A Review on Practical Application and Potentials of Phytohormone-Producing Plant Growth-Promoting Rhizobacteria for Inducing Heavy Metal Tolerance in Crops


 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Heavy Metals as Inorganic Pollutants
3. Plant Response to Heavy Metal Stress
4. Heavy Metal Stress Mitigation Strategies
4.1. Exogenous Application of Phytohormones
4.2. Phytohormone Producing Microbes
5. Applications of Phytohormone-Producing Bacteria for Inducing Heavy Metal Stress Tolerance in Plants
6. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ensink, J.H.J.; Simmons, R.W.; Van der Hoek, W. Wastewater use in Pakistan: The case of Haroonabad and Faisalabad. In Wastewater Use in Irrigated Agriculture Confronting the Livelihood and Environmental Realities; Scott, C.A., Faruqui, N.I., Raschid, L., Eds.; CAB International: Wallingford, UK, 2004; pp. 91–99. [Google Scholar]
- Nauman, M.; Khalid, S.K. Heavy metals contamination of soils in response to wastewater irrigation in Rawalpindi region. Pak. J. Agric. Sci. 2010, 47, 215–224. [Google Scholar]
- Bashir, M.A.; Naveed, M.; Ahmad, Z.; Gao, B.; Mustafa, A.; Núñez-Delgado, A. Combined application of biochar and sulfur regulated growth, physiological, antioxidant responses and Cr removal capacity of maize (Zea mays L.) in tannery polluted soils. J. Environ. Manag. 2020, 259, 110051. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Du, B.; Liu, H.; Cui, H.; Zhang, W.; Fan, X.; Zhou, J. The bioavailability and contribution of the newly deposited heavy metals (copper and lead) from atmosphere to rice (Oryza sativa L.). J. Hazard. Mater. 2020, 384, 121285. [Google Scholar] [CrossRef] [PubMed]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Malik, Z.; Parveen, A.; Huang, L.; Riaz, M.; Bashir, S.; Mustafa, A.; Abbasi, G.H.; Xue, B.; Ali, U. Ameliorative effects of biochar on rapeseed (Brassica napus L.) growth and heavy metal immobilization in soil irrigated with untreated wastewater. J. Plant Growth Regul. 2020, 39, 266–281. [Google Scholar] [CrossRef]
- Mapanda, F.; Mangwayana, E.N.; Nyamangara, J.; Giller, K.E. The effect of long term irrigation using waste water on heavy metal content of soil under vegetables in Harare, Zimbabwe. Agric. Ecosyst. Environ. 2005, 107, 151–165. [Google Scholar] [CrossRef]
- Farid, G.; Sarwar, N.; Saifullah, A.A.; Ghafoor, A. Heavy metals (Cd, Ni and Pb) contamination of soils, plants and waters in Madina town of Faisalabad metropolitan and preparation of GIS based maps. ACST 2015, 4, 199. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Luo, S.; Chen, J.; Xiao, X.; Chen, L.; Zeng, G.; Liu, C.; He, Y. Effect of endophyte-infection on growth parameters and cd-induced phytotoxicity of Cd-hyperaccumulator Solanum nigrum L. Chemosphere 2012, 89, 743–750. [Google Scholar] [CrossRef]
- Saeed, Z.; Naveed, M.; Imran, M.; Bashir, M.A.; Sattar, A.; Mustafa, A.; Hussain, A.; Xu, M. Combined use of Enterobacter sp. MN17 and zeolite reverts the adverse effects of cadmium on growth, physiology and antioxidant activity of Brassica napus. PLoS ONE 2019, 14, e0213016. [Google Scholar] [CrossRef] [Green Version]
- Sabir, A.; Naveed, M.; Bashir, M.A.; Hussain, A.; Mustafa, A.; Zahir, Z.A.; Kamran, M.; Ditta, A.; Núñez-Delgado, A.; Saeed, Q.; et al. Cadmium mediated phytotoxic impacts in Brassica napus: Managing growth, physiological and oxidative disturbances through combined use of biochar and Enterobacter sp. MN17. J. Environ. Manag. 2020, 265, 110522. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Mustafa, A.; Majeed, S.; Naseem, Z.; Saeed, Q.; Khan, A.; Nawaz, A.; Baig, K.S.; Chen, J.T. Enhancing Cadmium Tolerance and Pea Plant Health through Enterobacter sp. MN17 Inoculation Together with Biochar and Gravel Sand. Plants 2020, 9, 530. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.A.; Popova, L.P. Functions and toxicity of cadmium in plants: Recent advances and future prospects. Turk. J. Bot. 2013, 37, 1–13. [Google Scholar]
- Faiz, Y.; Tufail, M.; Javed, M.T.; Chauhadry, M.; Siddique, N. Road dust pollution of Cd, Cu, Ni, Pb, and Zn along Islamabad expressway, Pakistan. Microchem. J. 2009, 92, 186–192. [Google Scholar] [CrossRef]
- Nawaz, A.; Khurshid, K.; Arif, M.S.; Ranjha, A.M. Accumulation of heavy metals in soil and rice plants (Oryza sativa L) irrigated with industrial effluents. Int. J. Agric. Biol. 2006, 8, 391–393. [Google Scholar]
- Karishna, A.K.; Govil, P.K. Heavy metal distribution and contamination in soils of Thane-Belapur industrial development area, Mumbai, Western India. Environ. Geol. 2005, 47, 1054–1061. [Google Scholar] [CrossRef]
- Fagliano, J.A.; Savrin, J.; Udasin, I.; Gochfeld, M. Community exposure and medical screening near chromium waste sites in New Jersey. Regul. Toxicol. Pharm. 1997, 26, S13–S22. [Google Scholar] [CrossRef]
- DalCorso, G.; Farinati, S.; Furini, A. Regulatory networks of cadmium stress in plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar] [CrossRef]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The Good, the Bad, and the Ugly of Rhizosphere Microbiome. In Probiotics and Plant Health; Springer: Singapore, 2017; pp. 253–290. [Google Scholar]
- Gill, S.S.; Tuteja, N. Cadmium stress tolerance in crop plants: Probing the role of sulfur. Plant Signal. Behav. 2011, 6, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Ghani, A. Effect of chromium toxicity on growth, chlorophyll and some mineral nutrients of Brassica juncea L. Egypt. Acad. J. Biol. Sci. 2011, 2, 9–15. [Google Scholar]
- Eick, M.J.; Peak, J.D.; Brady, P.V.; Pesek, J.D. Kinetics of lead adsorption/desorption on goethite: Residence time effect. Soil Sci. 1999, 164, 28–39. [Google Scholar] [CrossRef]
- Salam, A.; Bashir, S.; Khan, I.; Hussain, Q.; Gao, R.; Hu, H. Biochar induced Pb and Cu immobilization, Phytoavailability attenuation in Chinese cabbage, and improved biochemical properties in naturally co-contaminated soil. J. Soils Sediments 2019, 19, 2381–2392. [Google Scholar] [CrossRef]
- Hou, D.; Ding, Z.; Li, G.; Wu, L.; Hu, P.; Guo, G.; Wang, X.; Ma, Y.; O’Connor, D.; Wang, X. A sustainability assessment framework for agricultural land remediation in China. Land Degrad. Dev. 2017, 29, 4. [Google Scholar] [CrossRef]
- Quartacci, M.F.; Argilla, A.; Baker, A.J.M.; Navari-Izzo, F. Phytoextraction of metals from a multiply contaminated soil by Indian mustard. Chemosphere 2006, 63, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Barcelo, J.; Poschenrieder, C. Phytoremediation: Principles and perspectives. Contrib. Sci. 2003, 2, 333–344. [Google Scholar]
- Tiwari, S.; Lata, C. Heavy metal stress, signaling, and tolerance due to plant-associated microbes: An overview. Front. Plant Sci. 2018, 9, 452. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, K.; Mitra, S.; Sarkar, A.; Soren, T.; Maiti, T.K. Characterization of cadmium-resistant Klebsiella pneumoniae MCC 3091 promoted rice seedling growth by alleviating phytotoxicity of cadmium. Env. Sci. Pollut. Res. 2017, 24, 24419–24437. [Google Scholar] [CrossRef]
- Ahmad, M.; Naseer, I.; Hussain, A.; Zahid, M.M.; Mustafa, A.; Hilger, T.H.; Ahmad, Z.Z.; Minggang, X. Appraising Endophyte–Plant Symbiosis for Improved Growth, Nodulation, Nitrogen Fixation and Abiotic Stress Tolerance: An Experimental Investigation with Chickpea (Cicer arietinum L.). Agronomy 2019, 9, 621. [Google Scholar] [CrossRef] [Green Version]
- Naveed, M.; Mustafa, A.; Azhar, S.Q.T.A.; Kamran, M.; Ahmad, Z.Z.; Núñez-Delgado, A. Burkholderia phytofirmans PsJN and tree twigs derived biochar together retrieved Pb-induced growth, physiological and biochemical disturbances by minimizing its uptake and translocation in mung bean (Vigna radiata L.). Environ. Sci. Pollut. Res. 2020, 257, 109974. [Google Scholar]
- Pandey, S.; Ghosh, P.K.; Ghosh, S.; De, T.K.; Maiti, T.K. Role of heavy metal resistant Ochrobactrum sp. and Bacillus spp. strains in bioremediation of a rice cultivar and their PGPR like activities. J. Microbiol. 2013, 51, 11–17. [Google Scholar] [CrossRef]
- Guo, J.; Chi, J. Effect of Cd-tolerant plant growth-promoting rhizobium on plant growth and Cd uptake by Lolium multiflorum lam. and Glycine max (L.) merr. in Cd-contaminated soil. Plant Soil. 2014, 375, 205–214. [Google Scholar] [CrossRef]
- Moreira, H.; Marques, A.P.G.C.; Franco, A.R.; Rangel, A.O.; Castro, P.M. Phytomanagement of Cd-contaminated soils using maize (Zea mays L.) assisted by plant growth-promoting rhizobacteria. Environ. Sci. Pollut. Res. 2014, 21, 9742–9753. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Akhtar, M.J.; Zahir, Z.A.; Naveed, M.; Mitter, B.; Sessitsch, A. Cadmium-tolerant bacteria induce metal stress tolerance in cereals. Environ. Sci. Pollut. Res. 2014, 21, 11054–11065. [Google Scholar] [CrossRef]
- Cocozza, C.; Trupiano, D.; Lustrato, G.; Alfano, G.; Vitullo, D.; Falasca, A.; Lomaglio, T.; De Felice, V.; Lima, G.; Ranalli, G.; et al. Challenging synergistic activity of poplar-bacteria association for the Cd phytostabilization. Environ. Sci. Pollut. Res. 2015, 22, 19546–19561. [Google Scholar]
- Singh, R.; Pathak, B.; Fulekar, M.H. Characterization of PGP traits by heavy metals tolerant Pseudomonas putida and Bacillus safensis strain isolated from rhizospheric zone of weed (Phyllanthus urinaria) and its efficiency in Cd and Pb removal. IJCMAS 2015, 4, 954–975. [Google Scholar]
- Prapagdee, B.; Khonsue, N. Bacterial-assisted cadmium phytoremediation by Ocimum gratissimum L. in polluted agricultural soil: A field trial experiment. Int. J. Environ. 2015, 12, 3843–3852. [Google Scholar] [CrossRef] [Green Version]
- Pandey, N.; Bhatt, R. Role of soil associated Exiguobacterium in reducing arsenic toxicity and promoting plant growth in Vigna radiata. Eur. J. Soil Biol. 2016, 75, 142–150. [Google Scholar] [CrossRef]
- Kartik, V.P.; Jinal, H.N.; Amaresan, N. Characterization of cadmium-resistant bacteria for its potential in promoting plant growth and cadmium accumulation in Sesbania bispinosa root. Int. J. Phytoremediation 2016, 18, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chao, Y.; Li, Y.; Lin, Q.; Bai, J.; Tang, L.; Wang, S.; Ying, R.; Qiu, R. Survival strategies of the plant-associated bacterium Enterobacter sp. strain EG16 under cadmium stress. Appl. Environ. Microbiol. 2016, 82, 1734–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, I.; Akhtar, M.J.; Asghar, H.N.; Ghafoor, U.; Shahid, M. Differential effects of plant growth-promoting rhizobacteria on maize growth and cadmium uptake. J. Plant Growth Regul. 2016, 35, 303–315. [Google Scholar] [CrossRef]
- Li, W.C.; Ye, Z.H.; Wong, M.H. Metal mobilization and production of short-chain organic acids by rhizosphere bacteria associated with a Cd/Zn hyperaccumulating plant Sedum alfredii. Plant Soil 2010, 326, 453–467. [Google Scholar] [CrossRef]
- Hoberg, E.; Marschner, P.; Lieberei, R. Organic acid exudation and pH changes by Gordonia sp. and Pseudomonas fluorescens grown with P adsorbed to goethite. J. Microbiol. Res. 2005, 160, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, N.M.; Vedaraman, N. Remediation of soil contaminated with copper using rhamnolipids produced from Pseudomonas aeruginosa MTCC 2297 using waste frying rice bran oil. Ann. Microbiol. 2012, 62, 85–91. [Google Scholar] [CrossRef]
- Ashraf, S.; Naveed, M.; Afzal, M.; Seleiman, M.F.; Al-Suhaibani, N.A.; Zahir, Z.A.; Mustafa, A.; Refay, Y.; Alhammad, B.A.; Ashraf, S.; et al. Unveiling the potential of novel macrophytes for the treatment of tannery effluent in vertical flow pilot constructed wetlands. Water 2020, 2, 549. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, H.; Jaleel, F.; Nisar, M.F. Analyzing the status of heavy metals in irrigation water in suburban areas of Bahawalpur city, Pakistan. AEJAES 2014, 14, 732–738. [Google Scholar]
- Musa, J.J.; Ode, O.G.; Anijofor, S.C.; Adewumi, J.K. Quality evaluation of household wastewater for irrigation. J. Appl. Sci. Environ. Manag. 2011, 15, 431–437. [Google Scholar]
- Iqbal, H.H.; Taseer, R.; Anwar, S.; Mumtaz, M.; Qadir, A.; Shahid, N. Human health risk assessment: Heavy metal contamination of vegetables in Bahawalpur, Pakistan. Bull. Environ. Stud. 2016, 1, 10–17. [Google Scholar]
- Salomons, W.; Forstner, U.; Mader, P. Heavy Metals: Problems and Solutions; Springer: Berlin, Germany, 1995. [Google Scholar]
- Boyd, R.S. Heavy metal pollutants and chemical ecology: Exploring new frontiers. J. Chem. Ecol. 2010, 36, 46–58. [Google Scholar] [CrossRef]
- Herawati, N.; Suzuki, S.; Hayashi, K.; Rivai, I.F.; Koyoma, H. Cadmium, copper and zinc levels in rice and soil of Japan, Indonesia and China by soil type. Bull. Environ. Contam. Toxicol. 2000, 64, 33–39. [Google Scholar] [CrossRef]
- He, Z.L.; Yang, X.E.; Stoffella, P.J. Trace elements in agro ecosystems and impacts on the environment. J. Trace Elem. Med. Biol. 2005, 19, 125–140. [Google Scholar] [CrossRef]
- Athar, M.; Vohora, S.B. Heavy Metals and Environment; New Age International Limited: New Delhi, India, 2001. [Google Scholar]
- Lepp, N.W. Effect of Heavy Metal Pollution on Plants: Metals in the Environment; Pollution Monitoring Series; Applied Science Publishers, Department of Biology: Liverpool, UK, 2012. [Google Scholar]
- Masindi, V.; Muedi, K.L. Environmental Contamination by Heavy Metals. In Heavy Metals; El-Din, H., Saleh, M., Aglan, R.F., Eds.; Intech Open: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hauser, M.T. Molecular basis of natural variation and environmental control of trichome patterning. Front. Plant Sci. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.S.; Dietz, K.J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalvi, A.A.; Bhalerao, S.A. Response of plants towards heavy metal toxicity: An overview of avoidance, tolerance and uptake mechanism. Ann. Plant Sci. 2013, 2, 362–368. [Google Scholar]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 1–12. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Germaine, K.; Bourke, P.; Malone, R. AFLP analysis of genetic diversity and phylogenetic relationships of Brassica oleracea in Ireland. C. R. Biol. 2016, 339, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Vwioko, E.; Adinkwu, O.; El-Esawi, M.A. Comparative Physiological, Biochemical and Genetic Responses to Prolonged Waterlogging Stress in Okra and Maize Given Exogenous Ethylene Priming. Front. Physiol. 2017, 8, 632. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A.; Witczak, J.; Ahmad, M. Analysis of Genetic Variation and Enhancement of Salt Tolerance in French Pea (Pisum Sativum, L.). Int. J. Mol. Sci. 2018, 19, 2433. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 Transcription Factor Enhances Heat and Drought Stress Tolerance in Wheat (Triticum aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A. Genetic diversity and evolution of Brassica genetic resources: From morphology to novel genomic technologies—A review. Plant Genet. Resour. 2017, 15, 388–399. [Google Scholar] [CrossRef]
- John, R.; Ahmad, P.; Gadgil, K.; Sharma, S. Heavy metal toxicity: Effect on plant growth, biochemical parameters and metal accumulation by Brassica juncea L. Int. J. Plant Prod. 2009, 3, 65–76. [Google Scholar]
- Hameed, A.; Rasool, S.; Azooz, M.M.; Hossain, M.A.; Ahanger, M.A.; Ahmad, P. Heavy Metal Stress: Plant Responses and Signaling. In Plant Metal Interaction: Emerging Remediation Techniques; Ahmad, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 557–583. [Google Scholar]
- Chen, L.; Guo, Y.; Yang, L.; Wang, Q. Synergistic defensive mechanism of phytochelatins and antioxidative enzymes in Brassica chinensis L. against Cd stress. Chin. Sci. Bull. 2008, 53, 1503–1511. [Google Scholar] [CrossRef]
- Bundy, J.G.; Kille, P.; Liebeke, M.; Spurgeon, D.J. Metallothioneins may not be enough-the role of phytochelatins in invertebrate metal detoxification. Environ. Sci. Technol. 2014, 48, 885–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagorchev, L.; Seal, C.E.; Kranner, I.; Odjakova, M. A central role for thiols in plant tolerance to abiotic stress. Int. J. Mol. Sci. 2013, 14, 7405–7432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saba, H.; Jyoti, P.; Neha, S. Mycorrhizae and phytochelators as remedy in heavy metal contaminated land remediation. Int. J. Environ. Sci. 2013, 2, 74–78. [Google Scholar]
- Thangavel, P.; Long, S.; Minocha, R. Changes in phytochelatins and their biosynthetic intermediates in red spruce (Picea rubens Sarg.) cell suspension cultures under cadmium and zinc stress. Plant Cell Tissue Organ Cult. 2007, 88, 201–216. [Google Scholar] [CrossRef]
- Yurekli, F.; Kucukbay, Z. Synthesis of phytochelatins in Helianthus annuus is enhanced by cadmium nitrate. Acta Bot Croat. 2003, 62, 21–25. [Google Scholar]
- Heiss, S.; Wachter, A.; Bogs, J.; Cobbett, C.; Rausch, T. Phytochelatin synthase (PCS) protein is induced in Brassica juncea leaves after prolonged Cd exposure. J. Exp. Bot. 2003, 54, 1833–1839. [Google Scholar] [CrossRef]
- Du, J.; Yang, J.L.; Li, C.H. Advances in metallotionein studies in forest trees. Plant Omics 2012, 5, 46–51. [Google Scholar]
- Yang, Z.; Chu, C. Towards Understanding Plant Response to Heavy Metal Stress. In Abiotic Stress in Plants-Mechanisms and Adaptations; InTech: Shanghai, China, 2011; pp. 59–78. [Google Scholar]
- Leszczyszyn, O.I.; Imam, H.T.; Blindauer, C.A. Diversity and distribution of plant metallothioneins: A review of structure, properties and functions. METAJS 2013, 5, 1146–1169. [Google Scholar] [CrossRef]
- Gohari, M.; Habib-Zadeh, A.R.; Khayat, M. Assessing the intensity of tolerance to lead and its effect on amount of protein and proline in root and aerial parts of two varieties of rape seed (Brassica napus L.). JBASR 2012, 2, 935–938. [Google Scholar]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Saharan, B.S.; Nehra, V. Plant growth promoting rhizobacteria: A critical review. LSMR 2011, 21, 1–30. [Google Scholar]
- Arshad, M.; Frankenberger, W.T.J. Ethylene: Agricultural Sources and Applications; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002. [Google Scholar]
- Ahmad, P.; Nabi, G.; Ashraf, M. Cadmium-induced oxidative damage in mustard [Brassica juncea (L.) Czern. & Coss.] plants can be alleviated by salicylic acid. S. Afr. J. Bot. 2011, 77, 36–44. [Google Scholar]
- Vardhini, B.V.; Anjum, N.A. Brassinosteroids make plant life easier under abiotic stresses mainly by modulating major components of antioxidant defense system. Front. Environ. Sci. 2015, 2, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dar, T.A.; Uddin, M.; Khan, M.M.A.; Hakeem, K.R.; Jaleel, H. Jasmonates counter plant stress: A review. Environ. Exp. Bot. 2015, 115, 49–57. [Google Scholar] [CrossRef]
- Ayvaz, M.; Koyuncu, M.; Guven, A.; Fagerstedt, K.V. Does boron affect hormone levels of barley cultivars? Eurasia J. Biosci. 2012, 6, 113–120. [Google Scholar] [CrossRef]
- Atici, O.; Agar, G.; Battal, P. Changes in phytohormone contents in chickpea seeds germinating under lead or zinc stress. Biol. Plant 2005, 49, 215–222. [Google Scholar] [CrossRef]
- Brien, J.A.O.; Benkova, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4, 451. [Google Scholar] [CrossRef] [Green Version]
- Pospisilova, J. Participation of phytohormones in the stomatal regulation of gas exchange during water stress. Biol. Plant 2003, 46, 491–506. [Google Scholar] [CrossRef]
- Hollenbach, B.; Schreiber, L.; Hartung, W.; Dietz, K.J. Cadmium leads to stimulated expression of the lipid transfer protein genes in barley: Implications for the involvement of lipid transfer proteins in wax assembly. Planta 1997, 203, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between plant hormones and HMs responses. Genet. Mol. 2017, 40, 373–386. [Google Scholar]
- Kazan, K. Auxin and the integration of environmental signals into plant root development. Ann. Bot. 2013, 112, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Hussain, S.; Bano, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.A.; Khan, F.; Chen, Y.T.; Wu, C.; et al. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: Consequences for changing environment. Environ. Sci. Pollut. Res. 2015, 22, 4907–4921. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Panda, S.K.; Bhattacharjee, M.K.; Dutta, S. Role of arbuscular mycorrhiza in heavy metal tolerance in plants: Prospects for phytoremediation. J. Phytol. 2010, 2, 16–27. [Google Scholar]
- Aloui, A.; Recorbet, G.; Robert, F.; Schoefs, B.; Bertrand, M.; Henry, C.; Gianinazzi-Pearson, V.; Dumas-Gaudot, E.; Aschi-Smiti, S. Arbuscular mycorrhizal symbiosis elicits shoot proteome changes that are modified during cadmium stress alleviation in Medicago truncatula. BMC Plant Biol. 2011, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Solanki, R.; Dhankhar, R. Biochemical changes and adaptive strategies of plants under heavy metal stress. Biologia 2011, 66, 195–204. [Google Scholar] [CrossRef]
- Chen, T.M.; Gokhale, J.; Shofer, S.; Kuschner, W.G. Outdoor air pollution: Nitrogen dioxide, sulfur dioxide, and carbon monoxide health effects. Am. J. Med. Sci. 2007, 333, 249–256. [Google Scholar] [CrossRef]
- Lajayer, A.B.; Ghorbanpour, M.; Nikabadi, S. HMs in contaminated environment: Destiny of secondary metabolite biosynthesis, oxidative status and phytoextraction in medicinal plants. Ecotoxicol. Environ. Saf. 2017, 145, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Khalid, S.; Abbas, G.; Shahid, N.; Nadeem, M.; Sabir, M.; Aslam, M.; Dumat, C. Heavy Metal Stress and Crop Productivity. In Crop Production and Global Environmental Issues; Hakeem, K.R., Ed.; Springer: Cham, Switzerland, 2015; 1025p. [Google Scholar]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Bradl, H.; Xenidis, A. Remediation techniques. Interface Sci. Technol. 2005, 6, 165–261. [Google Scholar]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Schnoor, J. Phytoremediation; Technology Evaluation Report TE-98-01; Groundwater Remediation Technologies Analysis Center: Pittsburgh, PA, USA, 1997. [Google Scholar]
- Bashar, K.K.; Tareq, M.Z.; Amin, M.R.; Honi, U.; Tahjib-Ul-Arif, M.; Sadat, M.A.; Hossen, Q.M.M. Phytohormone-mediated stomatal response, escape and quiescence strategies in plants under flooding stress. Agronomy 2019, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone priming: Regulator for heavy metal stress in plants. J. Plant Growth Regul. 2018, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Bhardwaj, R. Effects of 24-epibrassino- lide on growth and metal uptake in Brassica juncea L. under copper metal stress. Acta Physiol. Plant. 2007, 29, 259–263. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Bhardwaj, R.; Gupta, B.D.; Dutt, P.; Gupta, R.K.; Biondi, S.; Kanwar, M. Epibrassinolide induces changes in indole-3-acetic acid, abscisic acid and polyamine concentrations and enhances anti-oxidant potential of radish seedlings under copper stress. Plant Physiol. 2010, 140, 280–296. [Google Scholar]
- Yadav, P.; Kaur, R.; Kanwar, M.K.; Sharma, A.; Verma, V.; Sirhindi, G.; Bhardwaj, R. Castasterone confers copper stress tolerance by regulating anti-oxidant enzyme responses, anti-oxidants, and amino acid balance in B. juncea seedlings. Ecotoxicol. Environ. Saf. 2018, 147, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Hayat, S.; Ali, B.; Ahmad, A. Effect of 28-homobrassinolide treatment on nickel toxicity in Brassica juncea. Photosynthetica 2007, 45, 139–142. [Google Scholar] [CrossRef]
- Janeczko, A.; Kościelniak, J.; Pilipowicz, M.; Lukaszewska, S. Protection of winter rape photosystem 2 by 24-Epibrassinolide under cadmium stress. Photosynthetica 2005, 43, 293–298. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, anti-oxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Hasan, S.A.; Hayat, S.; Ali, B.; Ahmad, A. 28-Homo-brassinolide protects chickpea (Cicer arietinum) from cadmium toxicity by stimulating anti-oxidants. Environ. Pollut. 2008, 151, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Siddique, K.H.; Ahmad, P. Interactive effect of 24-epibrassinolide and silicon alleviates cadmium stress via the modulation of anti-oxidant defense and glyoxalase systems and macronutrient content in Pisum sativum L. seedlings. BMC Plant Biol. 2018, 18, 146. [Google Scholar] [CrossRef]
- Kaur, R.; Yadav, P.; Sharma, A.; Kumar, T.A.; Kumar, V.; Kaur, K.S.; Bhardwaj, R. Castasterone and citric acid treatment restores photosynthetic attributes in Brassica juncea L. under Cd(II) toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Interaction of 24-epibrassinolide and salicylic acid regulates pigment contents, anti-oxidative defense responses, and gene expression in Brassica juncea L. seedlings under Pb stress. Environ. Sci. Pollut. Res. 2018, 25, 15159–15173. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Sharma, A.; Kumar, V.; Kaur, P.; Bhardwaj, R. Synergistic effect of 24-epibrassinolide and salicylic acid on photosynthetic efficiency and gene expression in Brassica juncea L. under Pb stress. Turk. J. Biol. 2017, 41, 943–953. [Google Scholar] [CrossRef]
- Pavlu, J.; Novak, J.; Koukalova, V.; Luklova, M.; Brzobohaty, B.; Cerny, M. Cytokinin at the crossroads of abiotic stress signalling pathways. Int. J. Mol. Sci. 2018, 19, 2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassi, E.; Pouget, J.; Petruzzelli, G.; Barbafieri, M. The effects of exogenous plant growth regulators in the phytoextraction of HMs. Chemosphere 2008, 71, 66–73. [Google Scholar] [CrossRef]
- Piotrowska-Niczyporuk, A.; Bajguz, A.; Zambrzycka, E.; Godlewska-Żyłkiewicz, B. Phytohormones as regulators of HM biosorption and toxicity in green alga Chlorella vulgaris (Chlorophyceae). Plant Physiol. Biochem. 2012, 52, 52–65. [Google Scholar] [CrossRef]
- Cassina, L.; Tassi, E.; Morelli, E.; Giorgetti, L.; Remorini, D.; Chaney, R.L.; Barbafieri, M. Exogenous cytokinin treatments of an NI Hyper-Accumulator, Alyssum murale, grown in a serpentine soil: Implications for phytoextraction. Int. J. Phytoremediation 2011, 13, 90–101. [Google Scholar] [CrossRef]
- Jalmi, S.K.; Bhagat, P.K.; Verma, D.; Noryang, S.; Tayyeba, S.; Singh, K.; Sharma, D.; Sinha, A.K. Traversing the links between heavy metal stress and plant signaling. Front. Plant Sci. 2018, 9, 12. [Google Scholar] [CrossRef]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy metal stress and responses in plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Fassler, E.; Evangelou, M.W.; Robinson, B.H.; Schulin, R. Effects of indole-3-acetic acid (IAA) on sunflower growth and HM uptake in combination with ethylene diamine disuccinic acid (EDDS). Chemosphere 2010, 80, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Hadi, F.; Bano, A.; Fuller, M.P. The improved phytoextraction of lead (Pb) and the growth of maize (Zea mays L.): The role of plant growth regulators (GA3 and IAA) and EDTA alone and in combinations. Chemosphere 2010, 80, 457–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aderholt, M.; Vogelien, D.L.; Koether, M.; Greipsson, S. Phytoextraction of contaminated urban soils by Panicum virgatum L. enhanced with application of a plant growth regulator (BAP) and citric acid. Chemosphere 2017, 175, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Bashri, G.; Prasad, S.M. Indole acetic acid modulates changes in growth, chlorophyll a fluorescence and anti-oxidant potential of Trigonella foenumgraecum L. grown under cadmium stress. Acta Physiol. Plant. 2015, 37, 1745–1758. [Google Scholar] [CrossRef]
- Srivastava, S.; Verma, P.C.; Chaudhary, V.; Singh, N.; Abhilash, P.C.; Kumar, K.V.; Sharma, N.; Singh, N. Inoculation of arsenic-resistant Staphylococcus arlettae on growth and arsenic uptake in Brassica juncea (L.) Czern. Var. R-46. J. Hazard. Mater. 2013, 262, 1039–1047. [Google Scholar] [CrossRef]
- Gangwar, S.; Singh, V.P.; Srivastava, P.K.; Maurya, J.N. Modification of chromium (VI) phytotoxicity by exogenous gibberellic acid application in Pisum sativum (L.) seedlings. Acta Physiol. Plant. 2011, 33, 1385–1397. [Google Scholar] [CrossRef]
- Zhu, X.F.; Jiang, T.; Wang, Z.W.; Lei, G.J.; Shi, Y.Z.; Li, G.X.; Zheng, S.J. Gibberellic acid alleviates cadmium toxicity by reducing nitric oxide accumulation and expression of IRT1 in Arabidopsis thaliana. J. Hazard. Mater. 2012, 239–240, 302–307. [Google Scholar] [CrossRef]
- Hadi, F.; Ali, N.; Ahmad, A. Enhanced phytoremediation of cadmium-contaminated soil by Parthenium hysterophorus plant: Effect of gibberellic acid (Ga3) and synthetic chelator, alone and in combinations. Bioremediat. J. 2014, 18, 46–55. [Google Scholar] [CrossRef]
- Wang, B.; Wei, H.; Xue, Z.; Zhang, W.H. Gibberellins regulate iron deficiency-response by influencing iron transport and translocation in rice seedlings (Oryza sativa). Ann. Bot. 2017, 119, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Chen, J.; Li, X. Methyl jasmonate as modulator of Cd toxicity in Capsicum frutescens var. fasciculatum seedlings. Ecotoxicol. Environ. Saf. 2013, 98, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Per, T.S.; Khan, M.I.R.; Anjum, N.A.; Masood, A.; Hussain, S.J.; Khan, N.A. Jasmonates in plants under abiotic stresses: Crosstalk with other phytohormones matters. Environ. Exp. Bot. 2018, 145, 104–120. [Google Scholar] [CrossRef]
- Belkhadi, A.; Hediji, H.; Abbes, Z.; Nouairi, I.; Barhoumi, Z.; Zarrouk, M.; Chaïbi, W.; Djebali, W. Effects of exogenous salicylic acid pre-treatment on cadmium toxicity and leaf lipid content in Linum usitatissimum L. Ecotoxicol. Environ. Saf. 2010, 73, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, N.; Khavari-Nejad, R.A.; Fahimi, H.; Saadatmand, S.; Nejad-Sattari, T. Effects of exogenous salicylic acid and nitric oxide on lipid peroxidation and anti-oxidant enzyme activities in leaves of Brassica napus L. under nickel stress. Sci. Hortic. 2010, 126, 402–407. [Google Scholar] [CrossRef]
- Xu, L.L.; Fan, Z.Y.; Dong, Y.J.; Kong, J.; Bai, X.Y. Effects of exogenous salicylic acid and nitric oxide on physiological characteristics of two peanut cultivars under cadmium stress. Biol. Plant 2015, 59, 171–182. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Zhou, G. Response and adaptation of photosynthesis, respiration, and anti-oxidant systems to elevated CO2 with environmental stress in plants. Front. Plant Sci. 2015, 6, 701. [Google Scholar] [CrossRef] [Green Version]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Combined effect of 24-epibrassinolide and salicylic acid mitigates lead (Pb) toxicity by modulating various metabolites in Brassica juncea L. seedlings. Protoplasma 2018, 255, 11–24. [Google Scholar] [CrossRef]
- Kohli, K.S.; Handa, N.; Bali, S.; Arora, S.; Sharma, A.; Kaur, R.; Bhardwaj, R. Modulation of anti-oxidative defense expression and osmolyte content by co-application of 24-epibrassinolide and salicylic acid in Pb exposed Indian mustard plants. Ecotoxicol. Environ. Saf. 2018, 147, 382–393. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Guo, K.; Elbaz, A.A.; Yang, Z.M. Salicylic acid alleviates mercury toxicity by preventing oxidative stress in roots of Medicago sativa. Environ. Exp. Bot. 2009, 65, 27–34. [Google Scholar] [CrossRef]
- Drazic, G.; Mihailovic, N. Modification of cadmium toxicity in soybean seedlings by salicylic acid. Plant Sci. 2005, 168, 511–517. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Khan, N.A. Ethylene reverses photosynthetic inhibition by nickel and zinc in mustard through changes in PS II activity, photosynthetic nitrogen use efficiency, and anti-oxidant metabolism. Protoplasma 2014, 251, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Asgher, M.; Per, T.S.; Masood, A.; Fatma, M.; Khan, M.I. Ethylene potentiates sulfur-mediated reversal of cadmium inhibited photosynthetic responses in mustard. Front. Plant Sci. 2016, 7, 1628. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 2016, 7, 1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd-Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef]
- Mustafa, A.; Naveed, M.; Saeed, Q.; Ashraf, M.N.; Hussain, A.; Abbas, T.; Kamran, M.; Minggang, X. Application Potentials of Plant Growth Promoting Rhizobacteria and Fungi as an Alternative to Conventional Weed Control Methods. In Crop Production; IntechOpen: London, UK, 2019. [Google Scholar]
- Cho, S.T.; Chang, H.H.; Egamberdieva, D.; Kamilova, F.; Lugtenberg, B.; Kuo, C.H. Genome Analysis of Pseudomonas fluorescens PCL1751: A rhizobacterium that controls root diseases and alleviates salt stress for its plant host. PLoS ONE 2015, 10, e0140231. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Alikhani, H.A.; Hosseini, H.M. Indole-3-acetic acid (IAA) production trait, a useful screening to select endophytic and rhizosphere competent bacteria for rice growth promoting agents. MethodsX 2015, 2, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Sorty, A.M.; Meena, K.K.; Choudhary, K.; Bitla, U.M.; Minhas, P.S.; Krishnani, K.K. Effect of plant growth promoting bacteria associated with halophytic weed (Psoralea corylifolia L.) on germination and seedling growth of wheat under saline conditions. Appl. Biochem. Biotechnol. 2016, 180, 872–882. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Hussain, I.; Rasheed, R.; Iqbal, M.; Riaz, M.; Arif, M.S. Advances in microbe-assisted reclamation of heavy metal contaminated soils over the last decade: A review. J. Environ. Manag. 2017, 198, 132–143. [Google Scholar] [CrossRef]
- Carlos, M.H.J.; Stefani, P.V.Y.; Janette, A.M.; Melani, M.S.S.; Gabriela, P.O. Assessing the effects of heavy metals in ACC deaminase and IAA production on plant growth-promoting bacteria. Microbiol. Res. 2016, 188, 53–61. [Google Scholar] [CrossRef]
- Seneviratne, M.; Gunaratne, S.; Bandara, T.; Weerasundara, L.; Rajakaruna, N.; Seneviratne, G.; Vithanage, M. Plant growh promotion by Bradyrhizobium japonicum under heavy metal stress. S. Afr. J. Bot. 2016, 105, 19–24. [Google Scholar] [CrossRef]
- Hrynkiewicz, K.; Zloch, M.; Kowalkowski, T.; Baum, C.; Niedojadlo, K.; Buszewski, B. Strain-specific bioaccumulation and intracellular distribution of Cd+2 in bacteria isolated from the rhizosphere, ectomycorrhizae, and fruitbodies of ectomycorrhizal fungi. Environ. Sci. Pollut. Res. 2015, 22, 3055–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgroy, V.; Cassán, F.; Masciarelli, O.; Del Papa, M.F.; Lagares, A.; Luna, V. Isolation and characterization of endophytic plant growth-promoting (PGPB) or stress homeostasis-regulating (PSHB) bacteria associated to the halophyte Prosopis strombulifera. Appl. Microbiol. Biotechnol. 2009, 85, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.; Behrendt, U.; Abd-Allah, E.F.; Berg, G. Biochar treatment resulted in a combined effect on soybean growth promotion and a shift in plant growth promoting rhizobacteria. Front. Microbiol. 2016, 7, 209. [Google Scholar] [CrossRef] [Green Version]
- Mishra, J.; Singh, R.; Arora, N.K. Alleviation of heavy metal stress in plants and remediation of soil by rhizosphere microorganisms. Front. Microbiol. 2017, 8, 1706. [Google Scholar] [CrossRef] [PubMed]
- Janzen, R.A.; Rood, S.B.; Dormaar, J.F.; McGill, W.B. Azospirillum brasilense produces gibberellin in pure culture and on chemically-defined medium in co-culture on straw. Soil Biol. Biochem. 1992, 24, 1061–1064. [Google Scholar] [CrossRef]
- Gutierrez-Manero, F.G.; Ramos-Solano, B.; Probanza, A.; Mehouachi, J.; Tadeo, F.R.; Talon, M. The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol. Plant. 2001, 111, 206–211. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.S.; Meena, V.S.; Ramteke, P.W. Plant growth-promoting rhizobacteria: Strategies to improve abiotic stresses under sustainable agriculture. J. Plant Nutr. 2019, 42, 1402–1415. [Google Scholar] [CrossRef]
- Singh, V.; Maharshi, A.; Singh, D.P.; Upadhyay, R.S.; Sarma, B.K.; Singh, H.B. Role of Microbial Seed Priming and Microbial Phytohormone in Modulating Growth Promotion and Defense Responses in Plants. In Advances in Seed Priming; Rakshit, A., Singh, H., Eds.; Springer: Singapore, 2018; pp. 115–126. [Google Scholar]
- Mustapha, M.U.; Halimoon, N. Screening and isolation of heavy metal tolerant bacteria in industrial effluent. Procedia Environ. Sci. 2015, 30, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.K.; Khan, M.H.; Misra, S.; Dixit, V.K.; Khare, P.; Srivastava, S.; Chauhan, P.S. Characterisation of Pseudomonas spp. and Ochrobactrum sp. isolated from volcanic soil. Antonie Van Leeuwenhoek 2017, 110, 253–270. [Google Scholar] [CrossRef]
- Román-Ponce, B.; Reza-Vázquez, D.M.; Gutiérrez-Paredes, S.; María de Jesús, D.E.; Maldonado-Hernández, J.; Bahena-Osorio, Y.; Estrada-de los Santose, P.; Wang, T.; Vásquez-Murrieta, M.S. Plant growth-promoting traits in rhizobacteria of heavy metal-resistant plants and their effects on Brassica nigra seed germination. Pedosphere 2017, 27, 511–526. [Google Scholar] [CrossRef]
- Dąbrowska, G.; Hrynkiewicz, K.; Trejgell, A.; Baum, C. The effect of plant growth-promoting rhizobacteria on the phytoextraction of Cd and Zn by Brassica napus L. Int. J. Phytoremediation 2017, 19, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.F.; Xia, J.J. Improvement of rape (Brassica napus) plant growth and cadmium uptake by cadmium-resistant bacteria. Chemosphere 2006, 64, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Weyens, N.; Beckers, B.; Schellingen, K.; Ceulemans, R.; Croes, S.; Janssen, J.; Haenen, S.; Witters, N.; Vangronsveld, J. Plant-associated bacteria and their role in the success or failure of metal phytoextraction projects: First observations of a field-related experiment. Microb. Biotechnol. 2013, 6, 288–299. [Google Scholar] [CrossRef]
- Sinha, S.; Mukherjee, S.K. Cadmium-induced siderophore production by a high Cd resistant bacteria strain relieved Cd-toxicity in plants through root colonization. Curr. Microbiol. 2008, 56, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.I.A.; Barbosa, L.; Castro, P.M.L. Rhizobacteria isolated from a metal-polluted area enhance plant growth in zinc and cadmium-contaminated soil. Int. J. Environ. Sci. Technol. 2015, 12, 2127–2142. [Google Scholar] [CrossRef] [Green Version]
- Hassan, T.U.; Bano, A.; Naz, I. Alleviation of heavy metals toxicity by the application of plant growth promoting rhizobacteria and effects on wheat grown in saline sodic field. Int. J. Phytoremediation 2017, 19, 522–529. [Google Scholar] [CrossRef]
- Fatima, H.E.; Ahmed, A. Micro-remediation of chromium contaminated soils. Peer J. 2018. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Fan, J.; Xing, Y.; Xu, G.; Wang, H.; Deng, J.; Wang, Y.; Zhang, F.; Li, P.; Li, Z. The Effects of Mulch and Nitrogen Fertilizer on the Soil Environment of Crop Plants. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Wang, X.; Wang, G.; Guo, T.T.; Xing, Y.; Mo, F.; Wang, H.; Fan, J.; Zhang, F. Effects of plastic mulch and nitrogen fertilizer on the soil microbial community, enzymatic activity, and yield performance in a dryland maize cropping system. Eur. J. Soil Sci. 2020, 1–13. [Google Scholar] [CrossRef]
- Khan, W.U.; Ahmad, S.R.; Yasin, N.A.; Ali, A.; Ahmad, A.; Akram, W. Application of Bacillus megaterium MCR-8 improved phytoextraction and stress alleviation of nickel in Vinca rosea. Int. J. Phytoremediation 2017, 19, 813–824. [Google Scholar] [CrossRef]
- Hansda, A.; Kumar, V.; Anshumali. Cu-resistant Kocuria sp. CRB15: A potential PGPR isolated from the dry tailing of Rakha copper mine. 3 Biotech 2017, 7, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Alleviation of mercury toxicity in wheat by the interaction of mercury-tolerant plant growth-promoting rhizobacteria. J. Plant Growth Regul. 2016, 35, 1000–1012. [Google Scholar] [CrossRef]
- Arora, K.; Sharma, S.; Monti, A. Bio-remediation of Pb and Cd polluted soils by switch grass: A case study in India. Int. J. Phytoremediation 2016, 18, 704–709. [Google Scholar] [CrossRef]
- Ma, Y.; Oliviera, R.S.; Nai, F.; Rajkumar, M.; Luo, Y.; Rocha, I.; Freitas, H. The hyperaccumulator Sedum plumbizincicola harbors metal-resistant endophytic bacteria that improve its phytoextraction capacity in multi-metal contaminated soil. J. Environ. Manag. 2015, 156, 62–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adediran, G.A.; Ngwenya, B.T.; Mosselmans, J.F.W.; Heal, K.V.; Harvie, B.A. Mechanism behind bacteria induced plant growth promotion and Zn accumulation in Brassica juncea. J. Hazard. Mater. 2015, 283, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Luo, S.; Li, X.; Wan, Y.; Chen, J.; Liu, C. Interaction of Cd hyperaccumulator Solanum nigrum L. and functional endophyte Pseudomonas sp. Lk9 on soil heavy metals uptake. Soil Biol. Biochem. 2014, 68, 300–308. [Google Scholar] [CrossRef]
- He, H.; Ye, Z.; Yang, D.; Yan, J.; Xiao, L.; Zhong, T.; Yuan, M.; Cai, X.; Fang, Z.; Jing, Y. Characterization of endophytic Rahnella sp. JN6 from Polygonum pubescens and its potential in promoting growth and Cd, Pb, Zn uptake by Brassica napus. Chemosphere 2013, 90, 1960–1965. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K.; Suprasanna, P.; D’Souza, S.F. Identification and profiling of arsenic stress-induced microRNAs in Brassica juncea. J. Exp. Bot. 2013, 64, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Rajkumar, M.; Fritas, H. Inoculation of plant growth promoting bacterium Achromobacter xylosoxidans strain Ax10 for the improvement of copper phytoextraction by Brassica juncea. J. Environ. Manag. 2008, 90, 831–837. [Google Scholar] [CrossRef] [Green Version]


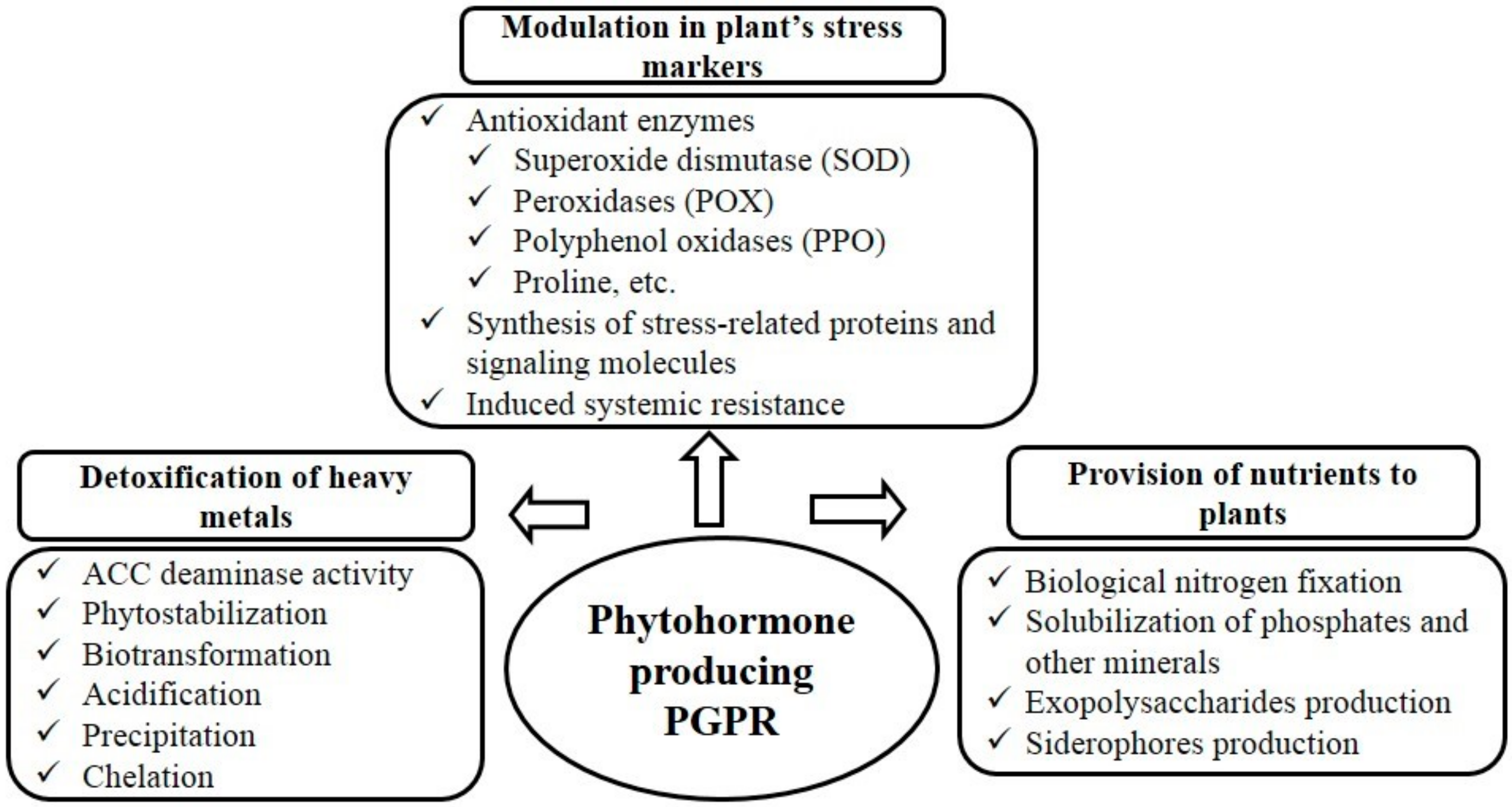{kind=link}
| Bacterial Strains | Plant Growth Promoting Traits | Plants | Heavy Metals | Effects on Plants and Metal Uptake | Reference |
|---|---|---|---|---|---|
| Enterobacter sp. | IAA production, Cd tolerance, siderophore production | Pea | Cd | Inoculation retrieved metal-induced growth disturbance by minimizing metal uptake in mung bean | [13] |
| Increase in growth parameters and xanthophyll, carotenoid, and chlorophyll content | |||||
| Bacillus sp. | Auxins production, phosphate solubilization | Lens culinaris | Cr | Inoculation reduced the negative effects of Cr on plants, improved plant growth parameters | [175] |
| Bacillus megaterium MCR−8 | IAA production, siderophore production, ACC deaminase activity, phosphate solubilization, Cd tolerance ability | Vinca rosea L. | Ni | Inoculation improved plant growth, proline contents and total soluble protein under Ni amended media | [176] |
| Enhanced the activity of antioxidant enzymes, i.e., APX, CAT, SOD, and POD under Ni stress | |||||
| Increased the Ni phytoextraction from soil | |||||
| Kocuria sp. CRB15 | IAA production, NH3 production, phosphate solubilization, tolerance to Cu stress | Brassica nigra | Cu | Improved root and shoot growth of Brassica nigra | [177] |
| Decrease in toxic effect of Cu on plants | |||||
| Klebsiella pneumoniae MCC 3091 | IAA production, siderophore production, ACC deaminase activity, phosphate solubilization, Cd tolerance ability | Oryza sativa | Cd | Significant improvement in seed germination and root and shoot growth under Cd stress | [29] |
| Improved the antioxidant SOD, MDA, and CAT activity | |||||
| Significantly improved the chlorophyll contents, total protein, total sugar, proline, α-amylase, and protease activity | |||||
| Reduced the ethylene production Cd uptake in rice seedlings | |||||
| Klebsiella pneumoniae (HG 3); | IAA production, siderophore production, NH3 production, exopolysaccharides secretion, ACC deaminase activity, phosphate solubilization, K and Zn solubilization, Hg-stress tolerance | Triticum aestivum | Hg | Inoculation significantly improved the relative water contents and root and shoot growth of wheat plants | [178] |
| Enterobacter ludwigii (HG 2) | Malondialdehyde content, electrolyte leakage, and proline contents were also improved under mercury stress | ||||
| Azospirillum | IAA production, tolerance to Pb and Cd, phosphate solubilization | Panicum virgatum | Cd, Pb | Inoculation increased the surface area, number of branches, root length, and root and shoot biomass | [179] |
| Neutralized the soil pH | |||||
| Prevented the Pb and Cd translocation to aerial parts | |||||
| Bacillus, Klebsiella, Leifsonia and Enterobacter | Auxin production, Cd-stress tolerance, phosphate solubilization, oxidase and catalase activity, EPS-production | Zea mays | Cd | Significant increase in shoot and root growth, dry biomass under Cd-stress | [42] |
| Inoculation significantly increased the membrane permeability and relative water contents in maize leaves | |||||
| Helped in Cd extraction and stabilization | |||||
| Pseudomonas putida (ATCC 39213) | IAA production, ACC deaminase activity | Eruca sativa | Cd | Inoculation was more effective in improving root and shoot growth and chlorophyll contents at lower levels, while efficiency decreased at higher | [7] |
| level, also lowered the proline contents in leaves | |||||
| Inoculation increased the Cd uptake by E. sativa plants | |||||
| Achromobacter sp. E4L5, Bacillus sp. E4S1, Bacillus sp. E1S2, Bacillus pumilus E2S2 and Stenotrophomonas sp. E1L | IAA production, ACC deaminase activity, siderophore production, phosphorus solubilization, tolerance to heavy metals | Sedum plumbizincicola | Zn, Cd, Pb | Heavy metal-tolerant bacterial strains effectively colonized the roots of S. plumbizincicola plants in multimetal contaminated soils. | [180] |
| Inoculation effectively improved the root and shoot growth and dry biomass of plants. | |||||
| Increase in water-extractable Cd and Zn concentration in soil. | |||||
| Enhanced the phytoextraction ability of S. plumbizincicola plants. | |||||
| Rhizobium leguminosarum | IAA production, siderophore production, phosphorus solubilization | Brassica juncea | Zn | Inoculation improved plant growth and Zn accumulation in Brassica juncea | [181] |
| Zinc stored in the roots as Zn cysteine and Zn phytate | |||||
| Induced metal resistance in plants through chelation of Zn | |||||
| Pseudomonas sp. LK9 | Ability to produce low molecular weight organic acids, siderophore and biosurfactants | Solanum nigrum | Cu, Zn, Cd | Inoculation improved the root and shoot growth and plant biomass of Solanum nigrum L. | [182] |
| Significantly improved the Cu, Zn, and Cd uptake in aerial parts | |||||
| Enhanced the P and Fe bioavailability in soil | |||||
| Inoculation improved the bioavailability of Cu, Zn, and Cd in soil | |||||
| Rahnella sp. JN6 | IAA production, ACC deaminase activity, phosphorus solubilization, tolerance to heavy metals | Brassica napus | Zn, Pb, Cd | Inoculation significantly improved the plant growth and dry weights of roots and shoots | [183] |
| Improved the uptake and accumulation of Zn, Pb, and Cd in roots and shoots | |||||
| Effectively colonized the tissue interior of rape plants | |||||
| Staphylococcus arlettae | IAA production, ACC deaminase activity, siderophore production | Brassica juncea | As | Inoculation with S. arlettae improved the growth and As uptake of Brassica juncea (L.) | [184] |
| Significantly improved the plant biomass, chlorophyll, and protein contents in plants | |||||
| Helped in phytostabilization of As by accumulating metal in roots | |||||
| Achromobacter xylosoxidans strain Ax10 | IAA production, ACC deaminase activity, phosphorus solubilization, tolerance to heavy metals | Brassica juncea | Cu | Inoculation with A. xylosoxidans significantly improved the root and shoot growth and fresh and dry weight of B. juncea plants under Cu stress | [184] |
| Improved the Cu uptake by plants and effectively sequestered the metal from soil |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazli, F.; Mustafa, A.; Ahmad, M.; Hussain, A.; Jamil, M.; Wang, X.; Shakeel, Q.; Imtiaz, M.; El-Esawi, M.A. A Review on Practical Application and Potentials of Phytohormone-Producing Plant Growth-Promoting Rhizobacteria for Inducing Heavy Metal Tolerance in Crops. Sustainability 2020, 12, 9056. https://doi.org/10.3390/su12219056
Nazli F, Mustafa A, Ahmad M, Hussain A, Jamil M, Wang X, Shakeel Q, Imtiaz M, El-Esawi MA. A Review on Practical Application and Potentials of Phytohormone-Producing Plant Growth-Promoting Rhizobacteria for Inducing Heavy Metal Tolerance in Crops. Sustainability. 2020; 12(21):9056. https://doi.org/10.3390/su12219056
Chicago/Turabian StyleNazli, Farheen, Adnan Mustafa, Maqshoof Ahmad, Azhar Hussain, Moazzam Jamil, Xiukang Wang, Qaiser Shakeel, Muhammad Imtiaz, and Mohamed A. El-Esawi. 2020. "A Review on Practical Application and Potentials of Phytohormone-Producing Plant Growth-Promoting Rhizobacteria for Inducing Heavy Metal Tolerance in Crops" Sustainability 12, no. 21: 9056. https://doi.org/10.3390/su12219056
APA StyleNazli, F., Mustafa, A., Ahmad, M., Hussain, A., Jamil, M., Wang, X., Shakeel, Q., Imtiaz, M., & El-Esawi, M. A. (2020). A Review on Practical Application and Potentials of Phytohormone-Producing Plant Growth-Promoting Rhizobacteria for Inducing Heavy Metal Tolerance in Crops. Sustainability, 12(21), 9056. https://doi.org/10.3390/su12219056