Glyphosate Resistance in Sonchus oleraceus and Alternative Herbicide Options for Its Control in Southeast Australia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
2.1.1. Experiment 1. Effect of Temperature on Glyphosate Efficacy
2.1.2. Experiment 2. Performance of Different Post-Emergence Herbicides
2.1.3. Experiment 3. Effect of Sorghum Residue Amount on Efficacy of Pre-Emergence Herbicides
2.2. Statistical Analyses
3. Results and Discussion
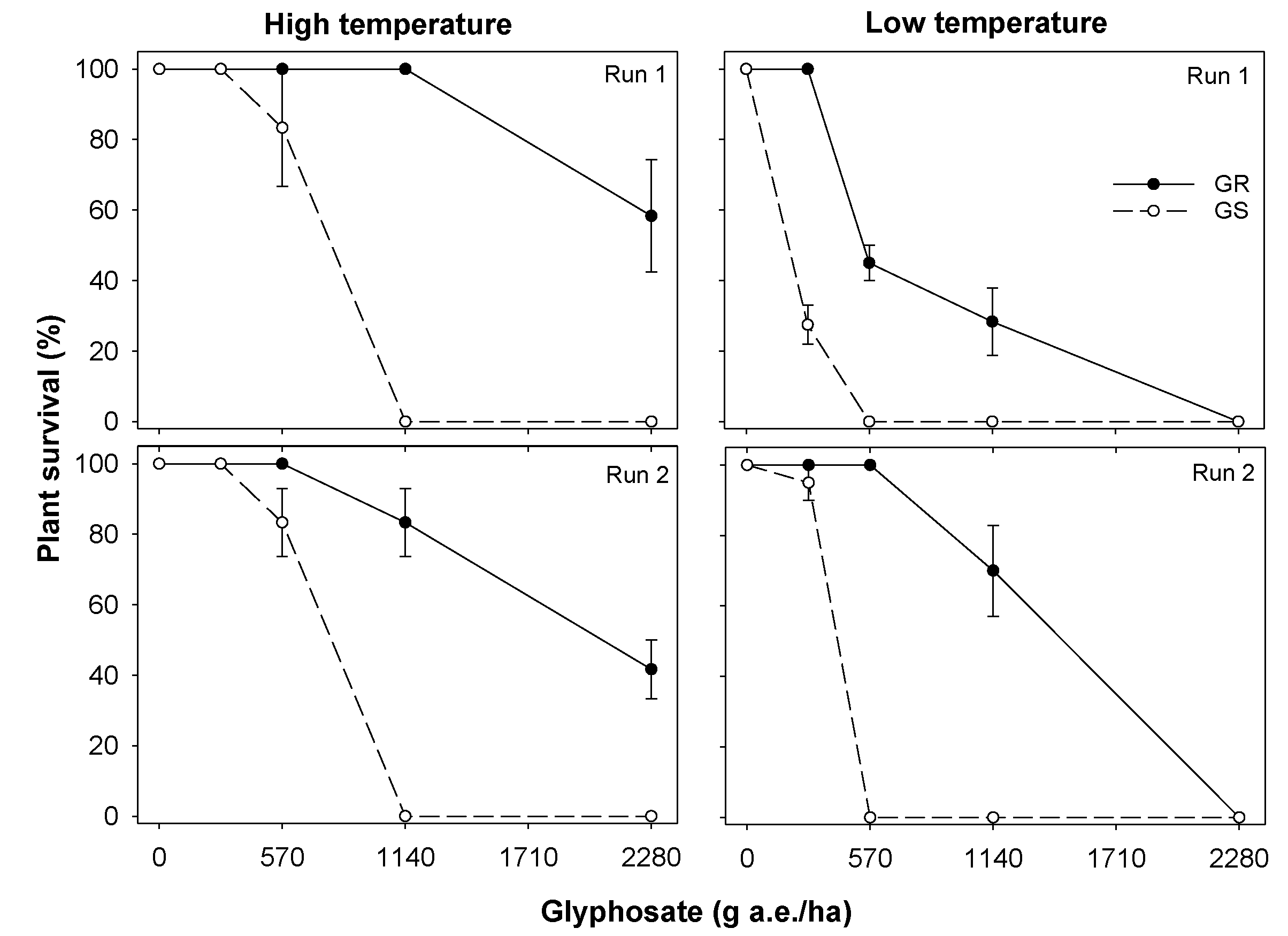
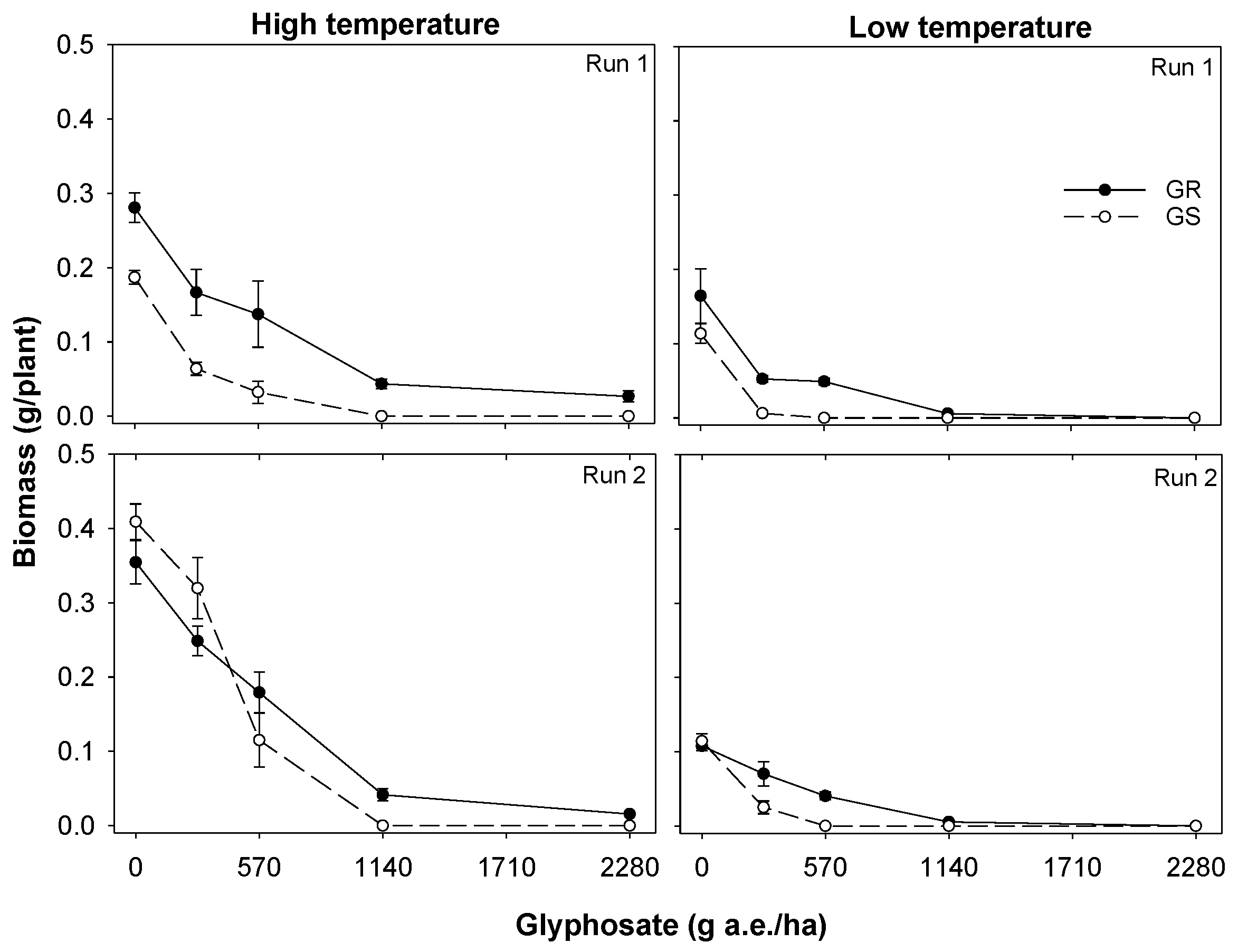
3.1. Experiment 1. Effect of Temperature on Glyphosate Efficacy
3.2. Experiment 2. Performance of Different Post-Emergence Herbicides
3.3. Experiment 3. Effect of Sorghum Residue Amount on Efficacy of Pre-Emergence Herbicides
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Llewellyn, R.; Ronning, D.; Clarke, M.; Mayfield, A.; Walker, S.; Ouzman, J. Impact of Weeds on Australian Grain Production: The Cost of Weeds to Australian Grain Growers and the Adoption of Weed Management and Tillage Practices; Report for GRDC; CSIRO: Canberra, ACT, Australia, 2016; p. 112. [Google Scholar]
- Manalil, S.; Ali, H.H.; Chauhan, B.S. Interference of annual sowthistle (Sonchus oleraceus) in wheat. Weed Sci. 2020, 68, 98–103. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Gill, G.; Preston, C. Factors affecting seed germination of annual sowthistle (Sonchus oleraceus) in southern Australia. Weed Sci. 2006, 54, 854–860. [Google Scholar] [CrossRef]
- Mobli, A.; Matloob, A.; Chauhan, B.S. The response of glyphosate-resistant and glyphosate-susceptible biotypes of annual sowthistle (Sonchus oleraceus) to mungbean density. Weed Sci. 2019, 67, 642–648. [Google Scholar] [CrossRef]
- Song, J.-S.; Kim, J.W.; Im, J.H.; Lee, K.J.; Lee, B.W.; Kim, D.S. The effects of single- and multiple-weed interference on soybean yield in the far-eastern region of Russia. Weed Sci. 2017, 65, 371–380. [Google Scholar] [CrossRef]
- Manalil, S.; Werth, J.; Jackson, R.; Chauhan, B.S.; Preston, C. An assessment of weed flora 14 years after the introduction of glyphosate-tolerant cotton in Australia. Crop Past. Sci. 2017, 68, 773–780. [Google Scholar] [CrossRef]
- Thomas, G.; Felton, W.; Radford, B. Tillage and crop residue management. In Sustainable Crop Production in the Sub-Tropics: An Australian Perspective; Clarke, A.L., Wylie, P.B., Eds.; Queensland Department of Primary Industries: Roma, QLD, Australia, 1997; pp. 195–213. [Google Scholar]
- Webb, A.; Grundy, M.; Powell, B.; Littleboy, M. The Australian sub-tropical cereal belt: Soils, climate and agriculture. In Sustainable Crop Production in the Sub-Tropics: An Australian Perspective; Clarke, A.L., Wylie, P.B., Eds.; Queensland Department of Primary Industries: Roma, QLD, Australia, 1997; pp. 8–23. [Google Scholar]
- Widderick, M.; Sindel, B.; Walker, S. Distribution, importance and management of Sonchus oleraceus (common sowthistle) in the northern cropping region of Australia. In Proceedings of the 12th Australian Weeds Conference, Hobart, TAS, Australia, 12–16 September 1999; p. 198. [Google Scholar]
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 1 June 2020).
- Manalil, S.; Coast, O.; Werth, J.; Chauhan, B.S. Weed management in cotton (Gossypium hirsutum L.) through weed-crop competition: A review. Crop Prot. 2017, 95, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.H.; Malone, J.M.; Boutsalis, P.; Shirley, N.; Preston, C. Temperature influences the level of glyphosate resistance in barnyardgrass (Echinochloa colona). Pest Manag. Sci. 2016, 72, 1031–1039. [Google Scholar] [CrossRef]
- Sarangi, D.; Sandell, L.D.; Kruger, G.R.; Knezevic, S.Z.; Irmak, S.; Jhala, A.J. Comparison of herbicide programs for season-long control of glyphosate-resistant common waterhemp (Amaranthus rudis) in soybean. Weed Technol. 2017, 31, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Harker, K.N.; Blackshaw, R.E. Influence of growth stage and broadleaf herbicides on tralkoxydim activity. Weed Sci. 1991, 39, 650–659. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Abugho, S.B. Effect of growth stage on the efficacy of postemergence herbicides on four weed species of direct-seeded rice. Sci. World J. 2012, 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Banks, P.A.; Robinson, E.L. The influence of straw mulch on the soil reception and persistence of metribuzin. Weed Sci. 1982, 30, 164–168. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Abugho, S.B. Interaction of rice residue and PRE herbicides on emergence and biomass of four weed species. Weed Technol. 2012, 26, 627–632. [Google Scholar] [CrossRef]
- Khalil, Y.; Flower, K.; Siddique, K.H.M.; Ward, P. Rainfall affects leaching of pre-emergent herbicide from wheat residue into the soil. PLoS ONE 2019, 14, e0210219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobli, A.; Rinwa, A.; Sahil; Chauhan, B.S. Effects of sorghum residue in presence of pre-emergence herbicides on emergence and biomass of Echinochloa colona and Chloris virgata. PLoS ONE 2020, 15, e0229817. [Google Scholar] [CrossRef] [PubMed]
- Genstat for Windows, 20th ed.; VSN International: Hemel Hempstead, UK, 2019.
- Shrestha, A.; Budhathoki, S.; Steinhauer, K. Temperature effects on glyphosate resistance in California populations of junglerice. Agron. J. 2018, 110, 1624–1626. [Google Scholar] [CrossRef]
- Matzrafi, M.; Brunharo, C.; Tehranchian, P.; Hanson, B.D.; Jasieniuk, M. Increased temperatures and elevated CO2 levels reduce the sensitivity of Conyza canadensis and Chenopodium album to glyphosate. Sci. Rep. 2019, 9, 2228. [Google Scholar] [CrossRef]
- Ali, H.H.; Kebaso, L.; Manalil, S.; Chauhan, B.S. Emergence and germination response of Sonchus oleraceus and Rapistrum rugosum to different temperatures and moisture stress regimes. Plant Species Biol. 2020, 35, 16–23. [Google Scholar] [CrossRef]
- VanGessel, M.J.; Scott, B.A.; Johnson, Q.R.; White-Hansen, S.E. Influence Of glyphosate-resistant horseweed (Conyza canadensis) growth stage on response to glyphosate applications. Weed Technol. 2009, 23, 49–53. [Google Scholar] [CrossRef]
- Kaur, S.; Sandell, L.D.; Lindquist, J.L.; Jhala, A.J. Glyphosate-resistant giant ragweed (Ambrosia trifida) control in glufosinate-resistant soybean. Weed Technol. 2014, 28, 569–577. [Google Scholar] [CrossRef]
- Landry, R.L.; Stephenson, D.O.; Woolam, B.C. Glufosinate rate and timing for control of glyphosate-resistant rhizomatous Johnsongrass (Sorghum halepense) in glufosinate-resistant soybean. Int. J. Agron. 2016, 8040235. [Google Scholar] [CrossRef] [Green Version]
- Steckel, G.J.; Wax, L.M.; Simmons, F.W.; Phillips, W.H. Glufosinate efficacy on annual weeds is influenced by rate and growth stage. Weed Technol. 1997, 11, 484–488. [Google Scholar] [CrossRef]
- Eubank, T.W.; Poston, D.H.; Nandula, V.K.; Koger, C.H.; Shaw, D.R.; Reynolds, D.B. Glyphosate-resistant horseweed (Conyza canadensis) control using glyphosate-, paraquat-, and glufosinate-based herbicide programs. Weed Technol. 2008, 22, 16–21. [Google Scholar] [CrossRef]
- Yu, Q.; Cairns, A.; Powles, S.B. Paraquat resistance in a population of Lolium rigidum. Funct. Plant Biol. 2004, 31, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Odero, D.C. Response of ragweed parthenium (Parthenium hysterophorus) to saflufenacil and glyphosate. Weed Technol. 2012, 26, 443–448. [Google Scholar] [CrossRef]
- Budd, C.M.; Soltani, N.; Robinson, D.E.; Hooker, D.C.; Miller, R.T.; Sikkema, P.H. Efficacy of saflufenacil for control of glyphosate-resistant horseweed (Conyza canadensis) as affected by height, density, and time of day. Weed Technol. 2017, 65, 275–284. [Google Scholar] [CrossRef]
- Alletto, L.; Coquet, Y.; Bergheaud, V.; Benoit, P. Water pressure head and temperature impact on isoxaflutole degradation in crop residues and loamy surface soil under conventional and conservation tillage management. Chemosphere 2012, 88, 1043–1050. [Google Scholar] [CrossRef]
- Stephenson, D.O.; Bond, J.A. Evaluation of thiencarbazone-methyl- and isoxaflutole-based herbicide programs in corn. Weed Technol. 2012, 26, 37–42. [Google Scholar] [CrossRef]
- Nelson, E.A.; Penner, D. Sensitivity of selected crops to isoxaflutole in soil and irrigation water. Weed Technol. 2005, 19, 659–663. [Google Scholar] [CrossRef]
- Soni, N.; Leon, R.G.; Erickson, J.E.; Ferrell, J.A.; Silveira, M.L. Biochar decreases atrazine and pendimethalin preemergence herbicidal activity. Weed Technol. 2015, 29, 359–366. [Google Scholar] [CrossRef]
- Price, A.J.; Balkcom, K.S.; Duzy, L.M.; Kelton, J.A. Herbicide and cover crop residue integration for Amaranthus control in conservation agriculture cotton and implications for resistance management. Weed Technol. 2012, 26, 490–498. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trial | Dates | Temperature (°C) | Assigned Details | |||
|---|---|---|---|---|---|---|
| Planting | Spray | Observation | Min | Av | ||
| 1 | 14 Feb 2019 | 25 Mar 2019 | 22 Apr 2019 | 17.3 | 28.1 | High temperature—Run 1 |
| 2 | 14 May 2019 | 20 June 2019 | 17 Jul 2019 | 8.6 | 19.0 | Low temperature—Run 1 |
| 3 | 14 Aug 2019 | 11 Sep 2019 | 10 Oct 2019 | 9.0 | 23.7 | Low temperature—Run 2 |
| 4 | 14 Nov 2019 | 4 Dec 2019 | 2 Jan 2020 | 18.2 | 30.1 | High temperature—Run 2 |
| Experiment 2: Post-Emergence | Experiment 3: Pre-Emergence | ||
|---|---|---|---|
| Herbicides | Rates (g a.i. or a.e./ha) | Herbicides | Rates (g a.i./ha) |
| 2,4-D | 700 | Isoxaflutole | 75 |
| 2,4-D | 1050 | Isoxaflutole | 150 |
| 2,4-D + picloram | 75 | Pendimethalin | 910 |
| 2,4-D + picloram | 112.5 | Pendimethalin | 1820 |
| Bromoxynil | 280 | S-metolachor | 960 |
| Bromoxynil | 420 | S-metolachlor | 1920 |
| Fluroxypyr * | 66.7 | ||
| Fluroxypyr * | 100 | ||
| Glufosinate | 500 | ||
| Glufosinate | 750 | ||
| Glyphosate | 496 | ||
| Glyphosate | 741 | ||
| Metsulfuron | 2 | ||
| Metsulfuron | 3 | ||
| Paraquat | 400 | ||
| Paraquat | 600 | ||
| Saflufenacil ** | 15.9 | ||
| Saflufenacil ** | 23.8 | ||
| Herbicide | Rates (g a.i./ha) | Seedling Survival (%) | |||
|---|---|---|---|---|---|
| 4-Leaf Stage | 6-Leaf Stage | ||||
| GR | GS | GR | GS | ||
| Control | 100 | 100 | 100 | 100 | |
| 2,4-D | 700 | 100 | 92 | 100 | 100 |
| 2,4-D | 1050 | 83 | 29 | 100 | 100 |
| 2,4-D + picloram | 75 | 0 | 0 | 0 | 100 |
| 2,4-D + picloram | 112.5 | 0 | 0 | 0 | 78 |
| Bromoxynil | 280 | 0 | 0 | 100 | 100 |
| Bromoxynil | 420 | 0 | 0 | 100 | 100 |
| Fluroxypyr | 66.7 | 100 | 100 | 100 | 100 |
| Fluroxypyr | 100 | 100 | 100 | 100 | 100 |
| Glufosinate | 500 | 0 | 0 | 0 | 0 |
| Glufosinate | 750 | 0 | 0 | 0 | 0 |
| Glyphosate | 496 | 100 | 0 | 100 | 36 |
| Glyphosate | 741 | 54 | 0 | 100 | 88 |
| Metsulfuron | 2 | 100 | 100 | 100 | 100 |
| Metsulfuron | 3 | 96 | 92 | 100 | 100 |
| Paraquat | 400 | 0 | 0 | 0 | 0 |
| Paraquat | 600 | 0 | 0 | 0 | 0 |
| Saflufenacil | 15.9 | 0 | 0 | 49 | 71 |
| Saflufenacil | 23.8 | 0 | 0 | 89 | 64 |
| LSD | 10.8 | 13.8 | 16.6 | 18.5 | |
| Herbicide | Rates (g a.i./ha) | Biomass (g/plant) | |||
|---|---|---|---|---|---|
| 4-Leaf Stage | 6-Leaf Stage | ||||
| GR | GS | GR | GS | ||
| Control | 0.34 | 0.54 | 0.27 | 0.38 | |
| 2,4-D | 700 | 0.23 (34) | 0.16 (71) | 0.18 (34) | 0.21 (44) |
| 2,4-D | 1050 | 0.12 (66) | 0.05 (91) | 0.17 (39) | 0.31 (20) |
| 2,4-D + picloram | 75 | 0 (100) | 0 (100) | 0 (100) | 0.10 (75) |
| 2,4-D + picloram | 112.5 | 0 (100) | 0 (100) | 0 (100) | 0.21 (45) |
| Bromoxynil | 280 | 0 (100) | 0 (100) | 0.20 (27) | 0.13 (66) |
| Bromoxynil | 420 | 0 (100) | 0 (100) | 0.15 (46) | 0.12 (68) |
| Fluroxypyr | 66.7 | 0.22 (37) | 0.30 (44) | 0.22 (21) | 0.30 (21) |
| Fluroxypyr | 100 | 0.22 (35) | 0.34 (37) | 0.23 (16) | 0.40 (−4) |
| Glufosinate | 500 | 0 (100) | 0 (100) | 0 (100) | 0 (100) |
| Glufosinate | 750 | 0 (100) | 0 (100) | 0 (100) | 0 (100) |
| Glyphosate | 496 | 0.05 (87) | 0 (100) | 0.14 (48) | 0.04 (90) |
| Glyphosate | 741 | 0.03 (92) | 0 (100) | 0.20 (26) | 0.01 (97) |
| Metsulfuron | 2 | 0.37 (−8) | 0.07 (87) | 0.30 (−10) | 0.19 (52) |
| Metsulfuron | 3 | 0.47 (−36) | 0.07 (88) | 0.30 (−9) | 0.17 (56) |
| Paraquat | 400 | 0 (100) | 0 (100) | 0 (100) | 0 (100) |
| Paraquat | 600 | 0 (100) | 0 (100) | 0 (100) | 0 (100) |
| Saflufenacil | 15.9 | 0 (100) | 0 (100) | 0.08 (71) | 0.05 (88) |
| Saflufenacil | 23.8 | 0 (100) | 0 (100) | 0.05 (81) | 0.06 (84) |
| LSD | 0.058 | 0.087 | 0.084 | 0.092 | |
| Herbicide | Rate (g a.i./ha) | Seedling Emergence (%) | |||||
|---|---|---|---|---|---|---|---|
| GR | GS | ||||||
| 0 t/ha | 3 t/ha | 6 t/ha | 0 t/ha | 3 t/ha | 6 t/ha | ||
| Control | - | 82 | 63 | 70 | 75 | 87 | 87 |
| Isoxaflutole | 75 | 13 | 38 | 50 | 37 | 72 | 62 |
| Isoxaflutole | 150 | 5 | 17 | 22 | 27 | 17 | 22 |
| Pendimethalin | 910 | 75 | 75 | 65 | 78 | 75 | 72 |
| Pendimethalin | 1820 | 68 | 20 | 30 | 78 | 33 | 25 |
| S-metolachor | 960 | 63 | 62 | 70 | 87 | 80 | 77 |
| S-metolachlor | 1920 | 30 | 58 | 72 | 40 | 78 | 80 |
| LSD | 21 | 19 | |||||
| Herbicide | Rate (g a.i./ha) | Biomass (g/plant) | |||||
|---|---|---|---|---|---|---|---|
| GR | GS | ||||||
| 0 t/ha | 3 t/ha | 6 t/ha | 0 t/ha | 3 t/ha | 6 t/ha | ||
| Control | - | 0.0400 | 0.0466 | 0.0285 | 0.0159 | 0.0211 | 0.0302 |
| Isoxaflutole | 75 | 0.0006 (99) | 0.0005 (99) | 0.0035 (88) | 0.0007 (96) | 0.0025 (88) | 0.0056 (81) |
| Isoxaflutole | 150 | 0.0001 (100) | 0.0067 (86) | 0.0012 (96) | 0.0016 (90) | 0.0001 (100) | 0.0001 (100) |
| Pendimethalin | 910 | 0.0074 (82) | 0.0038 (92) | 0.0028 (90) | 0.0283 (−78) | 0.0229 (−8) | 0.0210 (30) |
| Pendimethalin | 1820 | 0.0067 (83) | 0.0028 (94) | 0.0053 (81) | 0.0114 (28) | 0.0129 (39) | 0.0094 (69) |
| S-metolachor | 960 | 0.0115 (71) | 0.0239 (49) | 0.0219 (23) | 0.0180 (−13) | 0.0262 (−24) | 0.0223 (26) |
| S-metolachlor | 1920 | 0.0034 (92) | 0.0096 (79) | 0.0203 (29) | 0.0065 (59) | 0.0134 (34) | 0.0283 (6) |
| LSD | 0.0187 | 0.0106 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chauhan, B.S.; Jha, P. Glyphosate Resistance in Sonchus oleraceus and Alternative Herbicide Options for Its Control in Southeast Australia. Sustainability 2020, 12, 8311. https://doi.org/10.3390/su12208311
Chauhan BS, Jha P. Glyphosate Resistance in Sonchus oleraceus and Alternative Herbicide Options for Its Control in Southeast Australia. Sustainability. 2020; 12(20):8311. https://doi.org/10.3390/su12208311
Chicago/Turabian StyleChauhan, Bhagirath S., and Prashant Jha. 2020. "Glyphosate Resistance in Sonchus oleraceus and Alternative Herbicide Options for Its Control in Southeast Australia" Sustainability 12, no. 20: 8311. https://doi.org/10.3390/su12208311
APA StyleChauhan, B. S., & Jha, P. (2020). Glyphosate Resistance in Sonchus oleraceus and Alternative Herbicide Options for Its Control in Southeast Australia. Sustainability, 12(20), 8311. https://doi.org/10.3390/su12208311