Vegetation Characteristics and Response to the Soil Properties of Three Medicinal Plant Communities in Altay Prefecture, China



Abstract
:1. Introduction
2. Materials and Methods
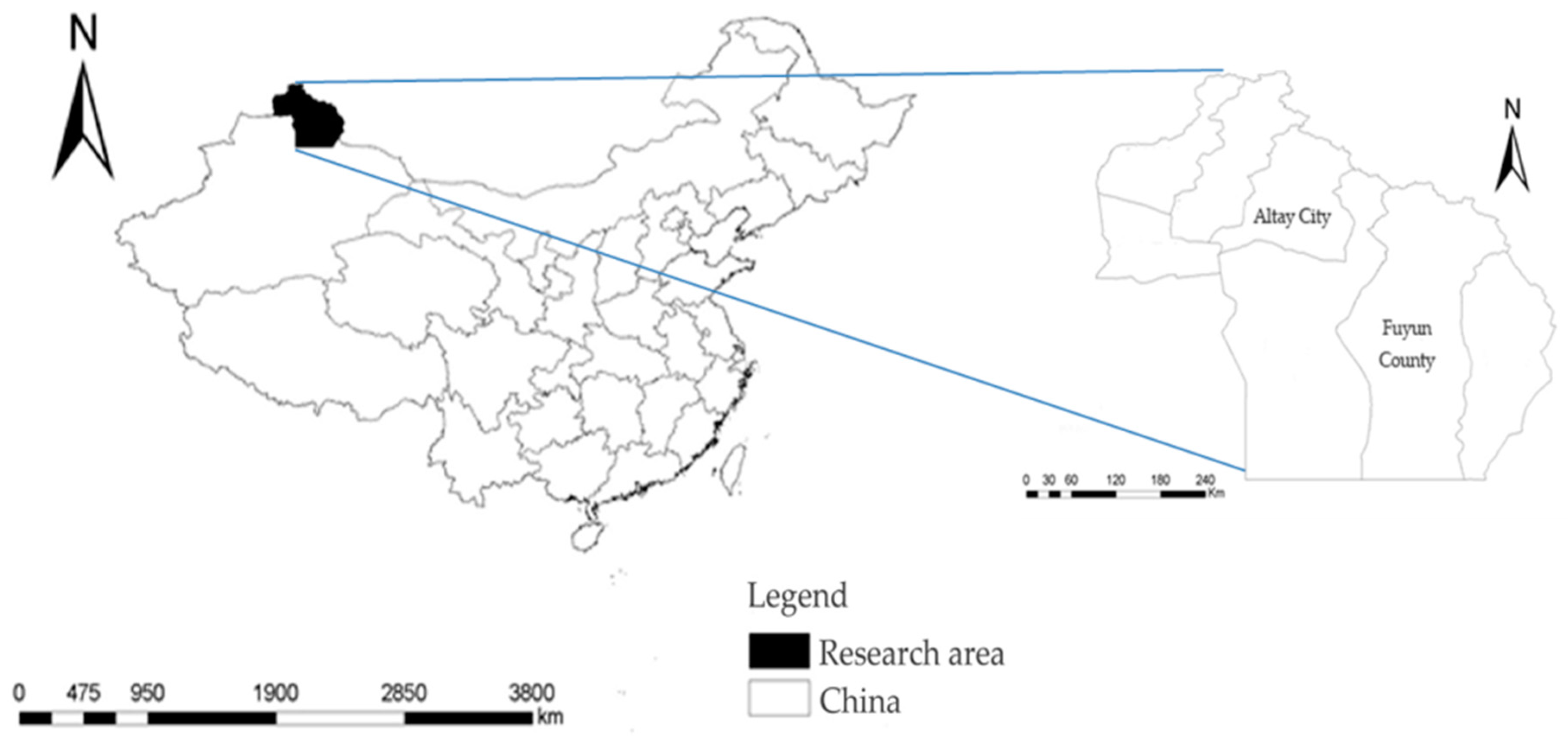
2.1. Study Area
2.2. Experimental Design
- (1)
- Plant species, abundance, height, crown width, herb layer coverage, and shrub layer coverage.
- (2)
- Accurate positioning, elevation, aspect, and soil type of each sample site.
2.3. Descriptive Statistical Analysis
2.4. Soil Collection and Processing
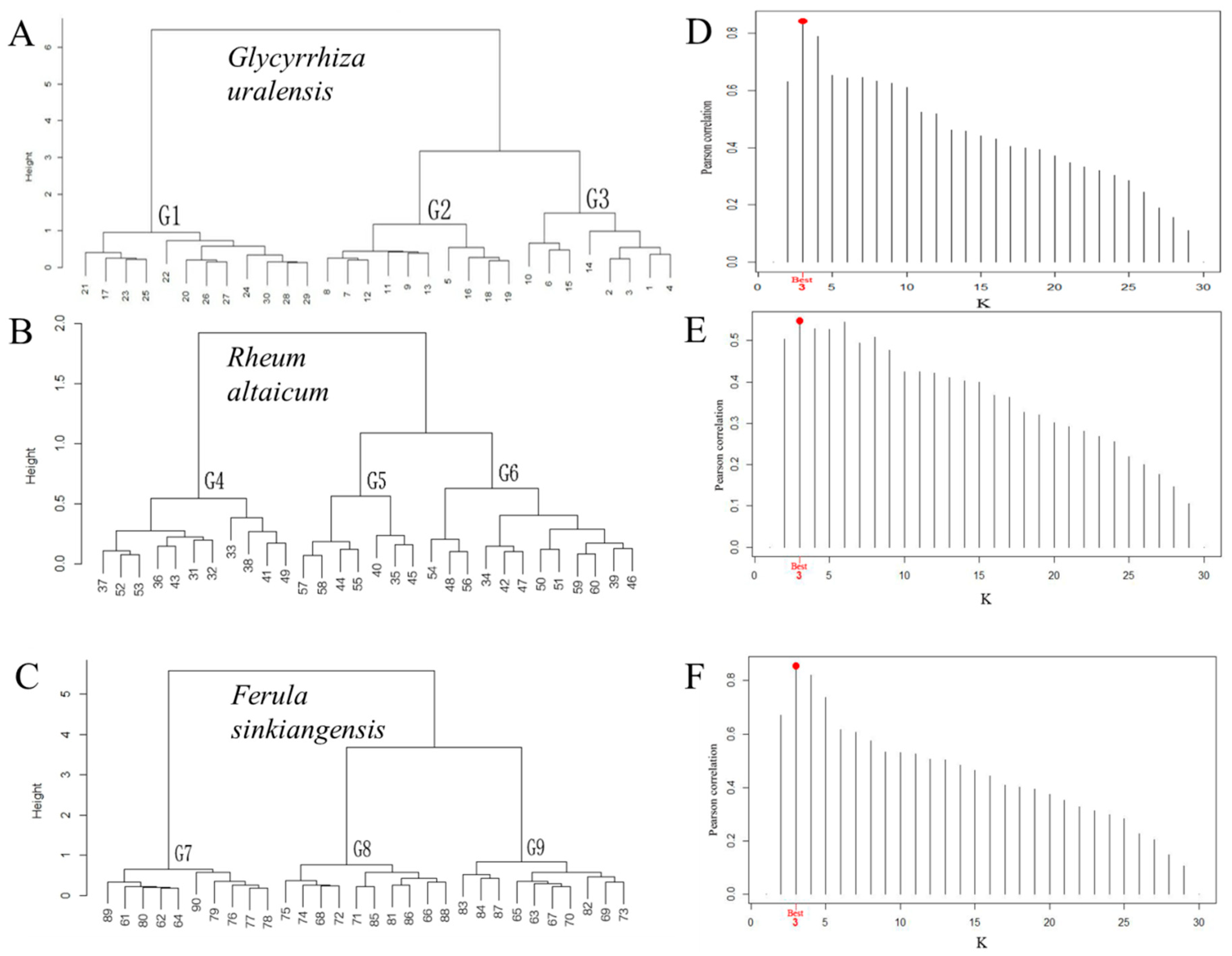
2.5. Description of Statistical Analysis and the Working Model
3. Results
3.1. Vegetation Characteristics of Medicinal Plant Communities
3.2. Vegetation Biodiversity of Medicinal Plant Communities
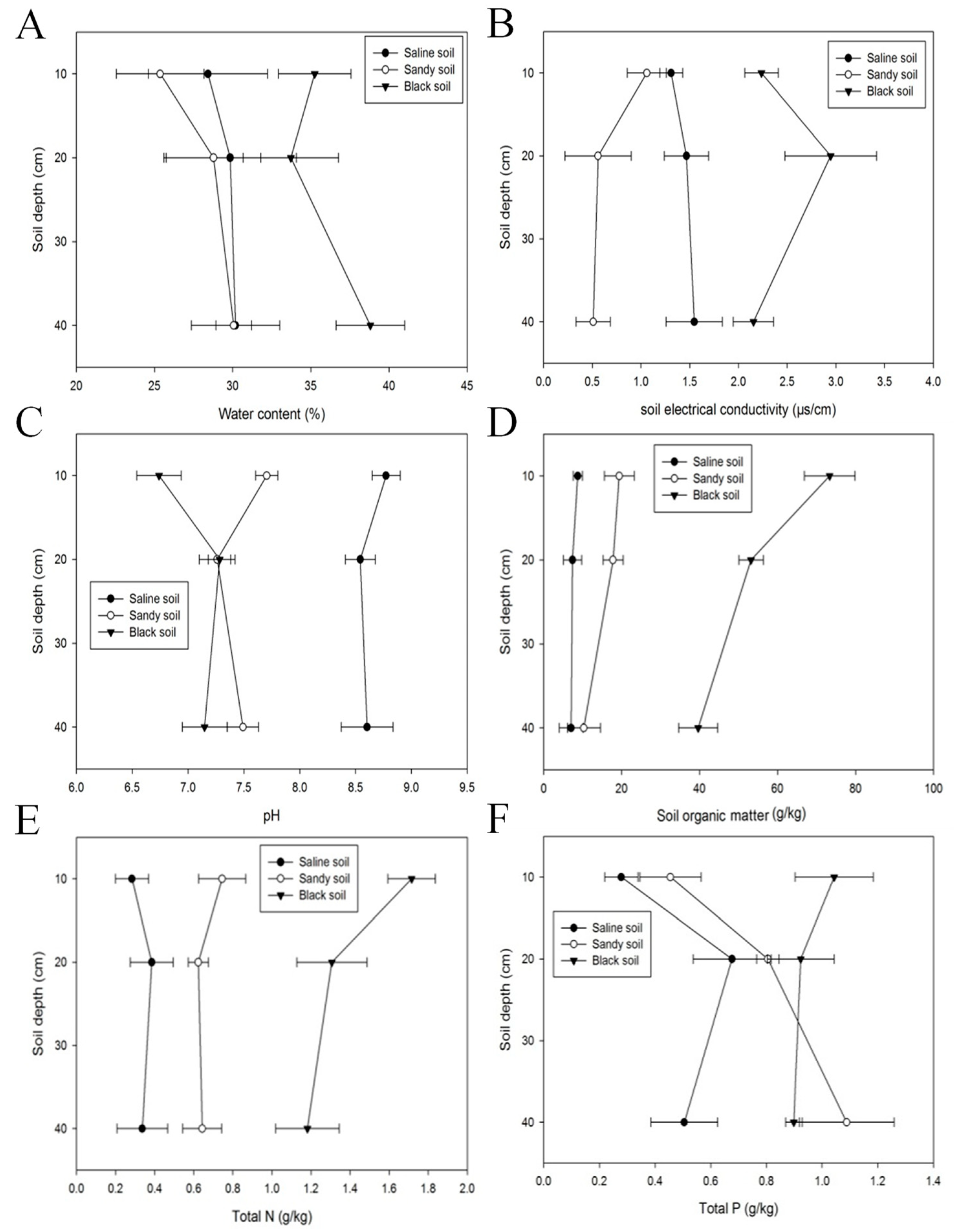
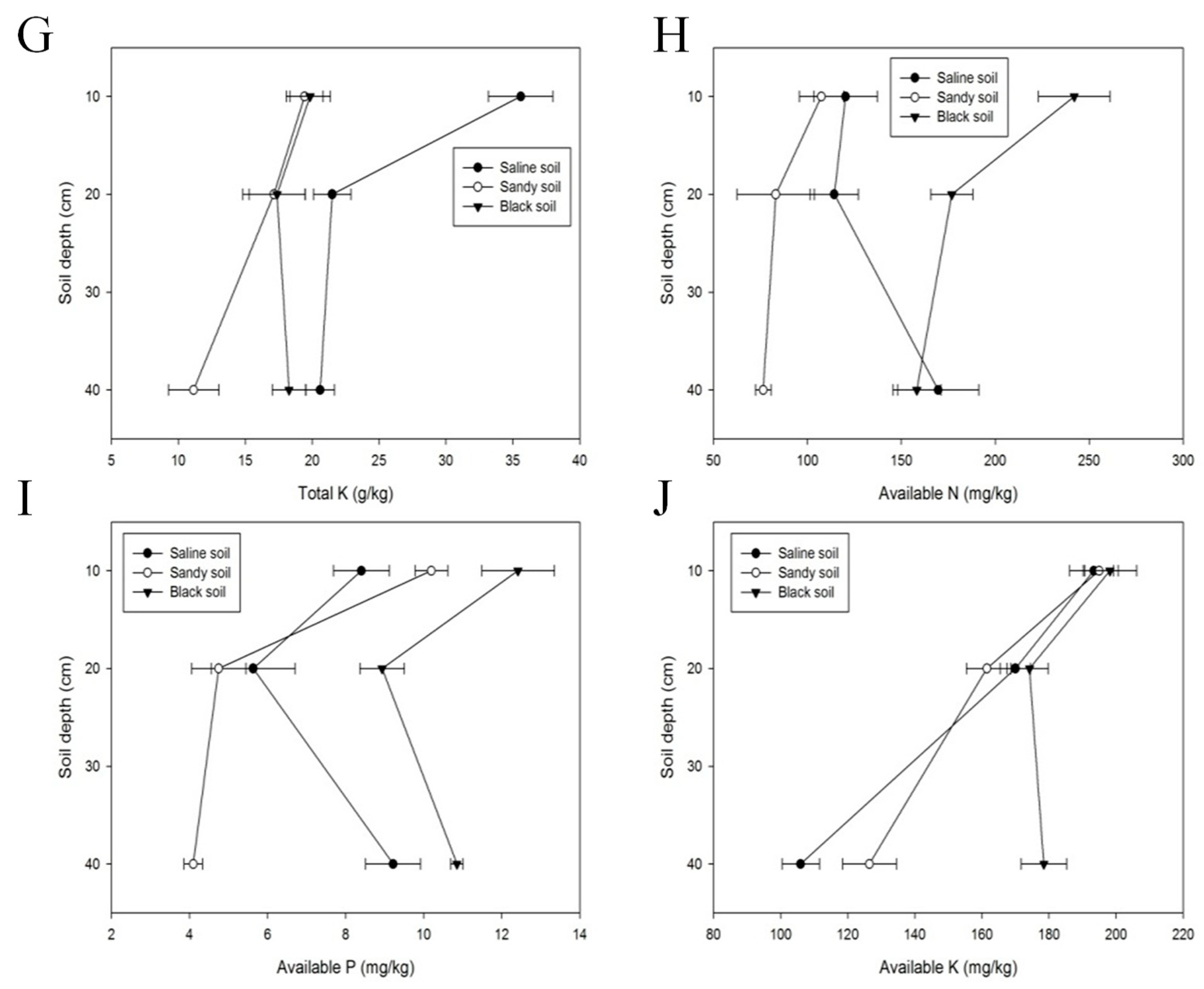
3.3. Soil Properties in Altay Prefecture
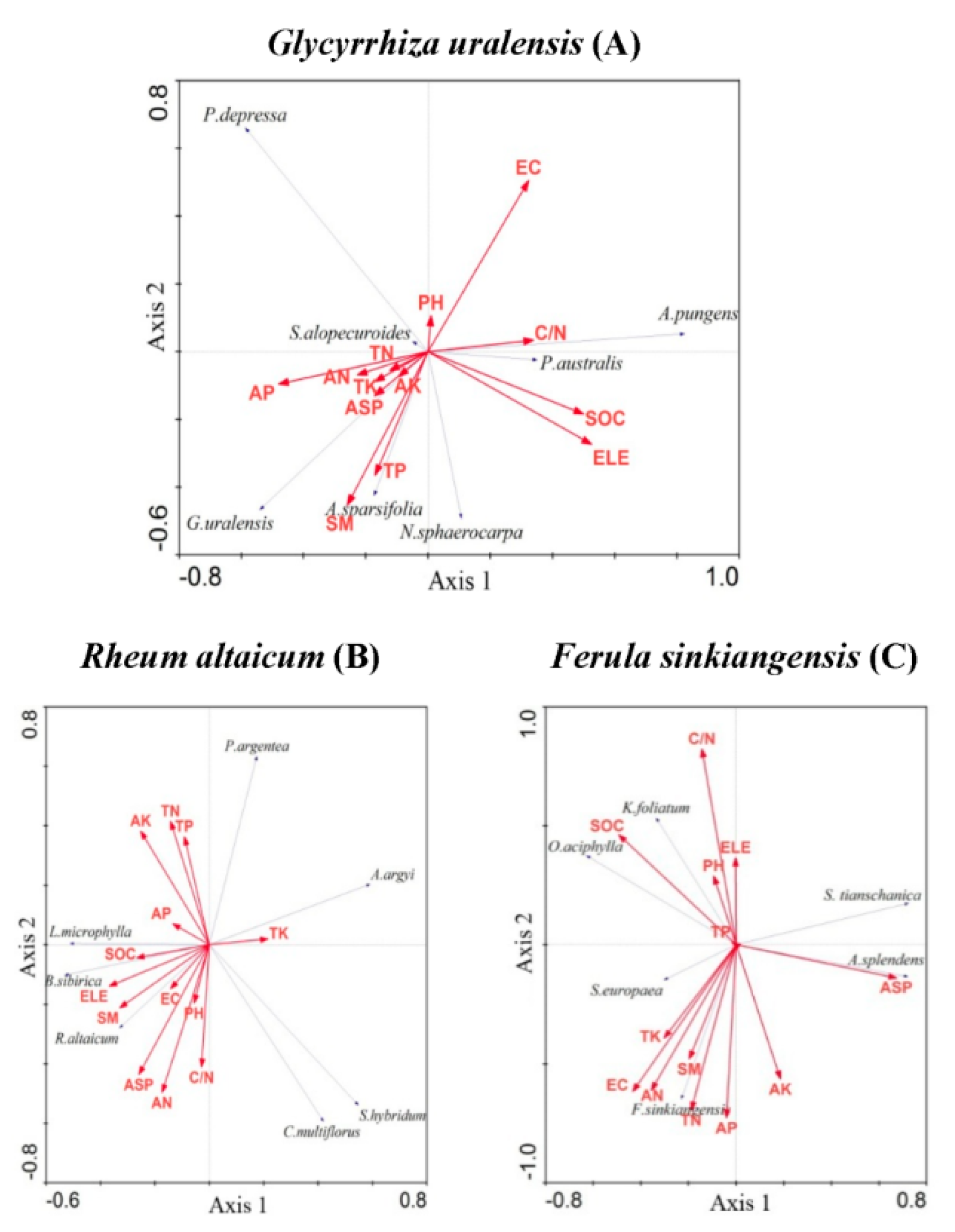
3.4. Response of Medicinal Plants to Their Soil Properties
4. Discussion
4.1. Medicinal Value of Medicinal Plants and Their Community Characteristics
4.2. Soil Properties in Altay Prefecture
4.3. Relationship between Medicinal Vegetation and Soil Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| R | Patrick richness index |
| D | Simpson index |
| H′ | Shannon–Wiener index |
| PIE | Interspecies encounter rate |
| J | Pielou community evenness index |
| C | Simpson dominance index |
| ELE | Elevation |
| Ph | Phanerophytes |
| Ch | Chamaephytes |
| ASP | Aspect |
| SM | Soil moisture |
| PH | pH value |
| EC | Electric conductivity |
| SOC | Soil organic content |
| TN | Total nitrogen |
| TP | Total phosphorus |
| TK | Total potassium |
| C/N | Carbon/nitrogen ratio |
| AN | Available nitrogen |
| AP | Available phosphorus |
| AK | Available potassium |
| RDA | Redundancy analysis |
References
- Legendre, P.; Fortin, M.J. Spatial Pattern and Ecological Analysis. Vegetatio 1989, 80, 107–138. [Google Scholar] [CrossRef]
- Pinder, J.E.; Kroh, G.C.; White, J.D.; May, A.M.B. The relationships between vegetation type and topography in Lassen volcanic national park. Plant Ecol. 1997, 131, 17–29. [Google Scholar] [CrossRef]
- Woodward, F.I.; Mckee, I.F. Vegetation and climate. Environ. Int. 1991, 17, 535–546. [Google Scholar] [CrossRef]
- Burke, A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia. J. Veg. Sci. 2001, 12, 53–60. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the Spatial Component of Ecological Variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- May, R.M. An Overview: Real and Apparent Patterns in Community Structure: Ecological Communities Conceptual Issues and the Evidence; Princeton University Press: Princeton, NJ, USA, 1992. [Google Scholar]
- Yinglan, A.; Wang, G.; Liu, T.; Shrestha, S.; Xue, B.; Tan, Z. Vertical variations of soil water and its controlling factors based on the structural equation model in a semi-arid grassland. Sci. Total Environ. 2019, 691, 1016–1026. [Google Scholar]
- Pausas, J.G.; Austin, M.P. Patterns of plant species richness in relation to different environments: An appraisal. J. Veg. Sci. 2001, 12, 153–166. [Google Scholar] [CrossRef]
- Merunkova, K.; Chytry, M. Environmental control of species richness and composition in upland grasslands of the southern Czech Republic. Plant Ecol. 2012, 213, 591–602. [Google Scholar] [CrossRef]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Hejcman, M.; Ceskova, M.; Schellberg, J.; Paetzold, S. The Rengen Grassland experiment: Effect of soil chemical properties on biomass production, plant species composition and species richness. Folia Geobot. 2010, 45, 125–142. [Google Scholar] [CrossRef]
- Cui, B.S.; Zhai, H.J.; Dong, S.K.; Chen, B.; Liu, S.L. Multivariate analysis of the effects of edaphic and topographical factors on plant distribution in the Yilong Lake Basin of Yun-Gui Plateau, China. Can. J. Plant Sci. 2009, 89, 209–219. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, Y.; Li, W.; Fu, A.; Ma, X.; Gui, D.; Chen, Y. Distribution pattern of plant species diversity in the mountainous Region of Ili River Valley, Xinjiang. Environ. Monit. Assess. 2011, 177, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Springob, G.; Kirchmann, H. Bulk soil C to N ratio as a simple measure of net N mineralization from stabilized soil organic matter in sandy arable soils. Soil Biol. Biochem. 2003, 35, 629–632. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Neuvonen, S. Species richness of vascular plantsin the subarctic landscape of northern Finland: Modelling relationships to the environment. Biodivers. Conserv. 1997, 6, 1181–1201. [Google Scholar] [CrossRef]
- Hajek, M.; Tichy, L.; Schamp, B.S.; Zeleny, D.; Rolecek, J.; Hajkova, P.; Apostolova, I.; Dite, D. Testing the species pool hypothesis for mire vegetation: Exploring the influence of pH specialists and habitat history. Oikos 2007, 116, 1311–1322. [Google Scholar] [CrossRef]
- Zhang, C.; Xie, G.; Fan, S.; Zhen, L. Variation in Vegetation structure and soil properties, and the relation between understory plants and environmental variables under different Phyllostachys pubescens forests in Southeastern China. Environ. Manag. 2010, 45, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.A.; Zhao, X.Y.; Zhao, H.L.; Zhang, T.H.; Li, Y.L.; Wang, S.K.; Li, W.J.; Powers, R. Scale dependent effects of environmental factors on vegetation pattern and composition in Horqin Sandy Land, Northern China. Geoderma 2012, 173–174, 1–9. [Google Scholar] [CrossRef]
- Asanuma, M.; Zhu, S.; Okura, N.; Cai, S.Q.; Yoshimatsu, K.; Komatsu, K. Genetic polymorphism of Japanese cultivated Rheum species in the internal transcribed spacer region of nuclear ribosomal DNA. J. Nat. Med. 2019, 73, 541–554. [Google Scholar] [CrossRef]
- Motai, T.; Kitanaka, S. Sesquiterpenoids from Ferula fukanensis and their inhibitory effects on nitric oxide production. J. Nat. Med. 2006, 60, 54–57. [Google Scholar] [CrossRef]
- Mochida, K.; Sakurai, T.; Seki, H.; Yoshida, T.; Takahagi, K.; Sawai, S.; Uchiyama, H.; Muranaka, T.; Saito, K. Draft genome assembly and annotation of Glycyrrhiza uralensis, a medicinal legume. Plant J. 2017, 89, 181–194. [Google Scholar] [CrossRef]
- Zhang, Q.; Ye, M. Chemical analysis of the Chinese herbal medicine Gan-Cao (licorice). J. Chromatogr. A 2009, 1216, 1954–1969. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.F.; Zhao, Z.M.; Li, J.C.; Yang, G.Z.; Liu, Y.Q.; Dai, L.X.; Zhang, Z.J.; Yang, Z.G.; Miao, X.L.; Yang, C.J.; et al. Insecticidal and antifungal activities of Rheum palmatum L. anthraquinones and structurally related compounds. Ind. Crop. Prod. 2019, 137, 508–520. [Google Scholar] [CrossRef]
- Kuramato, R.T.; Bliss, L.C. Ecology of subalpine meadows in the Olympic mountain, Washingtion. Ecol. Monogr. 1970, 40, 317–347. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, W.; Ye, R.; Lu, X.; Zhou, Y. Responses of reproduction and important value of dominant plant species in different plant functional type in Kobresia meadow to temperature increase. Russ. J. Ecol. 2013, 44, 484–491. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-total. Methods of soil analysis. Part 2, chemical and microbiological. Soil Sci. Soc. Am. J. 1982, 595–624. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. Part 3. In Chemical Methods; SSSA Book Ser. 5; SSSA: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Bao, S.D. Soil Agricultural Chemical Elements Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Singh, N.; Parida, B.R. Environmental factors associated with seasonal variations of night-time plant canopy and soil respiration fluxes in deciduous conifer forest, Western Himalaya, India. Trees-Struct. Funct. 2019, 33, 599–613. [Google Scholar] [CrossRef]
- Fu, Q.; Li, B.; Hou, Y.; Bi, X.; Zhang, X. Effects of land use and climate change on ecosystem services in Central Asia’s arid regions: A case study in Altay Prefecture, China. Sci. Total Environ. 2017, 607, 633–646. [Google Scholar] [CrossRef]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Song, Y.; Xi, B.; Zhang, M.; He, X. Application of derivative synchronous fluorescence spectroscopy (DSFS) to indicate salinisation processes of saline soils in semi-arid region. Ecol. Indic. 2012, 18, 532–539. [Google Scholar] [CrossRef]
- Mou, X.J.; Sun, Z.G. Effects of sediment burial disturbance on seedling emergence and growth of Suaeda salsa in the tidal wetlands of the Yellow River estuary. J. Exp. Mar. Biol. Ecol. 2011, 409, 99–106. [Google Scholar] [CrossRef]
- Ma, M.; Zhou, X.; Ma, Z.; Du, G. Composition of the soil seed bank and vegetation changes after wetland drying and soil salinization on the Tibetan Plateau. Ecol. Eng. 2012, 44, 18–24. [Google Scholar] [CrossRef]
- Xi, H.Y.; Feng, Q.; Zhang, L.; Si, J.H.; Chang, Z.Q.; Yu, T.F.; Guo, R. Effects of water and salinity on plant species composition and community succession in Ejina Desert Oasis, northwest China. Environ. Earth Sci. 2016, 75, 138. [Google Scholar] [CrossRef]
- Knapp, P.A. Secondary plant succession and vegetation recovery in two western Great Basin Desert ghost towns. Biol. Conserv. 1992, 60, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Parraga-Aguado, I.; Gonzalez-Alcaraz, M.N.; Alvarez-Rogel, J.; Jimenez-Carceles, F.J.; Conesa, H.M. The importance of edaphic niches and pioneer plant species succession for the phytomanagement of mine tailings. Environ. Pollut. 2013, 176, 134–143. [Google Scholar] [CrossRef]
- Odland, A. Patterns in the secondary succession of a Carex vesicaria L. wetland following a permanent drawdown. Aquat. Bot. 2002, 74, 233–244. [Google Scholar] [CrossRef]
- Olff, H.; Leeuw, J.D.; Bakker, J.P.; Platerink, R.J.; Wijnen, H.J.V. Vegetation succession and herbivory in a salt marsh: Changes induced by sea level rise and silt deposition along an elevational gradient. J. Ecol. 1997, 85, 799–814. [Google Scholar] [CrossRef] [Green Version]
- Frederic, E.C. Scientific Books: Plant Succession. An analysis of the development of vegetation. Science 1916, 45, 339–341. [Google Scholar]
- Wang, T.; Zhang, X.; Li, C. Growth, abscisic acid content, and carbon isotope composition in wheat cultivars grown under different soil moisture. Biol. Plant. 2007, 51, 181–184. [Google Scholar] [CrossRef]
- Rocha, L.C.; Teixeira, F.A.; Pedreira, M.d.S.; Fries, D.D.; Dias Daniel, L.S.; Costa Evely, G.L.; Figueiredo, A.J.; Seixas, A.A.; Pacheco, C.C.; Santiago, B.M. Plant growth regulator and soil fertilizer improve production and growing stage of Brachiaria decumbens. Grassl. Sci. 2020, 66, 102–109. [Google Scholar] [CrossRef]
- Major, J.; Lehmann, J.; Rondon, M.; Goodale, C. Fate of soil-applied black carbon: Downward migration, leaching and soil respiration. Glob. Chang. Biol. 2010, 16, 1366–1379. [Google Scholar] [CrossRef]
- Lang, T.; Deng, S.; Zhao, N.; Deng, C.; Zhang, Y.; Zhang, Y.; Zhang, H.; Sa, G.; Yao, J.; Wu, C.; et al. Salt-sensitive signaling networks in the mediation of K+/Na+ homeostasis gene expression in Glycyrrhiza uralensis roots. Front. Plant Sci. 2017, 8, 1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingo, F.; Brenner, A.J.; Gutiérrez, L.; Clark, S.C.; Incoll, L.D.; Aguilera, C. Water relations only partly explain the distributions of three perennial plant species in a semi-arid environment. Biol. Plant. 2003, 46, 257–262. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, L.; Liu, T.; Chu, Y.; Shao, X. Effect of different nitrogen rates on Parthenocissus tricuspidata Planch seedling growth and nutrient distribution. Acta Ecol. Sin. 2007, 27, 3435–3441. [Google Scholar]
- Cadot, S.; Bélanger, G.; Ziadi, N.; Morel, C.; Sinaj, S. Critical plant and soil phosphorus for wheat, maize, and rapeseed after 44 years of P fertilization. Nutr. Cycl. Agroecosyst. 2018, 112, 417–433. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Community Type | Altitude, m | Main Accompanying Species | Height, m | Plant Number | Crown Canopy, m2 | Canopy, % | Life Form |
|---|---|---|---|---|---|---|---|
| G. uralensis | 637.4 | P. australis A. pungens S. alopecuroides P. asiatica, and A. sparsifolia | 0.42 | 44.2 | 0.28 × 0.25 | 17.42 | Ph |
| R. altaicum | 1808.8 | S. hybridum A. argyi L. microphylla B. sibirica, and P. argentea | 0.41 | 19.8 | 0.39 × 0.34 | 16.97 | Ch |
| F. sinkiangensis | 997.2 | S. tianschanica, S. europaea A. splendens O. aciphylla, and K. foliatum | 0.80 | 28.5 | 0.42 × 0.29 | 10.9 | Ph |
| Community | Main Species | Quantity | Relative Density, % | Relative Frequency, % | Relative Dominance, % | Importance Value, % | Biomass, g |
|---|---|---|---|---|---|---|---|
| G. uralensis | G. uralensis | 44.2 | 9.23 | 66.67 | 35.83 | 37.24 | 79.66 |
| A. pungens | 271 | 56.60 | 46.67 | 18.87 | 40.71 | 12.37 | |
| P. asiatica | 85.7 | 17.90 | 46.67 | 18.16 | 27.58 | 24.28 | |
| R. altaicum | R. altaicum | 19.8 | 4.38 | 66.67 | 25.08 | 32.04 | 255.00 |
| S. hybridum | 120 | 26.57 | 46.67 | 11.25 | 28.16 | 27.13 | |
| A. argyi | 175 | 38.74 | 33.33 | 16.49 | 29.52 | 12.26 | |
| F. sinkiangensis | F. sinkiangensis | 28.5 | 27.61 | 53.33 | 44.45 | 41.80 | 267.00 |
| S. europaea | 16.6 | 16.09 | 46.67 | 12.04 | 24.93 | 215.90 | |
| S. tianschanica | 29.3 | 28.39 | 66.67 | 29.57 | 41.54 | 29.35 |
| Community | R | D | H′ | PIE | J | C |
|---|---|---|---|---|---|---|
| G. uralensis | 12 | 0.6318 ± 0.0772 a | 1.3705 ± 0.1072 a | 0.7152 ± 0.3397 b | 0.6591 ± 0.0516 a | 0.3682 ± 0.1114 b |
| R. altaicum | 12 | 0.7647 ± 0.1439 b | 1.8528 ± 0.1015 b | 0.0302 ± 0.0682 a | 0.7456 ± 0.0408 a,b | 0.2353 ± 0.0457 a |
| F. sinkiangensis | 7 | 0.7955 ± 0.1012 b | 1.7331 ± 0.0911 b | 0.8033 ± 0.0880 b | 0.8906 ± 0.0468 b | 0.2045 ± 0.0346 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, T.; Pan, L.; Liu, B.; Guo, T.; Hou, X. Vegetation Characteristics and Response to the Soil Properties of Three Medicinal Plant Communities in Altay Prefecture, China. Sustainability 2020, 12, 10306. https://doi.org/10.3390/su122410306
Lang T, Pan L, Liu B, Guo T, Hou X. Vegetation Characteristics and Response to the Soil Properties of Three Medicinal Plant Communities in Altay Prefecture, China. Sustainability. 2020; 12(24):10306. https://doi.org/10.3390/su122410306
Chicago/Turabian StyleLang, Tao, Libo Pan, Bo Liu, Tong Guo, and Xikang Hou. 2020. "Vegetation Characteristics and Response to the Soil Properties of Three Medicinal Plant Communities in Altay Prefecture, China" Sustainability 12, no. 24: 10306. https://doi.org/10.3390/su122410306
APA StyleLang, T., Pan, L., Liu, B., Guo, T., & Hou, X. (2020). Vegetation Characteristics and Response to the Soil Properties of Three Medicinal Plant Communities in Altay Prefecture, China. Sustainability, 12(24), 10306. https://doi.org/10.3390/su122410306