Changes of Soil Microbes Related with Carbon and Nitrogen Cycling after Long-Term CO2 Enrichment in a Typical Chinese Maize Field


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Field Sampling
2.3. Measurement of Soil Carbon and Nitrogen
2.4. DNA Extraction
2.5. PCR and Quantitative Real-Time PCR
2.6. Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
3.1. Effect of ECO2 on the Concentration of Soil Inorganic N, MBC, and MBN
3.2. Responses of Microbes in Terms of Quantity under ECO2 Conditions
3.3. The Diversity of Microbes in Response to the ECO2
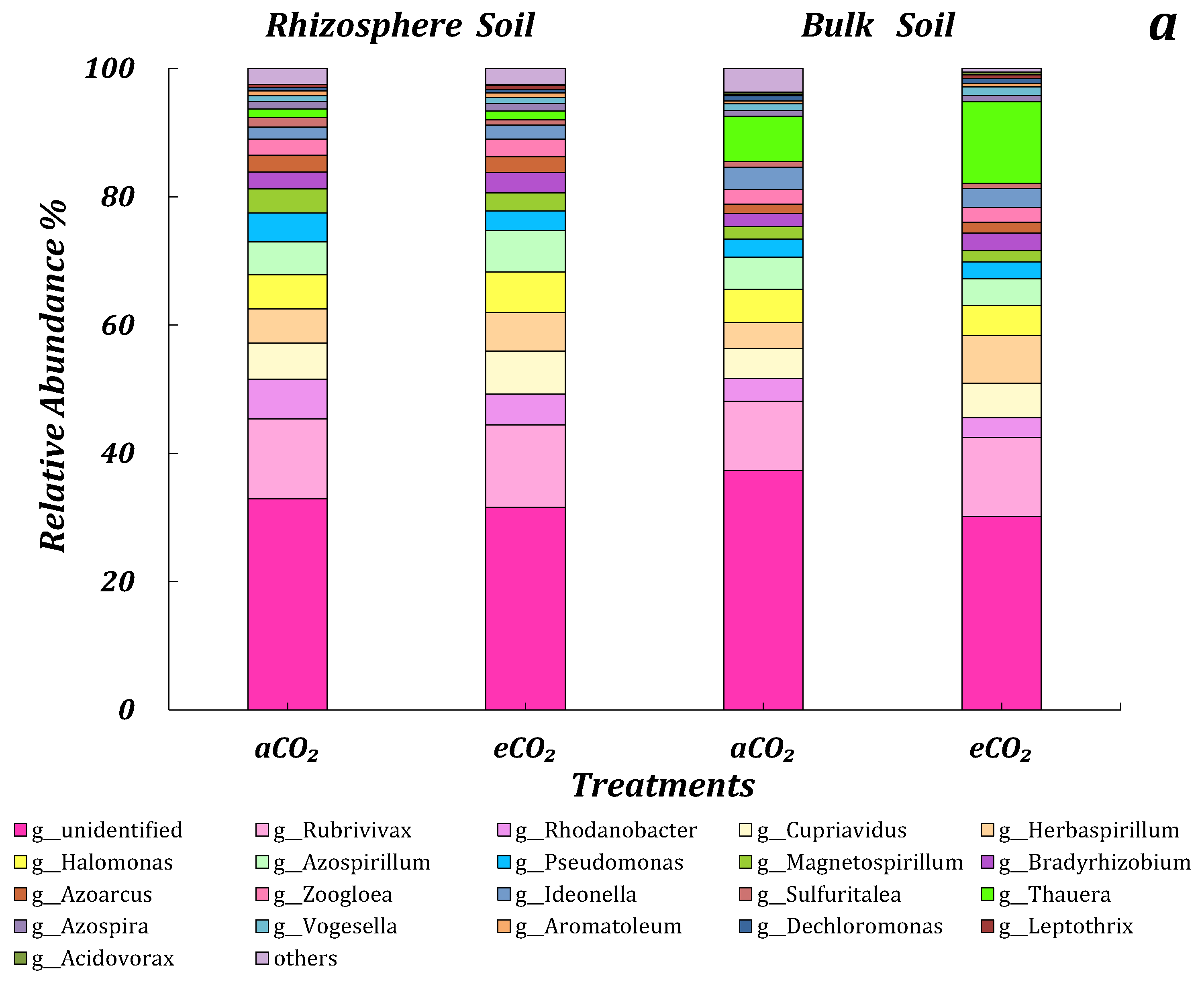
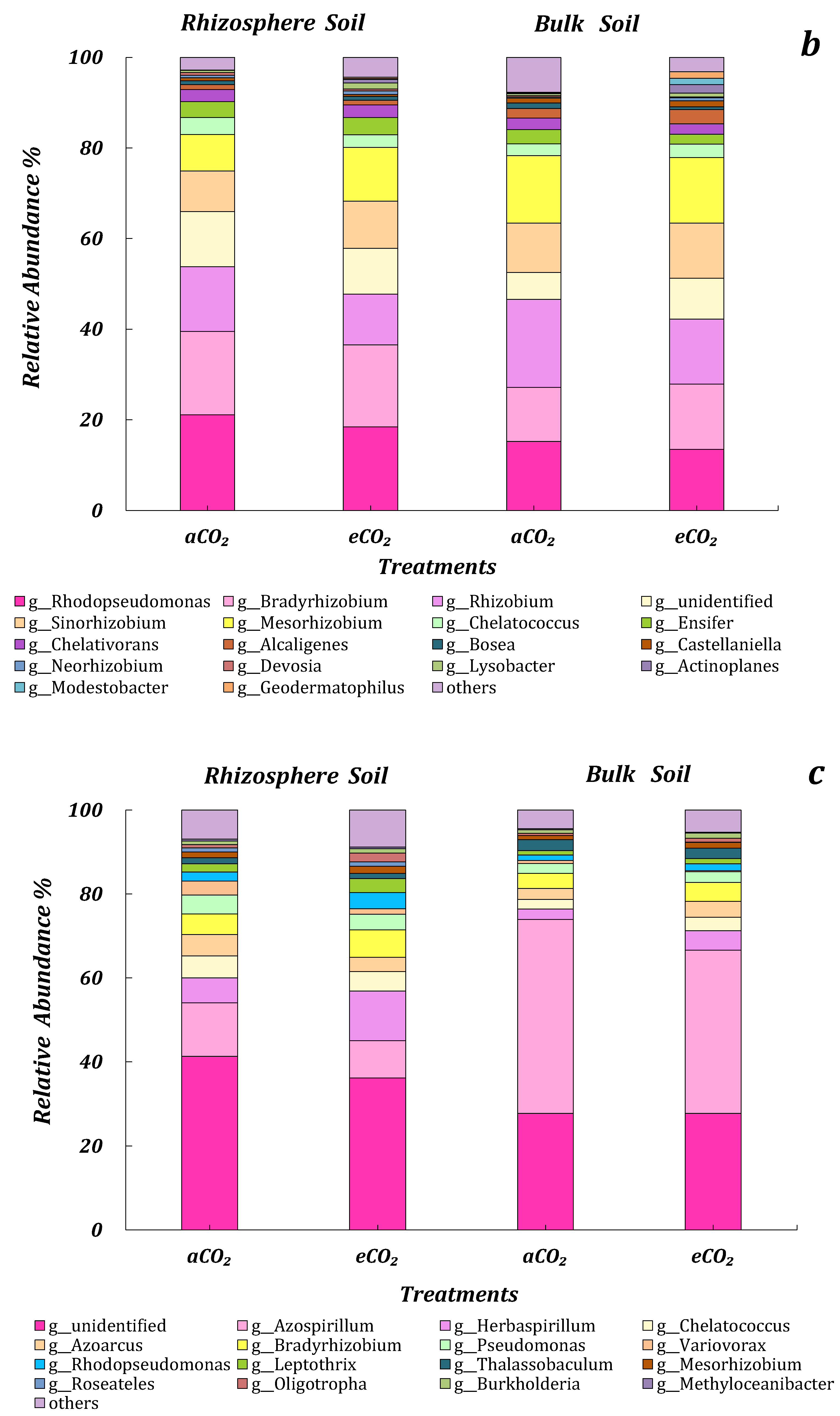
3.4. The Community Structure of Microbes in Response to ECO2
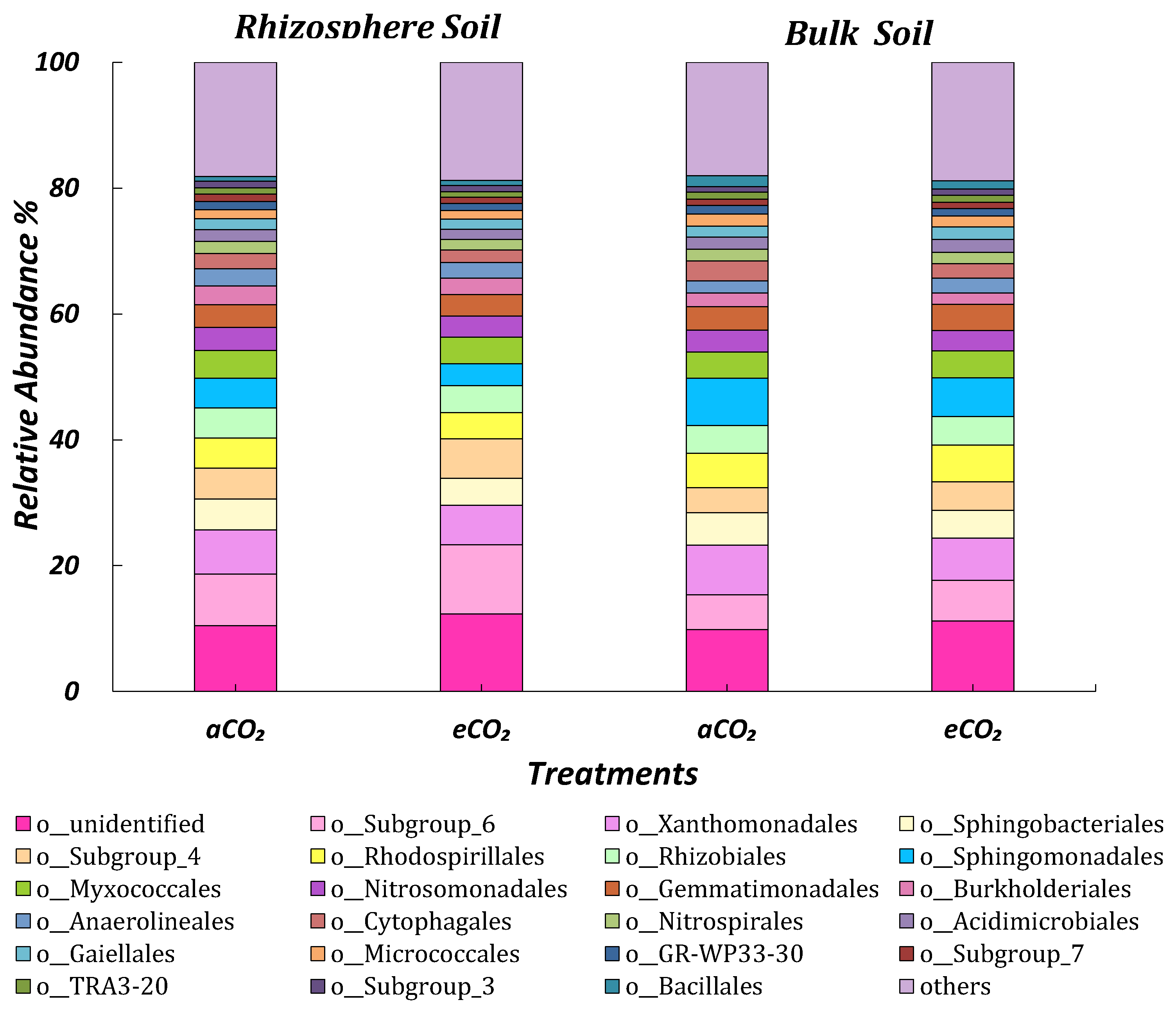
3.4.1. 16s rRNA
3.4.2. ITS
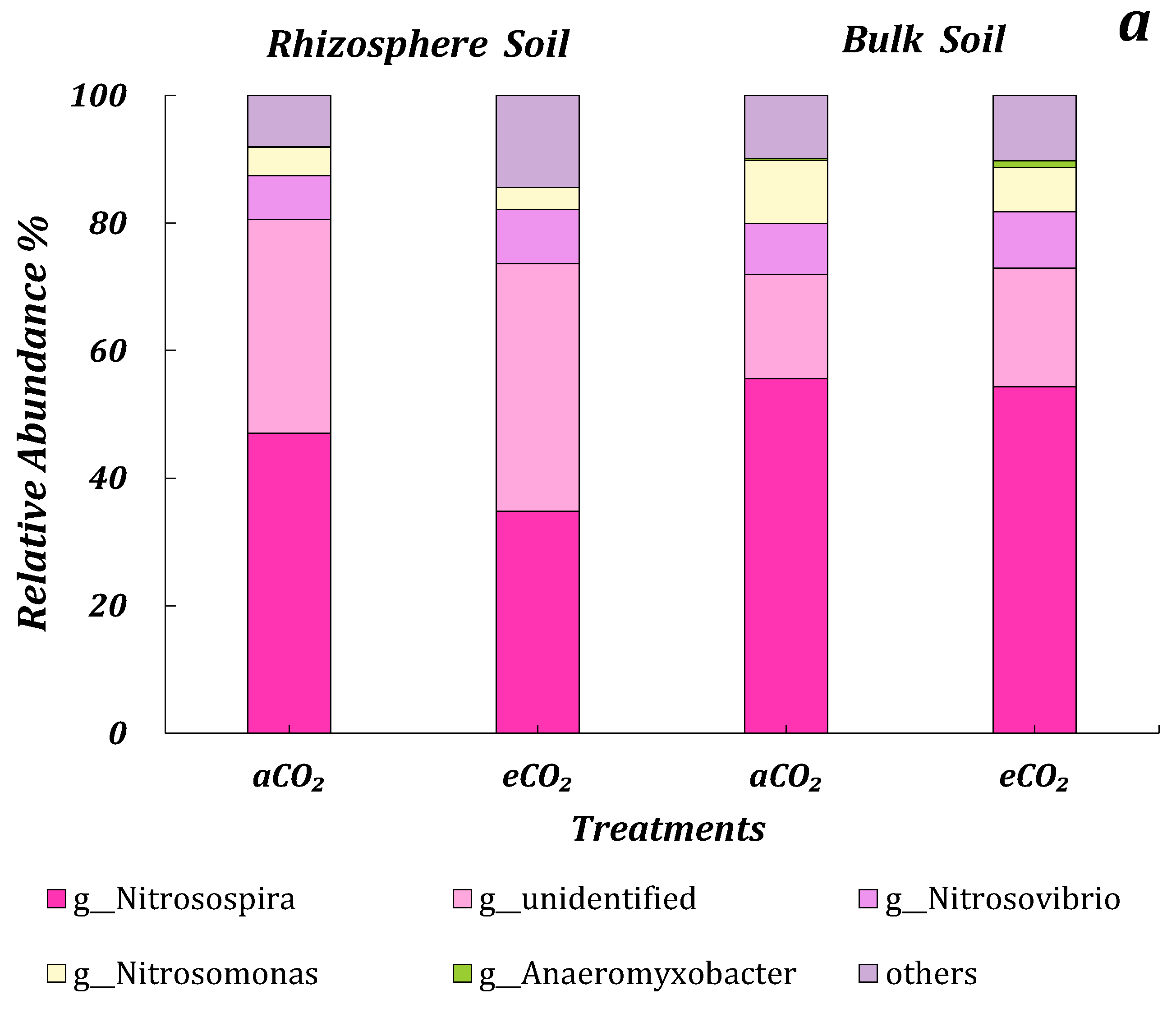
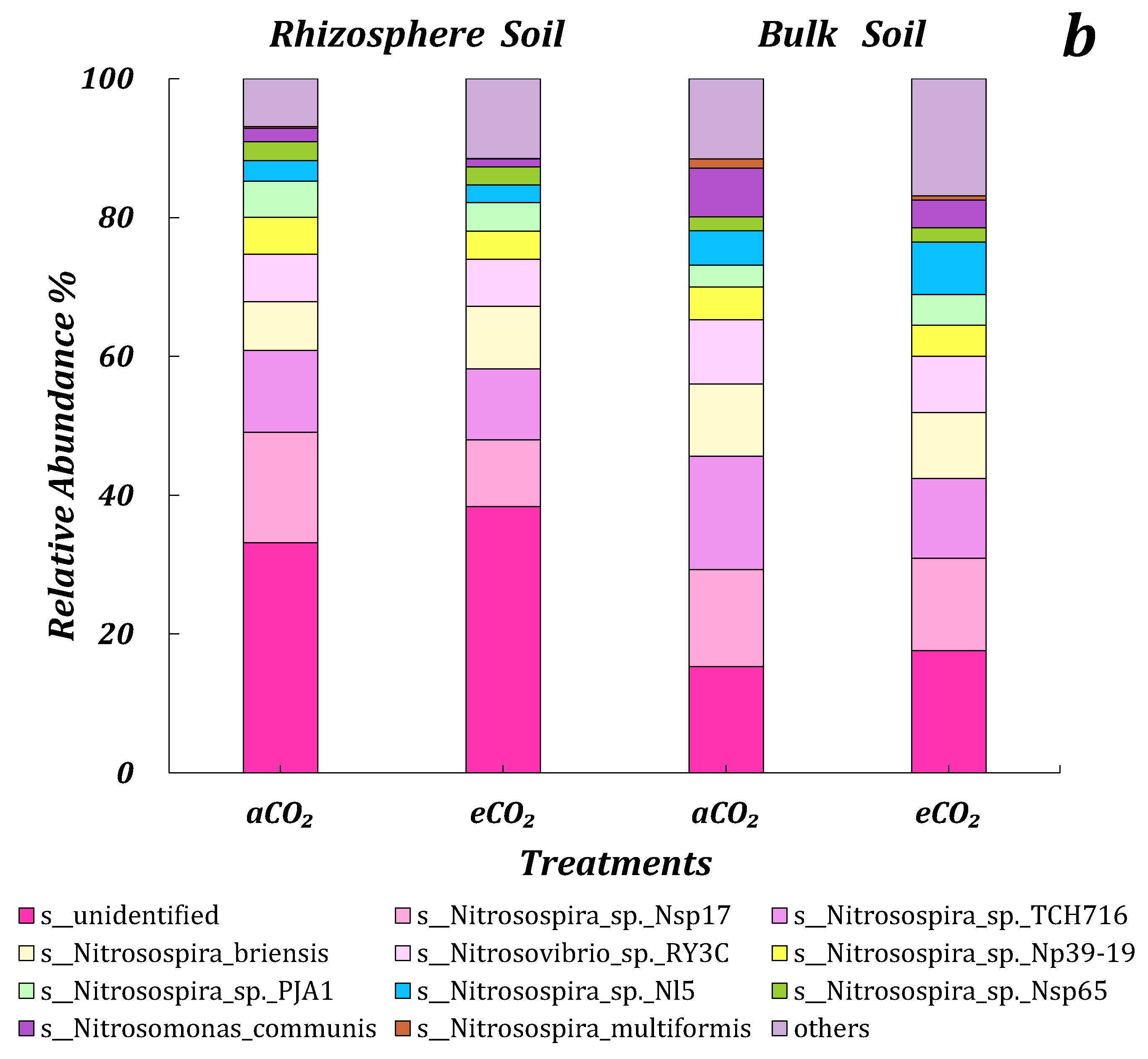
3.4.3. AOB
3.4.4. nirS, nirK and nosZ
4. Discussion
4.1. Effects of Elevated CO2 on Soil Carbon and Associated Microbes
4.2. Effects of Elevated CO2 on Soil N and Relevant Nitrifiers and Detrifiers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruddiman, W.F. The Anthropocene. Annu. Rev. Earth Planet. Sci. 2013, 41, 45–68. [Google Scholar] [CrossRef]
- Pieter Tans, R.K. NOAA/ESRL. 2017. Available online: www.esrl.noaa.gov/gmd/ccgg/trends/ (accessed on 9 July 2017).
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Austin, E.E.; Castro, H.F.; Sides, K.E.; Schadt, C.W.; Classen, A.T. Assessment of 10 years of CO2 fumigation on soil microbial communities and function in a sweetgum plantation. Soil Boil. Biochem. 2009, 41, 514–520. [Google Scholar] [CrossRef]
- Zak, D.R.; Pregitzer, K.S.; Curtis, P.S.; Holmes, W.E. Atmospheric CO2 and the composition and function of soil microbial communities. Ecol. Appl. 2000, 10, 47–59. [Google Scholar]
- Li, Y.; Huang, G.; Shi, Y. Effect of atmospheric CO2 enrichment on soil microbes and related factors. Ying yong The J. of Appl. Ecol. 2003, 14, 2321–2325. (In Chinese) [Google Scholar]
- Phillips, R.P.; Finzi, A.C.; Bernhardt, E.S. Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation. Ecol. Lett. 2011, 14, 187–194. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, J.; Zhu, J.; Liu, G.; Zhu, C.; Wang, S. Ten years of elevated atmospheric CO2 doesn’t alter soil nitrogen availability in a rice paddy. Soil Boil. Biochem. 2016, 98, 99–108. [Google Scholar] [CrossRef]
- Carney, K.M.; Hungate, B.A.; Drake, B.G.; Megonigal, J.P. Altered soil microbial community at elevated CO2 leads to loss of soil carbon. Proc. Natl. Acad. Sci. USA 2007, 104, 4990–4995. [Google Scholar] [CrossRef] [Green Version]
- Langley, J.A.; McKinley, D.C.; Wolf, A.A.; Hungate, B.A.; Drake, B.G.; Megonigal, J.P. Priming depletes soil carbon and releases nitrogen in a scrub-oak ecosystem exposed to elevated CO2. Soil Boil. Biochem. 2009, 41, 54–60. [Google Scholar] [CrossRef]
- Sayer, E.J.; Heard, M.S.; Grant, H.K.; Marthews, T.R.; Tanner, E.V. Soil carbon release enhanced by increased tropical forest litter fall. Nature Clim. Change. 2011, 1, 304–307. [Google Scholar] [CrossRef]
- Drissner, D.; Blum, H.; Tscherko, D.; Kandeler, E. Nine years of enriched CO2changes the function and structural diversity of soil microorganisms in a grassland. Eur. J. Soil Sci. 2007, 58, 260–269. [Google Scholar] [CrossRef]
- He, Z.; Xu, M.; Deng, Y.; Kang, S.; Kellogg, L.; Wu, L.; Van Nostrand, J.D.; Hobbie, S.E.; Reich, P.B.; Zhou, J. Metagenomic analysis reveals a marked divergence in the structure of belowground microbial communities at elevated CO2. Ecol. Lett. 2010, 13, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Zak, D.R.; Pregitzer, K.S.; Curtis, P.S.; Teeri, J.A.; Fogel, R.; Randlett, D.L. Elevated atmospheric CO2 and feedback between C and N cycles. Plant and Soil. 1993, 151, 105–117. [Google Scholar] [CrossRef]
- Schortemeyer, M.; Hartwig, U.A.; Hendrey, G.R.; Sadowsky, M.J. Microbial community changes in the rhizospheres of white clover and perennial ryegrass exposed to Free Air Carbon dioxide Enrichment (FACE). Soil Boil. Biochem. 1996, 28, 1717–1724. [Google Scholar] [CrossRef]
- Cheng, L.; Booker, F.L.; Tu, C.; Burkey, K.O.; Zhou, L.; Shew, H.D.; Rufty, T.W.; Hu, S. Arbuscular Mycorrhizal Fungi Increase Organic Carbon Decomposition Under Elevated CO2. Science 2012, 337, 1084–1087. [Google Scholar] [CrossRef]
- Verhamme, D.T.; Prosser, J.I.; Nicol, G.W. Ammonia concentration determines differential growth of ammonia-oxidizing archaea and bacteria in soil microcosms. The ISME Journal. 2011, 6, 1067e1071. [Google Scholar]
- Xia, W.; Zhang, C.; Zeng, X.; Feng, Y.; Weng, J.; Lin, X.; Zhu, J.; Xiong, Z.; Xu, J.; Cai, Z.; et al. Autotrophic growth of nitrifying community in an agricultural soil. ISME J. 2011, 5, 1226–1236. [Google Scholar] [CrossRef] [Green Version]
- Philippot, L.; Hallin, S.; Schloter, M. Ecology of Denitrifying Prokaryotes in Agricultural Soil. Adv. Agron. 2007, 96, 249–305. [Google Scholar]
- Shapleigh, J.P. Denitrifying Prokaryotes. In The Prokaryotes; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2013; pp. 405–425. [Google Scholar]
- Hungate, B.A.; Canadell, J.; Chapin, F.S. Plant Species Mediate Changes in Soil Microbial N in Response to Elevated CO2. Ecology 1996, 77, 2505–2515. [Google Scholar] [CrossRef]
- Horz, H.-P.; Barbrook, A.; Field, C.B.; Bohannan, B.J.M. Ammonia-oxidizing bacteria respond to multifactorial global change. Proc. Nat. Acad. Sci. USA 2004, 101, 15136–15141. [Google Scholar]
- Barnard, R.; Barthes, L.; Le Roux, X.; Harmens, H.; Raschi, A.; Soussana, J.-F.; Winkler, B.; Leadley, P.W. Atmospheric CO2 elevation has little effect on nitrifying and denitrifying enzyme activity in four European grasslands. Glob. Chang. Boil. 2004, 10, 488–497. [Google Scholar] [CrossRef]
- Barnard, R.; Leadley, P.W.; Hungate, B.A. Global change, nitrification, and denitrification: A review. Glob. Biogeochem. Cycles 2005, 19, 1007. [Google Scholar] [CrossRef]
- Hunger, S.; Schmidt, O.; Hilgarth, M.; Horn, M.A.; Kolb, S.; Conrad, R.; Drake, H.L. Competing Formate- and Carbon Dioxide-Utilizing Prokaryotes in an Anoxic Methane-Emitting Fen Soil▿†. Appl. Environ. Microbiol. 2011, 77, 3773–3785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, D.G.; Firestone, M.; Herman, D.J.; Turetsky, M.; Waldrop, M.; Blazewicz, S.J. Abundance of microbial genes associated with nitrogen cycling as indices of biogeochemical process rates across a vegetation gradient in Alaska. Environ. Microbiol. 2012, 14, 993–1008. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Megonigal, P.J.; Kang, H. How do Elevated CO2 and Nitrogen Addition Affect Functional Microbial Community Involved in Greenhouse Gas Flux in Salt Marsh System. Microb. Ecol. 2017, 81, 169–680. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Gao, B.; Hu, X.-K.; Lu, X.; Well, R.; Christie, P.; Bakken, L.R.; Ju, X.-T. Ammonia-oxidation as an engine to generate nitrous oxide in an intensively managed calcareous Fluvo-aquic soil. Sci. Rep. 2014, 4, 3950. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Diana, R.N.; Steven, K.S.; Alan, R.T. Increases in soil respiration following labile carbon additions linked to rapid shifts in soil microbial community composition. Biogeochemistry 2007, 82, 229–240. [Google Scholar] [CrossRef]
- Henry, S.; Texier, S.; Hallet, S.; Bru, D.; Dambreville, C.; Chèneby, D.; Bizouard, F.; Germon, J.C.; Philippot, L. Disentangling the rhizosphere effect on nitrate reducers and denitrifiers: Insight into the role of root exudates. Environ. Microbiol. 2008, 10, 3082–3092. [Google Scholar] [CrossRef]
- Han, X.; Hao, X.; Lam, S.K.; Wang, H.; Li, Y.; Wheeler, T.; Ju, H.; Lin, E. Yield and nitrogen accumulation and partitioning in winter wheat under elevated CO2: A 3-year free-air CO2 enrichment experiment. Agric. Ecosyst. Environ. 2015, 209, 132–137. [Google Scholar] [CrossRef]
- Drigo, B.; Kowalchuk, G.A.; Van Veen, J.A. Climate change goes underground: Effects of elevated atmospheric CO2 on microbial community structure and activities in the rhizosphere. Boil. Fertil. Soils 2008, 44, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Lukac, M.; Lagomarsino, A.; Moscatelli, M.C.; De Angelis, P.; Cotrufo, M.F.; Godbold, D.L. Forest soil carbon cycle under elevated CO2 - a case of increased throughput? Forestry 2009, 82, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Janus, L.R.; Angeloni, N.L.; McCormack, J.; Rier, S.T.; Tuchman, N.C.; Kelly, J.J. Elevated Atmospheric CO2 Alters Soil Microbial Communities Associated with Trembling Aspen (Populus tremuloides) Roots. Microb. Ecol. 2005, 50, 102–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, R.L. Lignin biodegradation and transformation; Wiley: New York, NY, USA, 1981; p. 137. [Google Scholar]
- Phillip, B.; Babou, O.J.; Armand, R.K.; Lloyd, A.P.; David, A.L. Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant and Soil. 1997, 197, 61–69. [Google Scholar]
- Guo, L.; Wang, X.; Diao, T.; Ju, X.; Niu, X.; Zheng, L.; Zhang, X.; Han, X. N2O emission contributions by different pathways and associated microbial community dynamics in a typical calcareous vegetable soil. Environ. Pollut. 2018, 242, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting Soil pH Effects on Fungal and Bacterial Growth Suggest Functional Redundancy in Carbon Mineralization▿ †. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, E.; Thornton, B.; Midwood, A.J.; Osborne, S.M.; Sim, A.; Millard, P. Atmospheric CO2 enrichment and nutrient additions to planted soil increase mineralisation of soil organic matter, but do not alter microbial utilisation of plant- and soil C-sources. Soil Boil. Biochem. 2008, 40, 2434–2440. [Google Scholar] [CrossRef]
- Sharona, A.B.; Susane, E.Z. Altered patterns of soil carbon substrate usage and heterotrophic respiration in a pine forest with elevated CO2 and N fertilization. Glob. Change Biol. 2008, 14, 1025–1036. [Google Scholar]
- Kristin, K.; Bernd, W.; Vera, K.; Franziska, W.; Heiko, N.; Ingo, S.; Marion, S.; Rolf, D. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Report. 2016, 6, 33696. [Google Scholar]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Yasukawa, H.; Yamaoka, Y.; Takahashi, N.; Tamura, C.; Morimoto, Y.; Fukushima, S.; Vasavada, R.C. Effects of Oleic Acid/Propylene Glycol on Rat Abdominal Stratum Corneum: Lipid Extraction and Appearance of Propylene Glycol in the Dermis Measured by Fourier Transform Infrared/Attenuated Total Reflectance(FT-IR/ATR) Spectroscopy. Chem. Pharm. Bull. 1993, 41, 1434–1437. [Google Scholar] [CrossRef]
- Gautam, S.P.; Bundela, P.S.; Pandey, A.K.; Jamaluddin; Awasthi, M.K.; Sarsaiya, S. Diversity of Cellulolytic Microbes and the Biodegradation of Municipal Solid Waste by a Potential Strain. Int. J. Microbiol. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clare, E.K. Soil carbon cycling responses to elevated CO2 and nitrogen addition. University of Minnesota, 2017. Available online: https://conservancy.umn.edu/handle/11299/193426 (accessed on 6 November 2008).
- Madi, L.; Katan, T.; Katan, J.; Henis, Y. Biological Control of Sclerotium rolfsii and Verticillium dahliae by Talaromyces flavus Is Mediated by Different Mechanisms. Phytopathology 1997, 87, 1054–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norby, R.J.; Cotrufo, M.F.; Ineson, P.; Canadell, J.G.; Cortufo, M.F.; O’Neill, E.G. Elevated CO2, litter chemistry, and decomposition: A synthesis. Oecologia 2001, 127, 153–165. [Google Scholar] [CrossRef]
- Nowak, R.S.; Ellsworth, D.; Smith, S.D. Functional responses of plants to elevated atmospheric CO2- do photosynthetic and productivity data from FACE experiments support early predictions? New Phytol. 2004, 162, 253–280. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.W.; Wan, G.F.; Zhang, P.; Zhang, S.L.; Guo, Y.; Zhang, W.H. Effects of doubled CO2 and enhanced UV-B radiation on rhizosphere ammonia-oxidizing bacteria and soil enzymes in soybean (Glycine max Merr.). Soybean Sci. 2012, 31, 69–72. (In Chinese) [Google Scholar]
- Liu, Y.; Wang, G.L.; Li, L.Q.; Pan, G.X. Response of Soil Nitrifier and Denitrifier Community and Activity to Elevated Atmospheric CO2 Concentration and Temperature. Environ. Sci. 2017, 38, 1245–1252. (In Chinese) [Google Scholar]
- Nelson, D.M.; Cann, I.K.O.; Mackie, R.I. Response of Archaeal Communities in the Rhizosphere of Maize and Soybean to Elevated Atmospheric CO2 Concentrations. PLoS ONE 2010, 5, e15897. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.G.; Hu, J.L.; Chu, H.Y. Response of soil ammonia-oxidizing bacteria to elevated atmospheric CO2. Rural Eco-Environ. 2005, 21, 44–46. [Google Scholar]
- Kowalchuk, G.A.; Stephen, J.R. Ammonia-Oxidizing Bacteria: A Model for Molecular Microbial Ecology. Annu. Rev. Microbiol. 2001, 55, 485–529. [Google Scholar] [CrossRef] [Green Version]
- Kowalchuk, G.A.; Stienstra, A.W.; Heilig, G.H.; Stephen, J.R.; Woldendorp, J.W. Changes in the community structure of ammonia-oxidizing bacteria during secondary succession of calcareous grasslands. Environ. Microbiol. 2, 99–110. [CrossRef]
- Kowalchuk, G.A.; Stienstra, A.W.; Heilig, G.H.; Stephen, J.R.; Woldendorp, J.W. Molecular analysis of ammonia-oxidizing bacteria in soil of successional grasslands of the Drentsche A (The Netherlands). FEMS Microbiol. Ecol. 31, 207–215. [CrossRef] [PubMed]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; He, P.; Zhou, W. Different roles of rhizosphere effect and long-term fertilization in the activity and community structure of ammonia oxidizers in a calcareous fluvo-aquic soil. Soil Boil. Biochem. 2013, 57, 30–42. [Google Scholar] [CrossRef]
- Shen, J.-P.; Zhang, L.-M.; Zhu, Y.-G.; Zhang, J.-B.; He, J.-Z. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam. Environ. Microbiol. 2008, 10, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lu, L.; Wang, B.; Lin, X.; Zhu, J.; Cai, Z.; Yan, X.; Jia, Z. Long-Term Field Fertilization Significantly Alters Community Structure of Ammonia-Oxidizing Bacteria rather than Archaea in a Paddy Soil. Soil Sci. Soc. Am. J. 2011, 75, 1431–1439. [Google Scholar] [CrossRef]
- Fan, F.; Yang, Q.; Li, Z.; Wei, D.; Cui, X.; Liang, Y. Impacts of Organic and Inorganic Fertilizers on Nitrification in a Cold Climate Soil are Linked to the Bacterial Ammonia Oxidizer Community. Microb. Ecol. 2011, 62, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Yeom, J.; Kim, J.; Han, J.; Lim, H.S.; Park, H.; Hyun, S.; Park, W. Change in gene abundance in the nitrogen biogeochemical cycle with temperature and nitrogen addition in Antarctic soils. Res. Microbiol. 2011, 162, 1018–1026. [Google Scholar] [CrossRef]
- Dambreville, C.; Hallet, S.; Nguyen, C.; Morvan, T.; Germon, J.-C.; Philippot, L.; Hallet, S. Structure and activity of the denitrifying community in a maize-cropped field fertilized with composted pig manure or ammonium nitrate. FEMS Microbiol. Ecol. 2006, 56, 119–131. [Google Scholar] [CrossRef]
- Baldani, J.I.; Baldani, V.L.D.; Seldin, L.; Döbereiner, J. Characterization of Herbaspirillum seropedicae gen. nov., sp. nov., a Root-Associated Nitrogen-Fixing Bacterium. Int. J. Syst. Bacteriol. 1986, 36, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Ishii, S.; Ohno, H.; Tsuboi, M.; Otsuka, S.; Senoo, K. Identification and isolation of active N2O reducers in rice paddy soil. ISME J. 2011, 5, 1936–1945. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, H.M.; Wang, J.Q.; Liu, X.Y.; Cheng, K.; Lia, L.Q.; Zheng, J.W.; Zhang, X.H.; Zheng, J.F.; Pan, G.X. Short-term response of nitrifier communities and potential nitrification activity to elevated CO2 and temperature interaction in a Chinese paddy field. Appl. Soil Ecol. 2015, 96, 88–98. [Google Scholar] [CrossRef]
- Deng, Q.; Cheng, X.; Bowatte, S.; Newton, P.C.; Zhang, Q. Rhizospheric carbon-nitrogen interactions in a mixed-species pasture after 13 years of elevated CO2. Agric. Ecosyst. Environ. 2016, 235, 134–141. [Google Scholar] [CrossRef]
- Booth, M.S.; Stark, J.M.; Rastetter, E. Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data. Ecol. Monogr. 2005, 75, 139–157. [Google Scholar] [CrossRef] [Green Version]
- Sterngren, A.E.; Hallin, S.; Bengtson, P. Archaeal Ammonia Oxidizers Dominate in Numbers, but Bacteria Drive Gross Nitrification in N-amended Grassland Soil. Front. Microbiol. 2015, 6, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, X.; Chen, C.; Xu, Z.; Oren, R.; He, J.-Z. Abundance and community structure of ammonia-oxidizing bacteria and archaea in a temperate forest ecosystem under ten-years elevated CO2. Soil Boil. Biochem. 2012, 46, 163–171. [Google Scholar] [CrossRef]
- Mosler, A.C.; Francls, C.A. Denitrifier abundance and activity across the san Francisco Bay estuary. Environ. Microbiol. 2010, 2, 667e676. [Google Scholar]








{kind=link}
{kind=link}
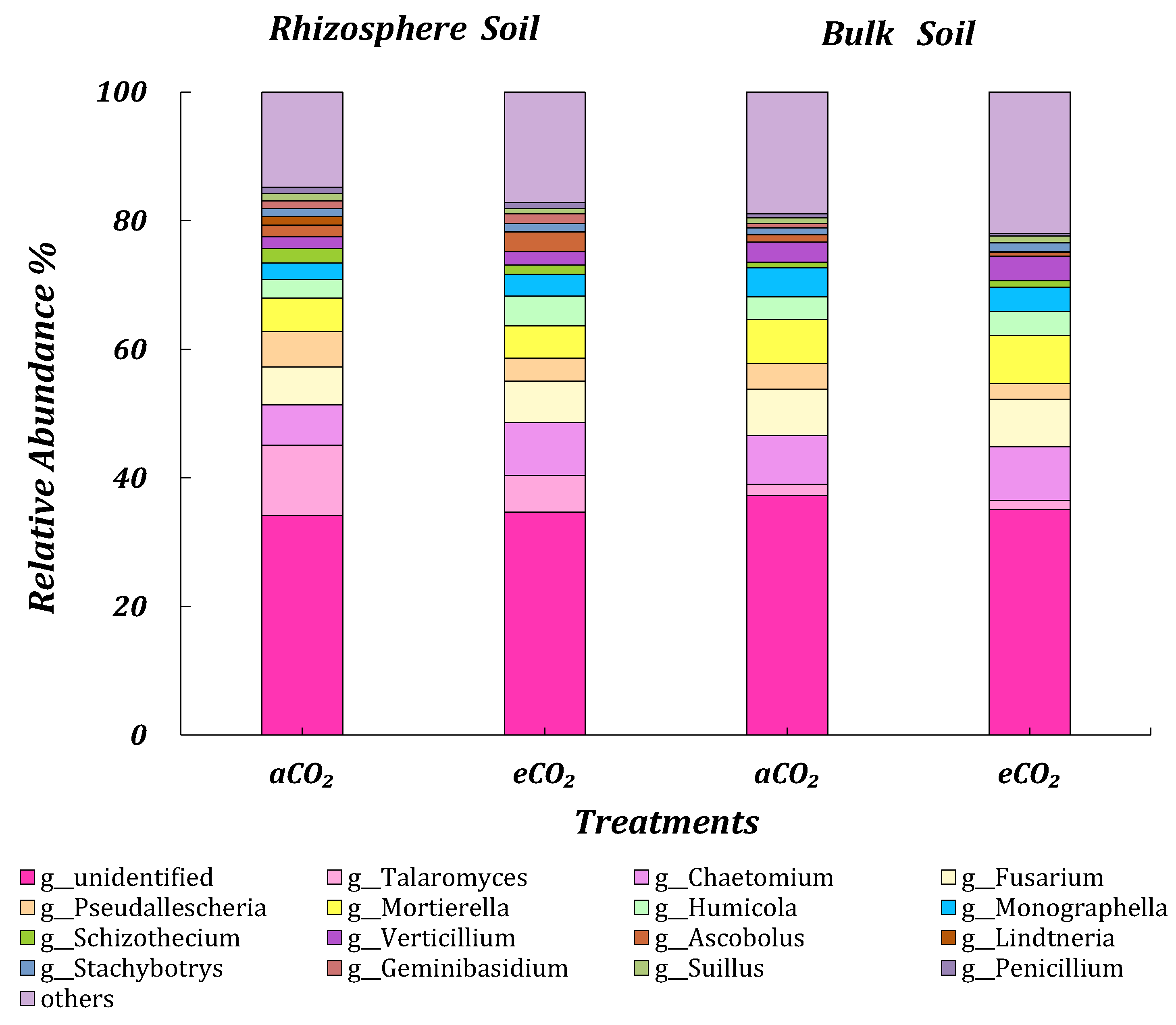{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | 16s rDNA (*109) | ITS (*107) | 16s rDNA/ITS |
|---|---|---|---|
| aCO2-R | 2.36 ± 0.28 a | 1.58 ± 0.05 b | 149.38 b |
| eCO2-R | 1.77 ± 0.10 b | 2.01 ± 0.16 a | 88.67 c |
| aCO2-B | 1.37 ± 0.30 c | 0.16 ± 0.02 d | 547.90 a |
| eCO2-B | 1.95 ± 0.26 b | 1.21 ± 0.09 c | 137.35 b |
| Treatment | AOB (107) | nirS (*106) | nirK (*109) | nosZ (*108) | nosZ/ (nirS + nirK) |
|---|---|---|---|---|---|
| aCO2-R | 6.38 ± 0.10 b | 2.77 ± 0.17 d | 4.64 ± 0.30 b | 4.75 ± 0.48 c | 0.16 b |
| eCO2-R | 6.56 ± 0.39 b | 6.31 ± 0.37 b | 7.63 ± 0.13 a | 25.41 ± 5.53 b | 0.59 a |
| aCO2-B | 6.58 ± 0.99 b | 3.54 ± 0.39 c | 5.29 ± 0.82 ab | 8.56 ± 0.84 c | 0.10 b |
| eCO2-B | 7.54 ± 0.23 a | 9.29 ± 0.54 a | 6.39 ± 0.55 ab | 52.3 ± 2.51 a | 0.33 ab |
| Shannon | 16s rDNA | ITS | AOB | nirS | nirK | nosZ |
|---|---|---|---|---|---|---|
| aCO2-R | 9.85 ± 0.04 a | 5.96 ± 0.25 a | 5.35 ± 0.38 a | 7.09 ± 0.13 a | 6.97 ± 0.27a | 7.91 ± 0.29 a |
| eCO2-R | 9.82 ± 0.05 a | 6.22 ± 0.08 a | 5.31 ± 0.51 a | 7.29 ± 0.02 a | 6.54 ± 0.25a | 8.08 ± 0.39 a |
| aCO2-B | 9.68 ± 0.08 b | 5.93 ± 0.49 a | 5.31 ± 0.61 a | 6.37 ± 0.51 b | 6.60 ± 0.13a | 5.69 ± 1.43 b |
| eCO2-B | 9.76 ± 0.04ab | 6.19 ± 0.13 a | 4.96 ± 0.54 a | 6.85 ± 0.11ab | 6.87 ± 0.22a | 6.02 ± 3.7ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diao, T.; Peng, Z.; Niu, X.; Yang, R.; Ma, F.; Guo, L. Changes of Soil Microbes Related with Carbon and Nitrogen Cycling after Long-Term CO2 Enrichment in a Typical Chinese Maize Field. Sustainability 2020, 12, 1250. https://doi.org/10.3390/su12031250
Diao T, Peng Z, Niu X, Yang R, Ma F, Guo L. Changes of Soil Microbes Related with Carbon and Nitrogen Cycling after Long-Term CO2 Enrichment in a Typical Chinese Maize Field. Sustainability. 2020; 12(3):1250. https://doi.org/10.3390/su12031250
Chicago/Turabian StyleDiao, Tiantian, Zhengping Peng, Xiaoguang Niu, Rongquan Yang, Fen Ma, and Liping Guo. 2020. "Changes of Soil Microbes Related with Carbon and Nitrogen Cycling after Long-Term CO2 Enrichment in a Typical Chinese Maize Field" Sustainability 12, no. 3: 1250. https://doi.org/10.3390/su12031250
APA StyleDiao, T., Peng, Z., Niu, X., Yang, R., Ma, F., & Guo, L. (2020). Changes of Soil Microbes Related with Carbon and Nitrogen Cycling after Long-Term CO2 Enrichment in a Typical Chinese Maize Field. Sustainability, 12(3), 1250. https://doi.org/10.3390/su12031250