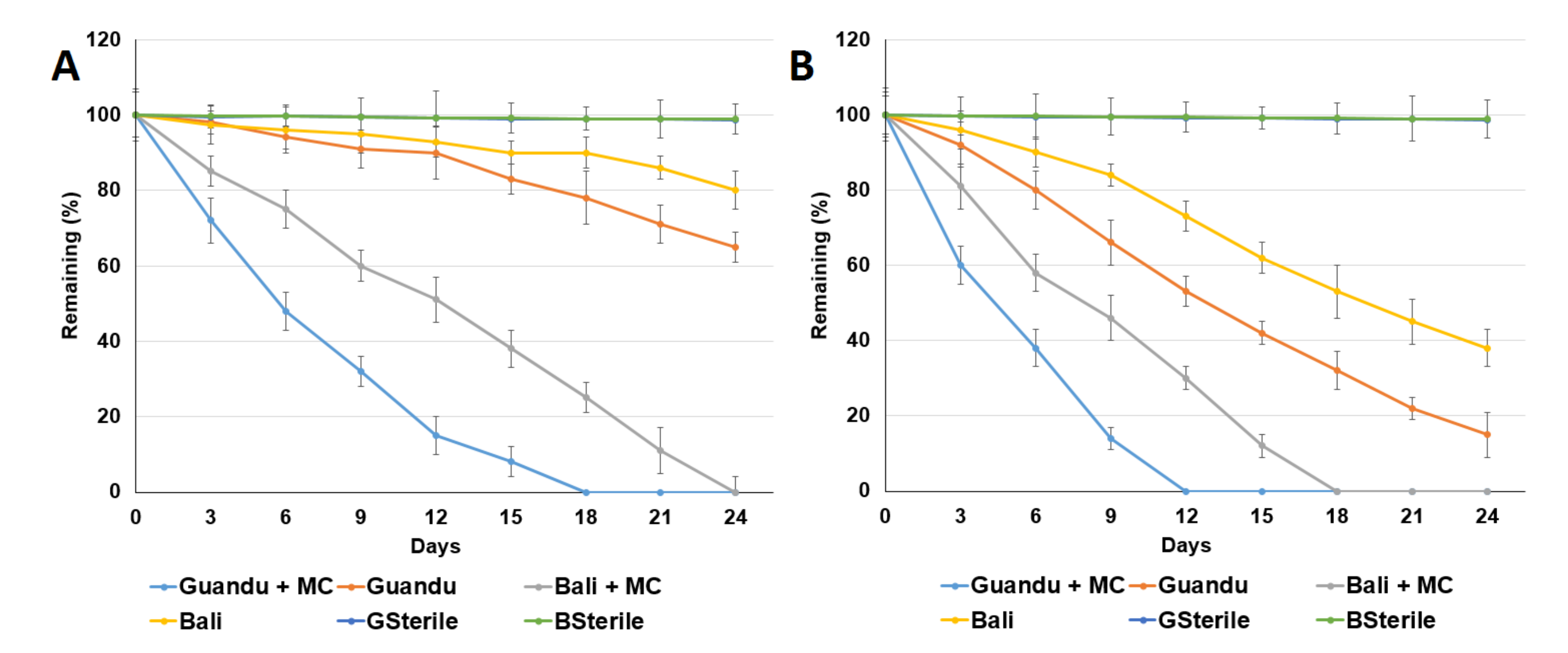
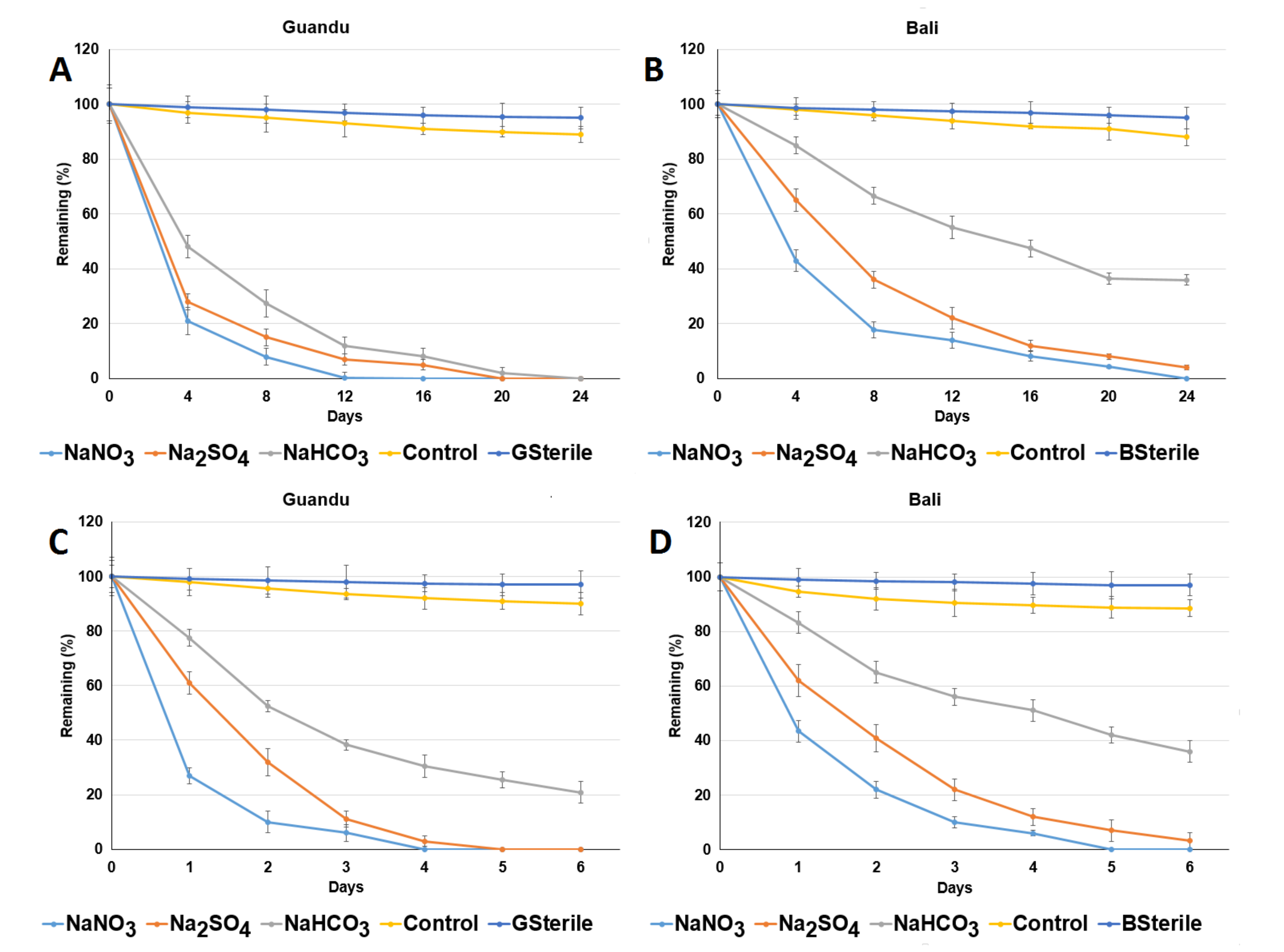
3.1. Aerobic ACE Degradation
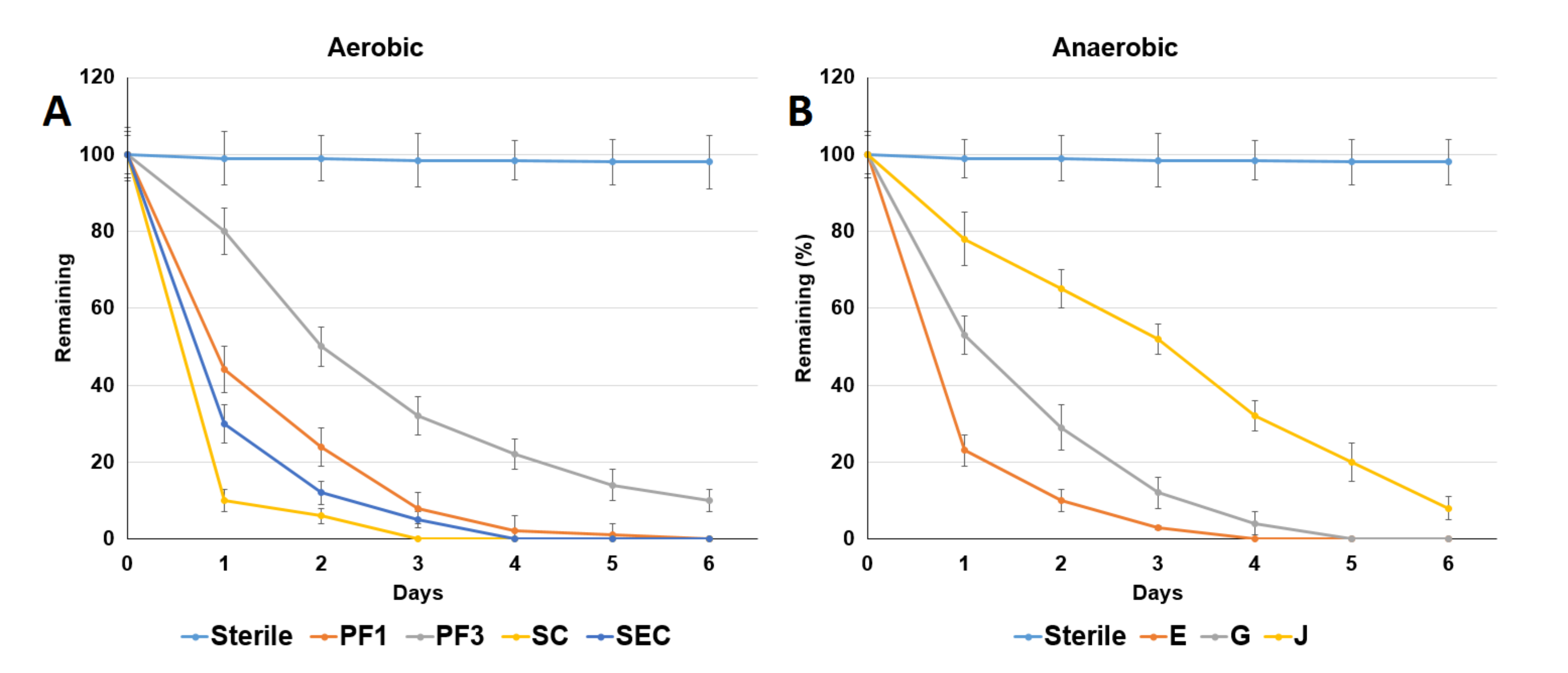
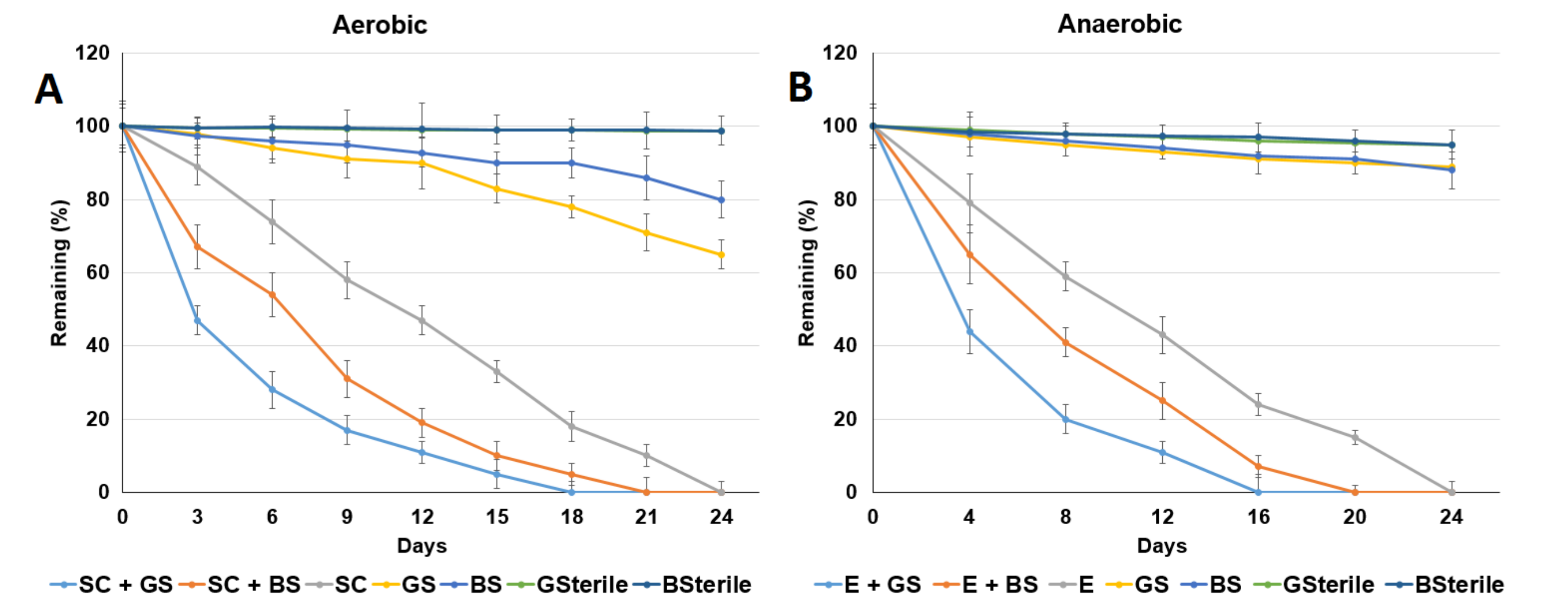
3.2. ACE Degradation by ACE-Degrading Bacteria
3.3. Addition of Electron Acceptors Improved Anaerobic ACE Degradation
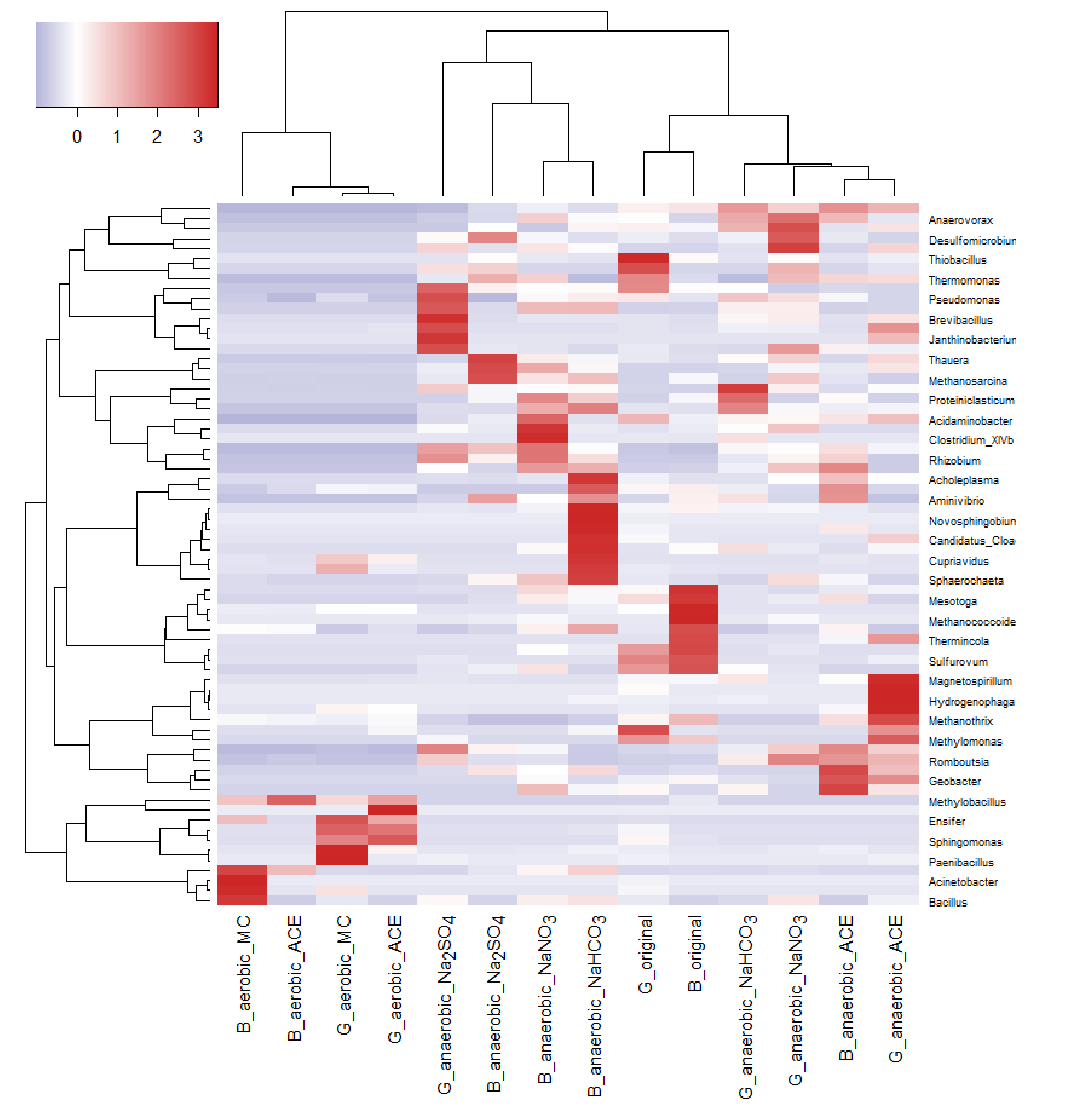
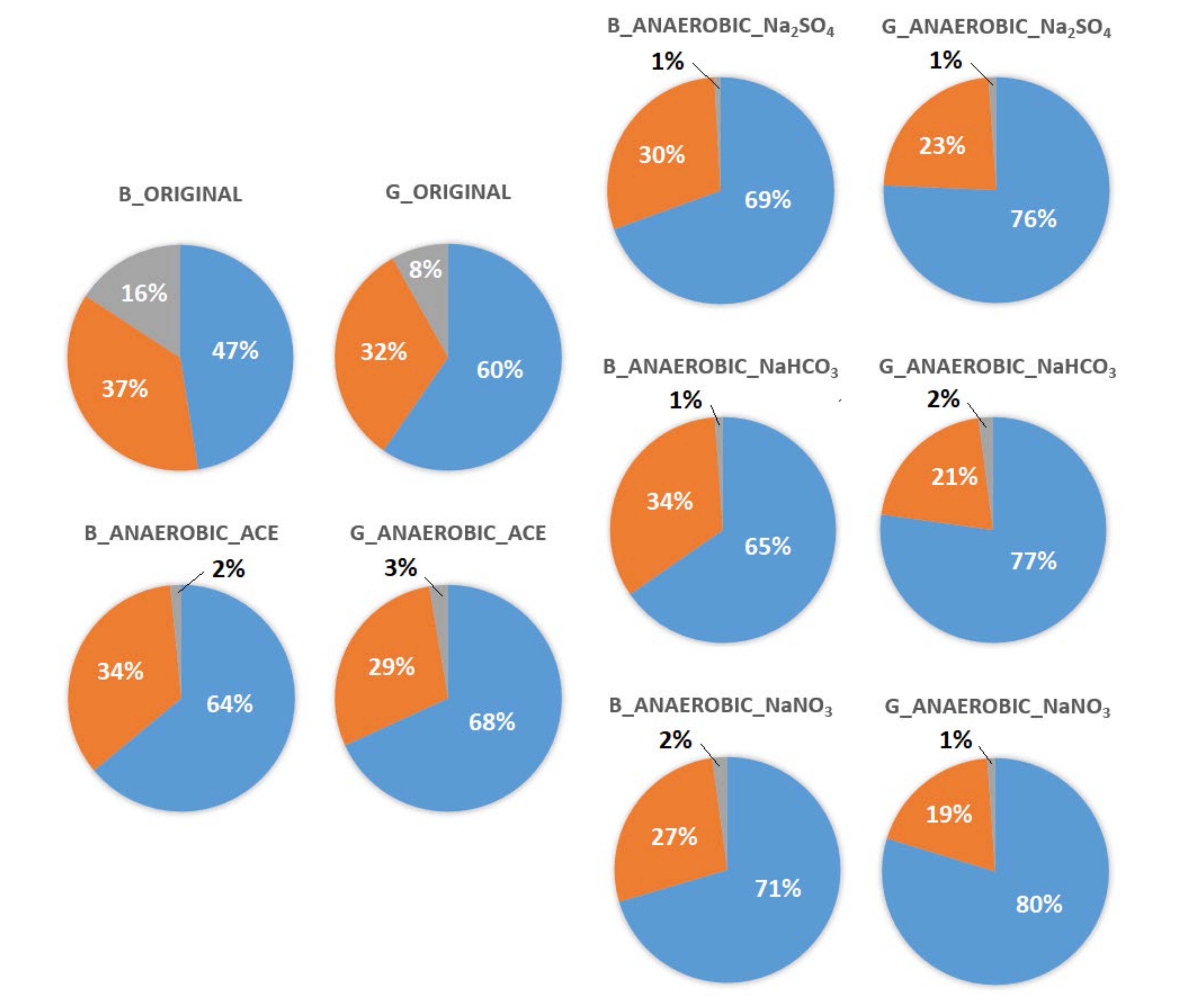
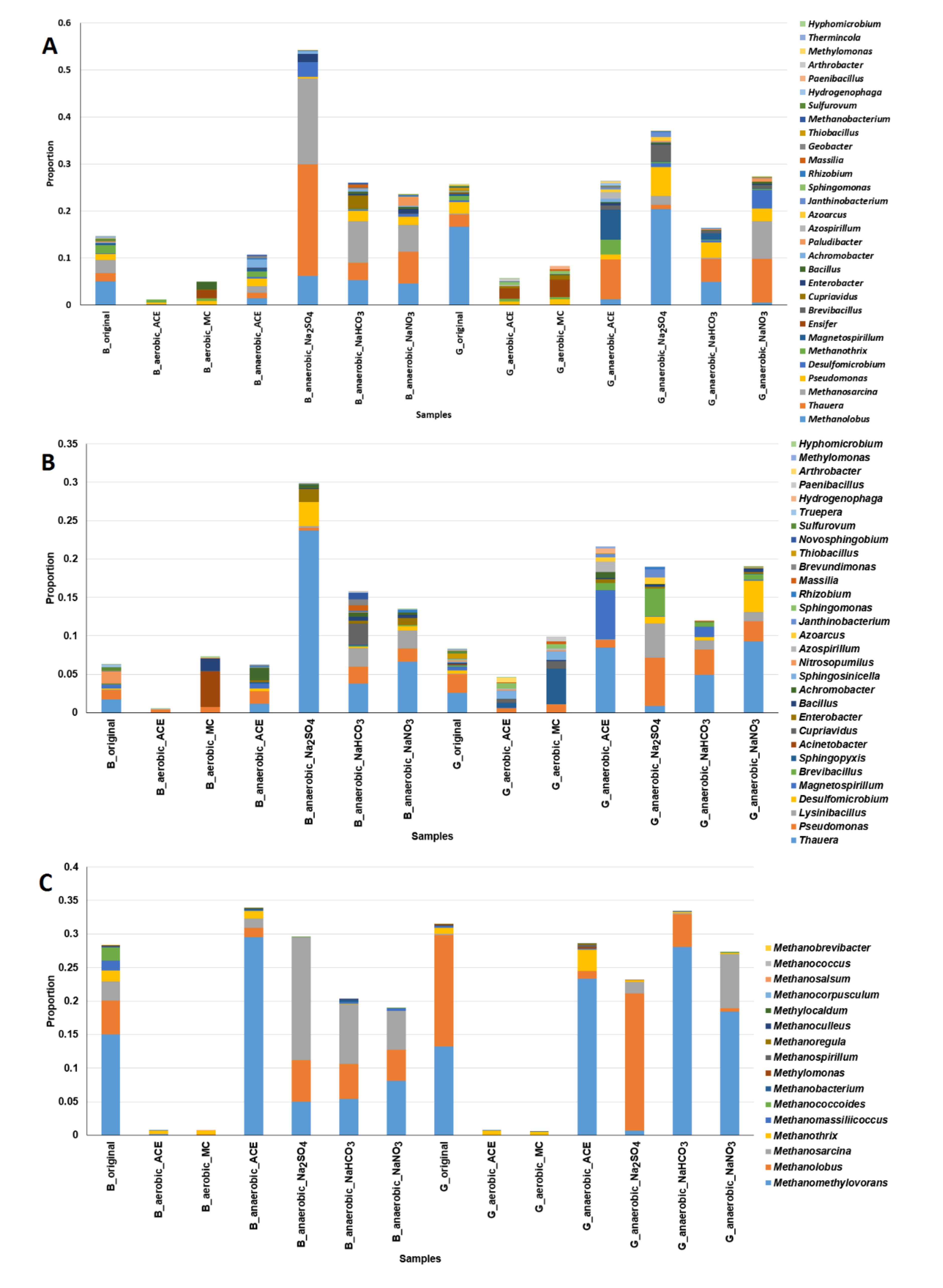
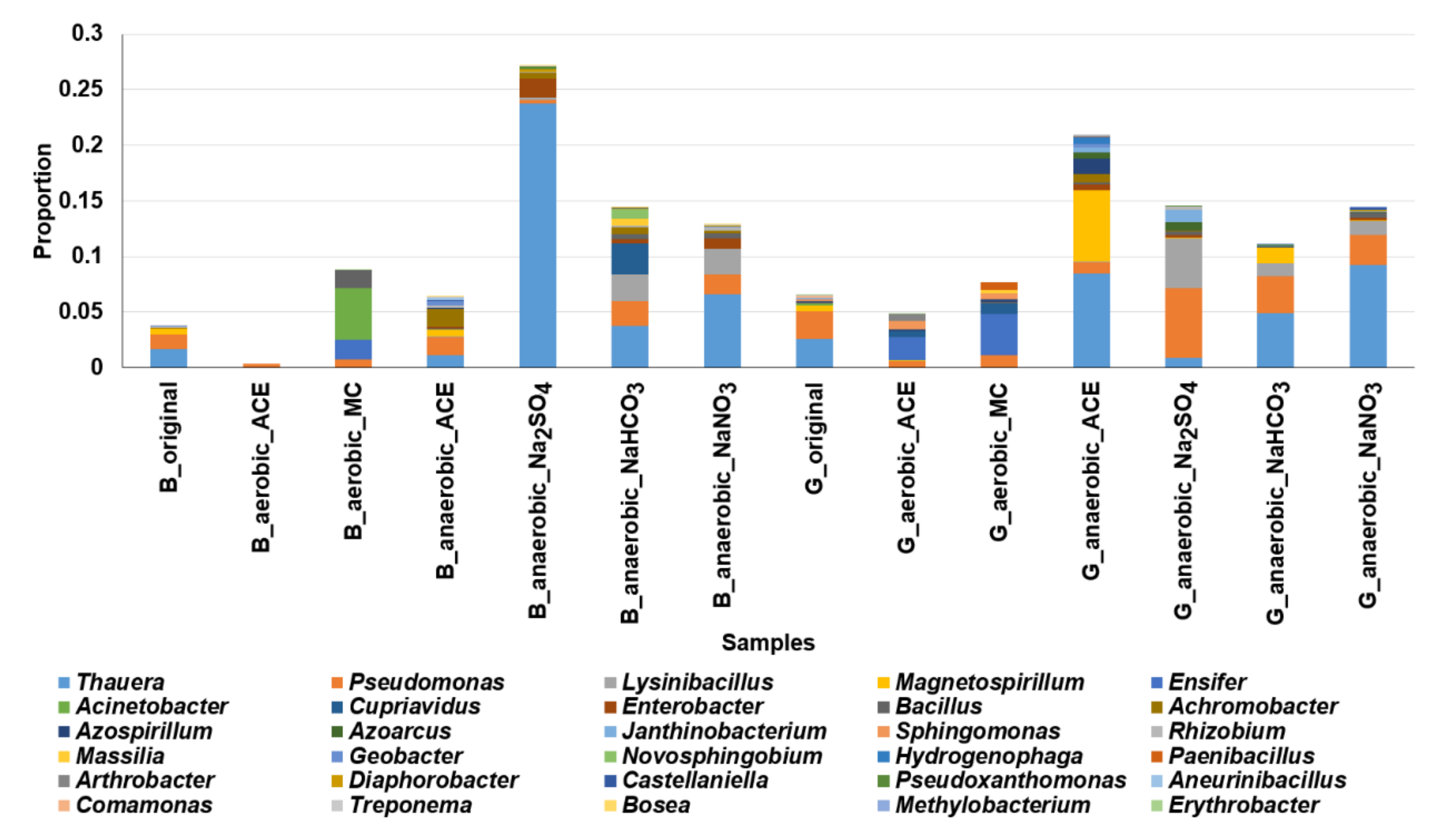
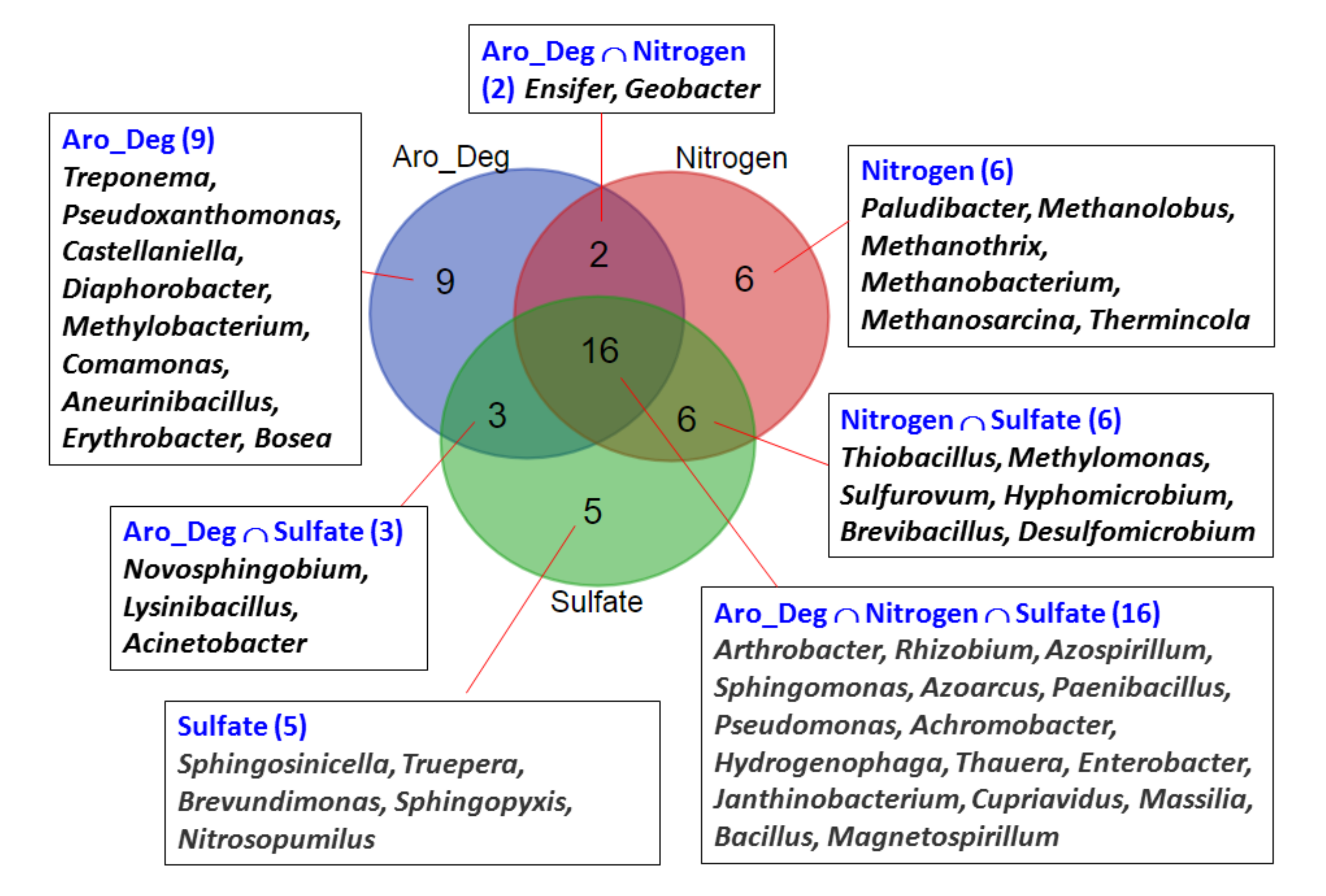
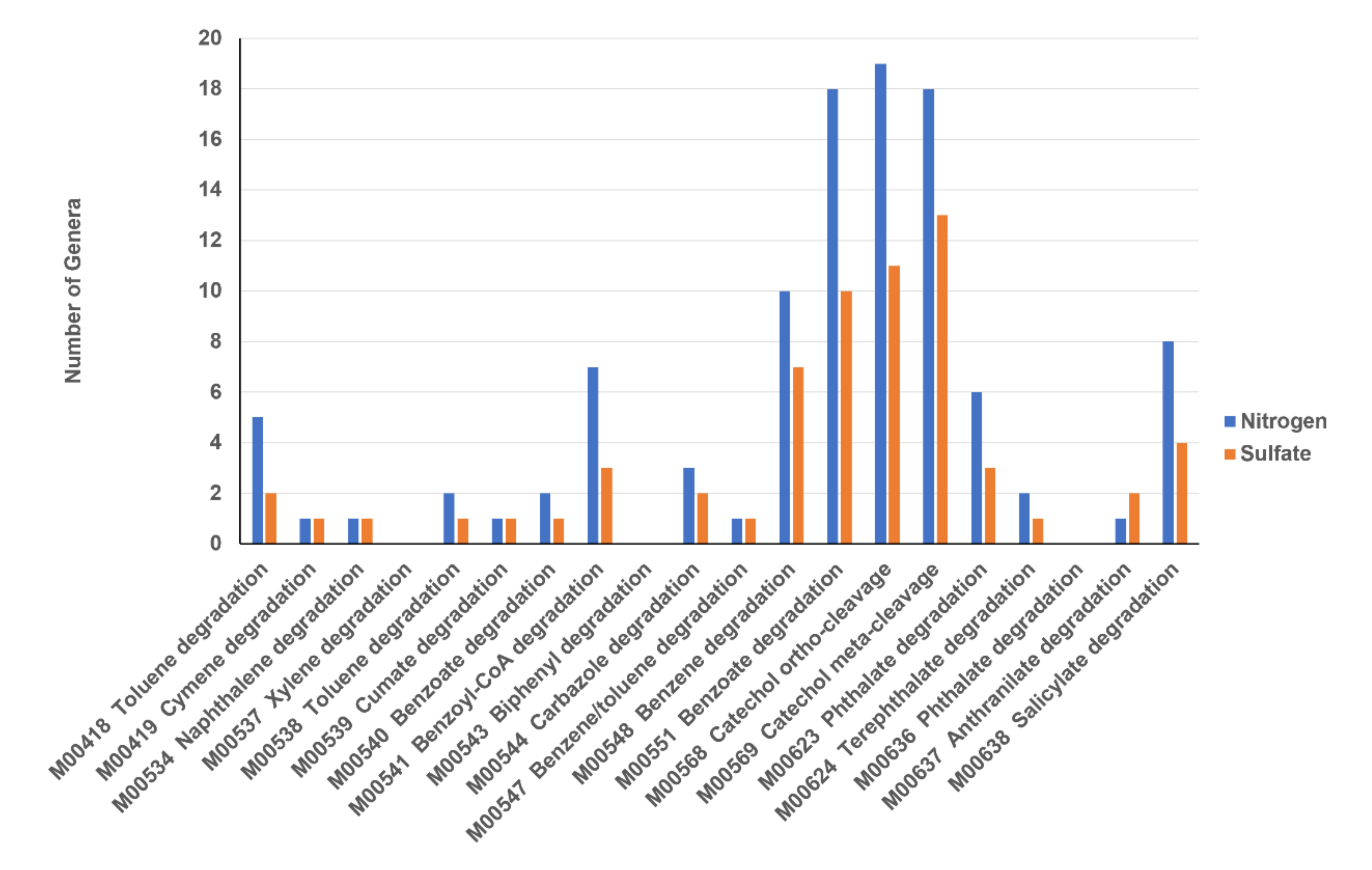
3.4. Analysis of Microbial Communities Associated with ACE Degradation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ouellet, M.; Percival, M.D. Acetaminophen inhibition of cyclooxygenase isoforms. Arch. Biochem. Biophys. 2001, 387, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.D.; Bartkow, M.; Blockwell, S.J.; Coleman, H.M.; Khan, S.J.; Lim, R.; McDonald, J.A.; Nice, H.; Nugegoda, D.; Pettigrove, V.; et al. A national survey of trace organic contaminants in Australian Rivers. J. Environ. Qual. 2014, 43, 1702–1712. [Google Scholar] [CrossRef] [PubMed]
- Campanha, M.B.; Awan, A.T.; De Sousa, D.N.R.; Grosseli, G.M.; Mozeto, A.A.; Fadini, P.S. A 3-year study on occurrence of emerging contaminants in an urban stream of So Paulo State of Southeast Brazil. Environ. Sci. Pollut. Res. 2015, 22, 7936–7947. [Google Scholar] [CrossRef]
- Tran, N.H.; Reinhard, M.; Khan, E.; Chen, H.; Nguyen, V.T.; Li, Y.; Goh, S.G.; Nguyen, Q.B.; Saeidi, N.; Gin, K.Y.H. Emerging contaminants in wastewater, stormwater runoff, and surface water: Application as chemical markers for diffuse sources. Sci. Total Environ. 2019, 676, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Zhang, M.; Yuan, S.; Feng, J.; Wang, Q.; Wang, W.; Hu, Z. Transformation of acetaminophen during water chlorination treatment: Kinetics and transformation products identification. Environ. Sci. Pollut. Res. Int. 2016, 23, 12303–12311. [Google Scholar] [CrossRef]
- Sun, C.; Dudley, S.; McGinnis, M.; Trumble, J.; Gan, J. Acetaminophen detoxification in cucumber plants via induction of glutathione S-transferases. Sci. Total Environ. 2019, 649, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Philips, P.J.; Smith, S.G.; Koplin, D.W.; Zaugg, S.D.; Buxton, H.T.; Furlong, E.T.; Esposito, K.; Stinson, B. Pharmaceutical formulation facilities as sources of opioids and other pharmaceuticals to wastewater treatment plant effluents. Environ. Sci. Technol. 2010, 44, 4910–4916. [Google Scholar] [CrossRef]
- Escher, B.I.; Baumgartner, R.; Koller, M.; Treyer, K.; Lienert, J.; McArdell, C.S. Environmental toxicology and risk assessment of pharmaceuticals from hospital wastewater. Water Res. 2011, 45, 75–92. [Google Scholar] [CrossRef] [Green Version]
- Bernard, D.; Pascaline, H.; Jeremie, J.J. Distribution and origin of hydrocarbons in sediments from lagoons with fringing mangrove communities. Mar. Pollut. Bull. 1996, 32, 734–739. [Google Scholar] [CrossRef]
- Chang, B.V.; Chang, I.T.; Yuan, S.Y. Anaerobic degradation of phenanthrene and pyrene in mangrove sediment. Bull. Environ. Contam. Toxicol. 2008, 80, 145–149. [Google Scholar] [CrossRef]
- Chang, B.V.; Lu, Z.J.; Yuan, S.Y. Anaerobic degradation of nonylphenol in subtropical mangrove sediments. J. Hazard. Mater. 2009, 165, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.W.; Tsai, L.L.; Chang, B.V. Fungi extracellular enzyme-containing microcapsules enhance degradation of sulfonamide antibiotics in mangrove sediments. Environ. Sci. Pollut. Res. 2017, 25, 10069–10079. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.W.; Tsai, L.L.; Chang, B.V. Anaerobic degradation of sulfamethoxazole in mangrove sediments. Sci. Total Environ. 2018, 634, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Żur, J.; Wojcieszyńska, D.; Hupert-Kocurek, K.; Marchlewicz, A.; Guzik, U. Paracetamol—Toxicity and microbial utilization. Pseudomonas moorei KB4 as a case study for exploring degradation pathway. Chemosphere 2018, 20, 192–202. [Google Scholar]
- Joss, A.; Keller, E.; Alder, A.C.; Gobel, A.; McArdell, C.S.; Ternes, T.; Siegrist, H. Removal of pharmaceuticals and fragrances in biological wastewater treatment. Water Res. 2005, 39, 3139–3152. [Google Scholar] [CrossRef]
- Groning, J.; Held, C.; Garten, C.; Claussnitzer, U.; Kaschabek, S.R.; Schlomann, M. Transformation of diclofenac by the indigenous microflora of river sediments and identification of a major intermediate. Chemosphere 2007, 69, 509–516. [Google Scholar] [CrossRef]
- Ying, G.G.; Kookana, R.S. Degradation of five selected endocrine-disrupting chemicals in seawater and marine sediment. Environ. Sci. Technol. 2003, 37, 1256–1260. [Google Scholar] [CrossRef]
- Pepper, L.L.; Gerba, C.P.; Gentry, T.J. Environmental Microbiology, 3rd ed.; Elsevier BV: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Hu, J.; Zhang, L.L.; Chen, J.M.; Liu, Y. Degradation of paracetamol by Pseudomonas aeruginosa strain HJ1012. J. Environ. Sci. Health A Tox. Hazard Subst. Environ. Eng. 2013, 48, 791–799. [Google Scholar] [CrossRef]
- De Gusseme, B.; Vanhaecke, L.; Verstraete, W.; Boon, N. Degradation of acetaminophen by Delftia tsuruhatensis and Pseudomonas aeruginosa in a membrane bioreactor. Water Res. 2011, 45, 1829–1837. [Google Scholar] [CrossRef]
- Li, X.; Lin, X.; Zhang, J.; Wu, Y.; Yin, R.; Feng, Y.; Wang, Y. Degradation of polycyclic aromatic hydrocarbons by crude extracts from spent mushroom substrate and its possible mechanisms. Curr. Microbiol. 2010, 60, 336–342. [Google Scholar] [CrossRef]
- Chang, B.V.; Ren, Y.L. Biodegradation of three tetracyclines in river sediment. Ecol. Eng. 2015, 75, 272–277. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Carvajal, L.C.; Sanz-Martín, J.L.; Barragán-Huerta, B.E. Biodegradation of organic pollutants in saline wastewater by halophilic microorganisms: A review. Environ. Sci. Pollut. Res. 2014, 21, 9578–9588. [Google Scholar] [CrossRef]
- Tam, N.F.; Guo, C.L.; Yau, W.Y.; Wong, Y.S. Preliminary study on biodegradation of phenanthrene by bacteria isolated from mangrove sediments in Hong Kong. Mar. Pollut. Bull. 2002, 45, 316–324. [Google Scholar] [CrossRef]
- Varma, S.S.; Lakshmi, M.B.; Rajagopal, P.; Velan, M. Degradation of total petroleum hydrocarbon (TPH) in contaminated soil using Bacillus pumilus MVSV3. Biocontrol. Sci. 2017, 22, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanapiah, M.; Zulkifli, S.Z.; Mustafa, M.; Mohamat-Yusuff, F.; Ismail, A. Isolation, characterization, and identification of potential diuron-degrading bacteria from surface sediments of Port Klang, Malaysia. Mar. Pollu. Bull. 2018, 127, 453–457. [Google Scholar] [CrossRef]
- Zainith, S.; Purchase, D.; Saratale, G.D.; Ferreira, L.F.R.; Bilal, M.; Bharagava, R.N. Isolation and characterization of lignin-degrading bacterium Bacillus aryabhattai from pulp and paper mill wastewater and evaluation of its lignin-degrading potential. 3 Biotech. 2019, 9, 92. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, M.Y.; Zhao, J.C.; Xu, J.; Zhang, Y.; Zhang, T.Y.; Wu, Y.Y.; Zhang, Y.X. Arthrobacter ginkgonis sp. nov., an actinomycete isolated from rhizosphere of Ginkgo biloba L. Int. J. Syst. Evol. Microbiol. 2017, 67, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Aissaoui, S.; Ouled-Haddar, H.; Sifour, M.; Harrouche, K.; Sghaier, H. Metabolic and co-metabolic transformation of diclofenac by Enterobacter hormaechei D15 isolated from activated sludge. Curr. Microbiol. 2017, 74, 381–388. [Google Scholar] [CrossRef]
- Zhong, G.F.; Wang, F.F.; Sun, J.H.; Ye, J.B.; Mao, D.B.; Ma, K.; Yang, X.P. Bioconversion of lutein by Enterobacter hormaechei to form a new compound, 8-methyl-alpha-ionone. Biotech. Lett. 2017, 39, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.J.; Dodge, C.J.; Meinken, G.E. Biotransformation of pertechnetate by Clostridia. Radiochim. Acta 2009, 90, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Sari, I.P.; Simarani, K. Decolorization of selected azo dye by Lysinibacillus fusiformis W1B6: Biodegradation optimization, isotherm, and kinetic study biosorption mechanism. Adsorp. Sci. Technol. 2019, 37, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.J.; Chang, Y.H.; Chao, W.L.; Jane, W.N.; Chang, Y.T. Effect of hydraulic retention time on electricity generation using a solid plain-graphite plate microbial fuel cell anoxic/oxic process for treating pharmaceutical sewage. J. Environ. Sci. Health Pert A 2018, 53, 1185–1197. [Google Scholar] [CrossRef]
- Chang, Y.T.; Yang, C.W.; Chang, Y.J.; Chang, T.C.; Wei, D.J. The treatment of PPCP-containing sewage in an anoxic/aerobic reactor coupled with a novel design of solid plain graphite-plates microbial fuel cell. Biomed. Res. Int. 2014, 2014, 765652. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, J.; Zhu, R.; Zhou, Q.; Chen, J. Degradation of paracetamol by pure bacterial cultures and their microbial consortium. Appl. Microbiol. Biotechnol. 2013, 97, 3687–3698. [Google Scholar] [CrossRef]
- Weng, B.; Xie, X.; Yang, J.; Liu, J.; Lu, H.; Yan, C. Research on the nitrogen cycle in rhizosphere of Kandelia obovata under ammonium and nitrate addition. Mar. Pollut. Bull. 2013, 76, 227–240. [Google Scholar] [CrossRef]
- Cui, X.; Song, W.; Feng, J.; Jia, D.; Guo, J.; Wang, Z.; Wu, H.; Qi, F.; Liang, J.; Lin, G. Increased nitrogen input enhances Kandelia obovata seedling growth in the presence of invasive Spartina alterniflora in subtropical regions of China. Biol. Lett. 2017, 13, 20160760. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Tam, N.F.Y.; Zan, Q.; Cheung, S.G.; Shin, P.K.S.; Wong, Y.S.; Zhang, L.; Chen, Z. Performance and bacterial community structure of a 10-years old constructed mangrove wetland. Mar. Pollut. Bull. 2017, 124, 1096–1105. [Google Scholar] [CrossRef]
- Jian, L.; Junyi, Y.; Jingchun, L.; Chongling, Y.; Haoliang, L.; Spencer, K.L. The effects of sulfur amendments on the geochemistry of sulfur, phosphorus and iron in the mangrove plant (Kandelia obovata (S. L.)) rhizosphere. Mar. Pollut. Bull. 2017, 114, 733–741. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Guandu Sampling Sites | Bali Sampling Sites |
|---|---|---|
| Bacterial counts | (1.9 ± 0.2) × 106 CFU g−1 | (2.6 ± 0.3) × 105 CFU g−1 |
| Temperature | 18.1 ± 1.1 °C | 20.5 ± 1.9 °C |
| Salinities | 11.1‰ ± 0.3‰ | 16.5‰ ± 1.5‰ |
| TOC | 3.6 wt% ± 0.2 wt% | 2.5 wt% ± 0.4 wt% |
| ACE concentrations | 0.89 mg kg−1 | 0.45 mg kg−1 |
| Strain | Name | Identity | Accession Number |
|---|---|---|---|
| PF1 | Bacillus pumilus strain NBRC 12092 | 1251/1255 (99%) | NR_112637 |
| PF3 | Bacillus aerius strain 24K | 1326/1326 (100%) | NR_118439 |
| SC | Arthrobacter ginkgonis strain SYP-A7299 | 1283/1302 (99%) | NR_156061 |
| SEC | Bacillus aryabhattai B8W22 | 1330/1330 (100%) | NR_115953 |
| E | Enterobacter hormechei subsp. xiangfangensis strain 10–17 | 1292/1299 (99%) | NR_126208 |
| G | Clostridium sphenoides JCM 1415 strain ATCC 19403 | 1260/1268 (99%) | NR_026409 |
| J | Lysinibacillus fusiformis strain NBRC15717 | 1301/1305 (99%) | NR_112569 |
| Treatment | Guandu Sediment | Bali Sediment | ||
|---|---|---|---|---|
| Without Additives | With Additives | Without Additives | With Additives | |
| Aerobic conditions | ||||
| MC | 33.55 ± 1.67 | 3.24 ± 0.16 | 66.66 ± 3.33 | 6.25 ± 0.31 |
| Arthrobacter sp. | 33.55 ± 1.67 | 2.54 ± 0.13 | 66.66 ± 3.33 | 3.30 ± 0.17 |
| Anaerobic conditions | ||||
| Enterobacter sp. | 95.59 ± 4.63 | 2.62 ± 0.13 | 98.04 ± 4.90 | 3.52 ± 0.17 |
| NaNO3 | 95.59 ± 4.63 | 1.16 ± 0.06 | 98.04 ± 4.90 | 3.05 ± 0.15 |
| Na2SO4 | 95.59 ± 4.63 | 2.39 ± 0.12 | 98.04 ± 4.90 | 3.84 ± 0.19 |
| NaHCO3 | 95.59 ± 4.63 | 2.79 ± 0.14 | 98.04 ± 4.90 | 10.75 ± 0.53 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-W.; Chen, Y.-E.; Chang, B.-V. Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment. Sustainability 2020, 12, 5410. https://doi.org/10.3390/su12135410
Yang C-W, Chen Y-E, Chang B-V. Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment. Sustainability. 2020; 12(13):5410. https://doi.org/10.3390/su12135410
Chicago/Turabian StyleYang, Chu-Wen, Yi-En Chen, and Bea-Ven Chang. 2020. "Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment" Sustainability 12, no. 13: 5410. https://doi.org/10.3390/su12135410
APA StyleYang, C. -W., Chen, Y. -E., & Chang, B. -V. (2020). Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment. Sustainability, 12(13), 5410. https://doi.org/10.3390/su12135410