Comparison of HPLC Pigment Analysis and Microscopy in Phytoplankton Assessment in the Seomjin River Estuary, Korea

Abstract
:1. Introduction
2. Materials and Methods
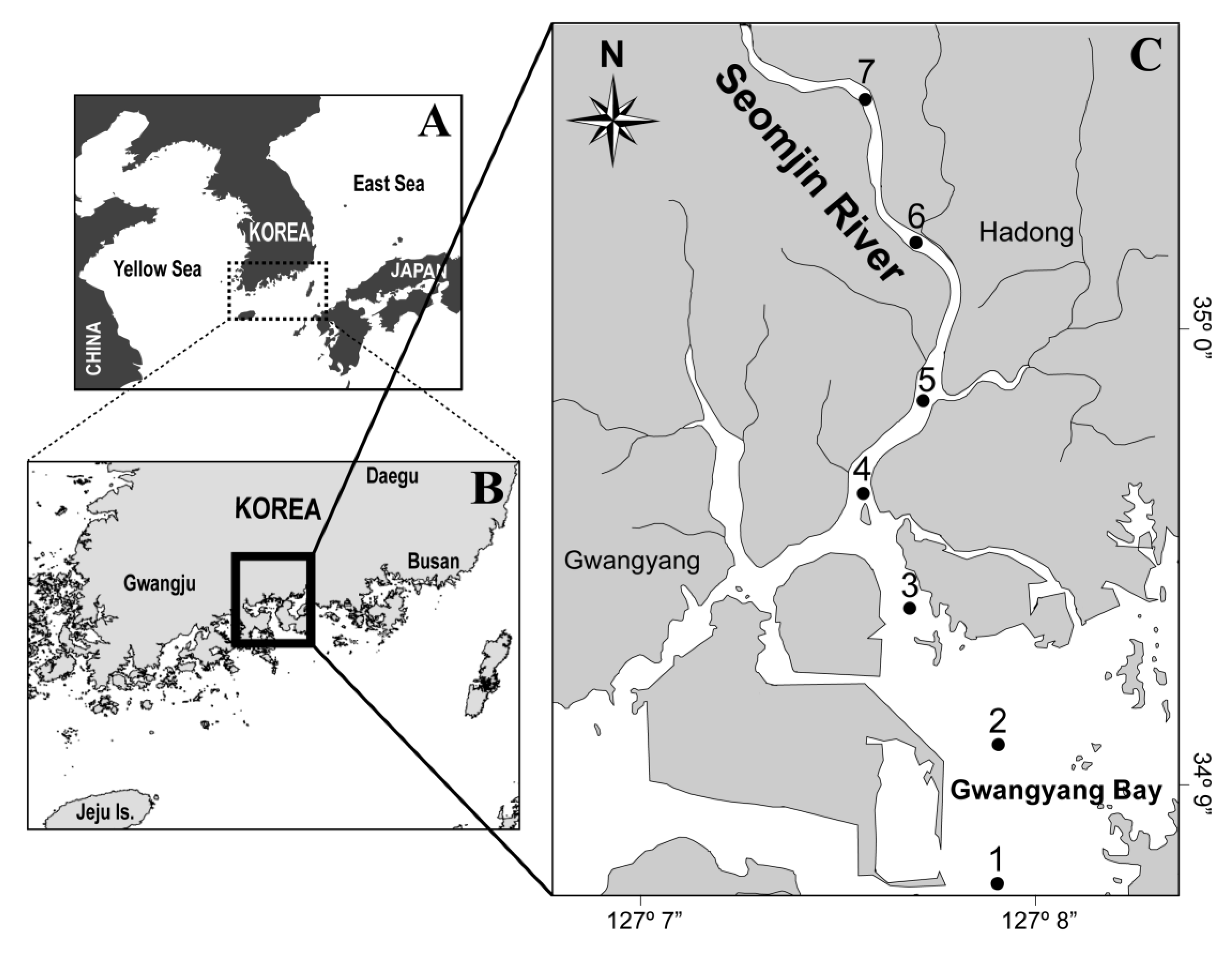
2.1. Field Sampling
2.2. Nutrients Analysis
2.3. Phytoplankton Identification Using Microscopy
2.4. HPLC Analysis
2.5. CHEMTAX Program
2.6. Pigment Indices
2.7. Environmental Data and Statistical Analysis
3. Results
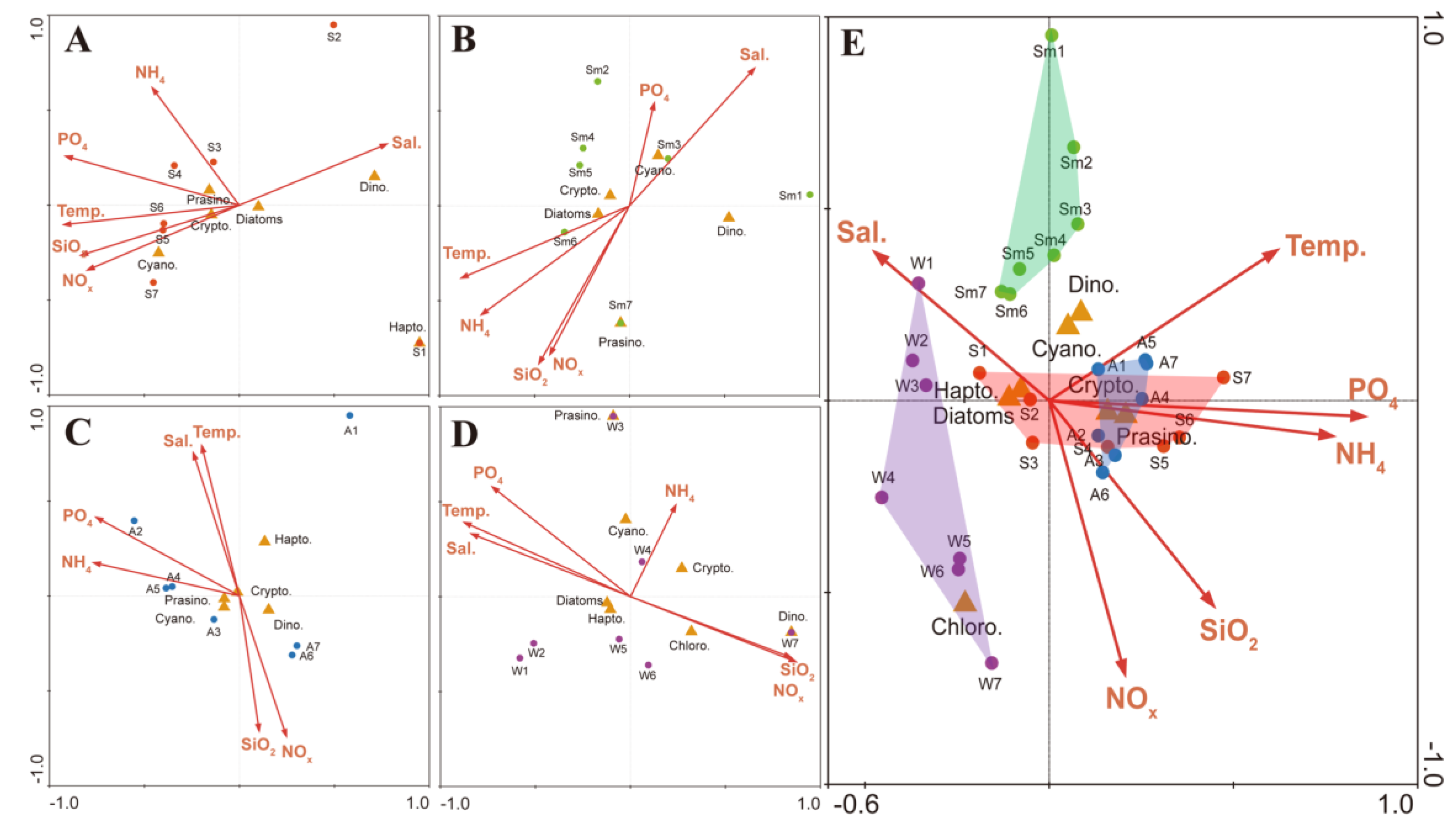
3.1. Environmental Factors
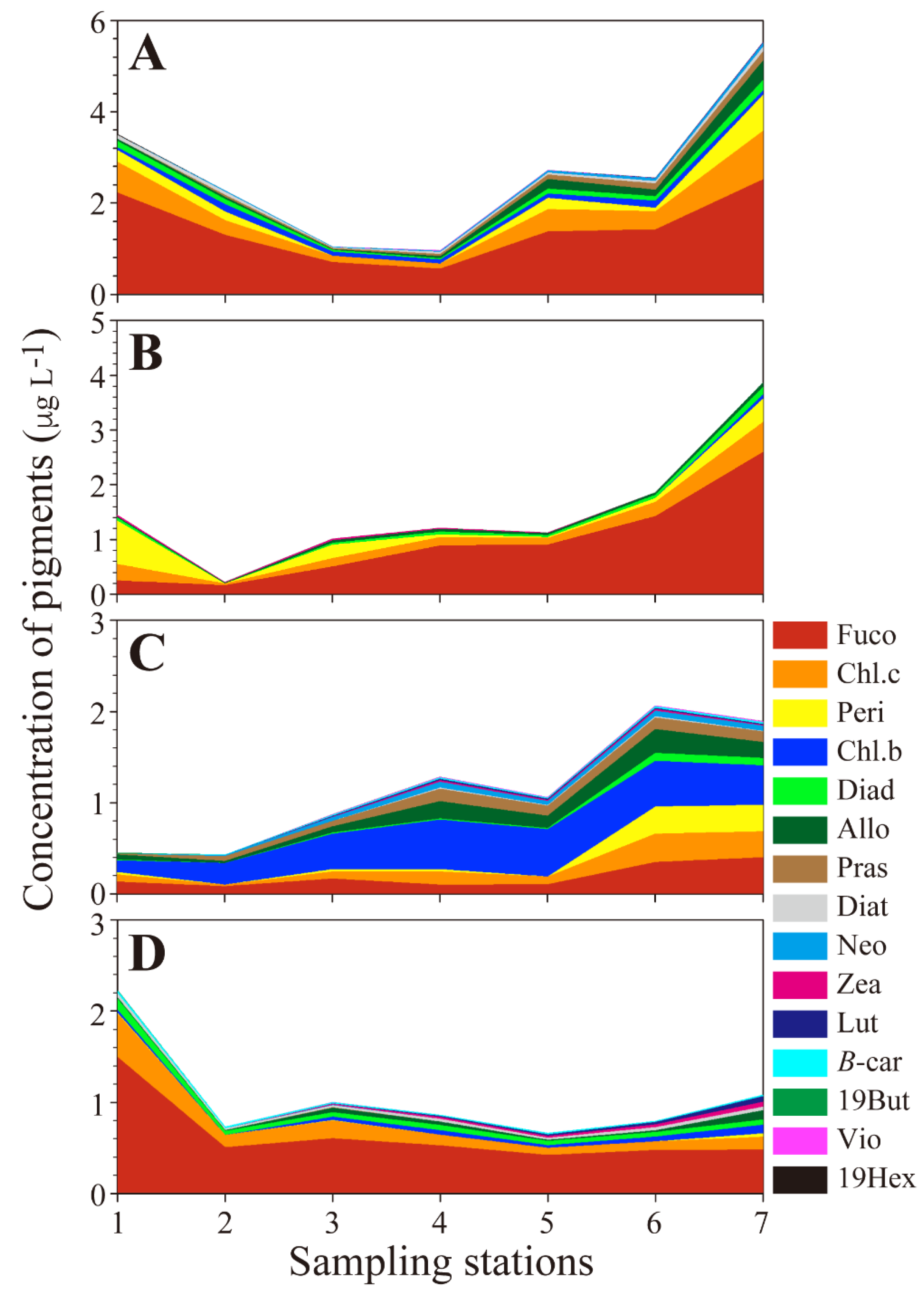
3.2. Distribution of Pigments and Phytoplankton Biomass
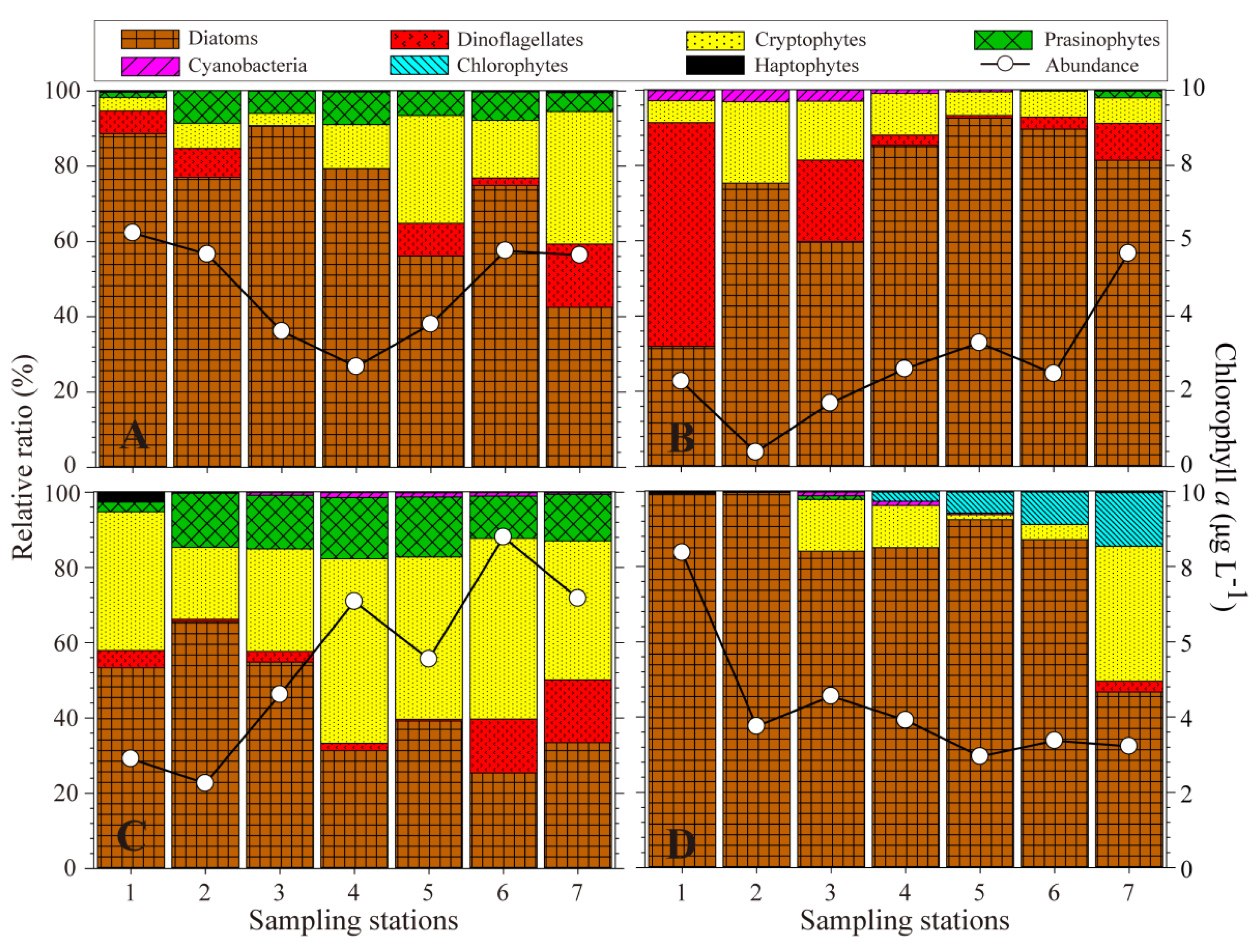
3.3. Phytoplankton Abundance and Composition Based on Microscopy and Pigment Analysis
4. Discussion
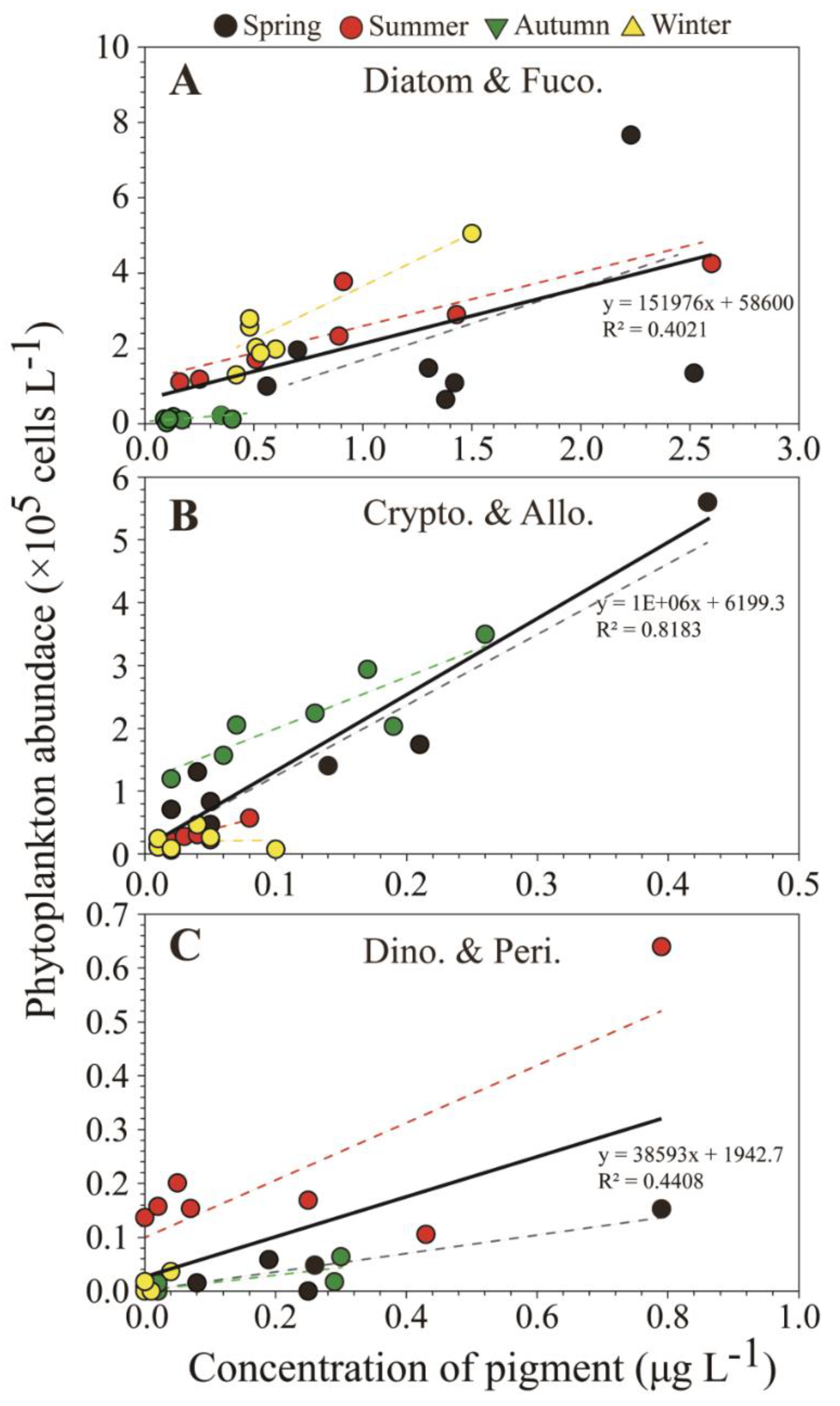
4.1. Comparison of Pigment and Microscopy Analyses in the SRE
4.2. Phytoplankton Population Dynamics in the SRE
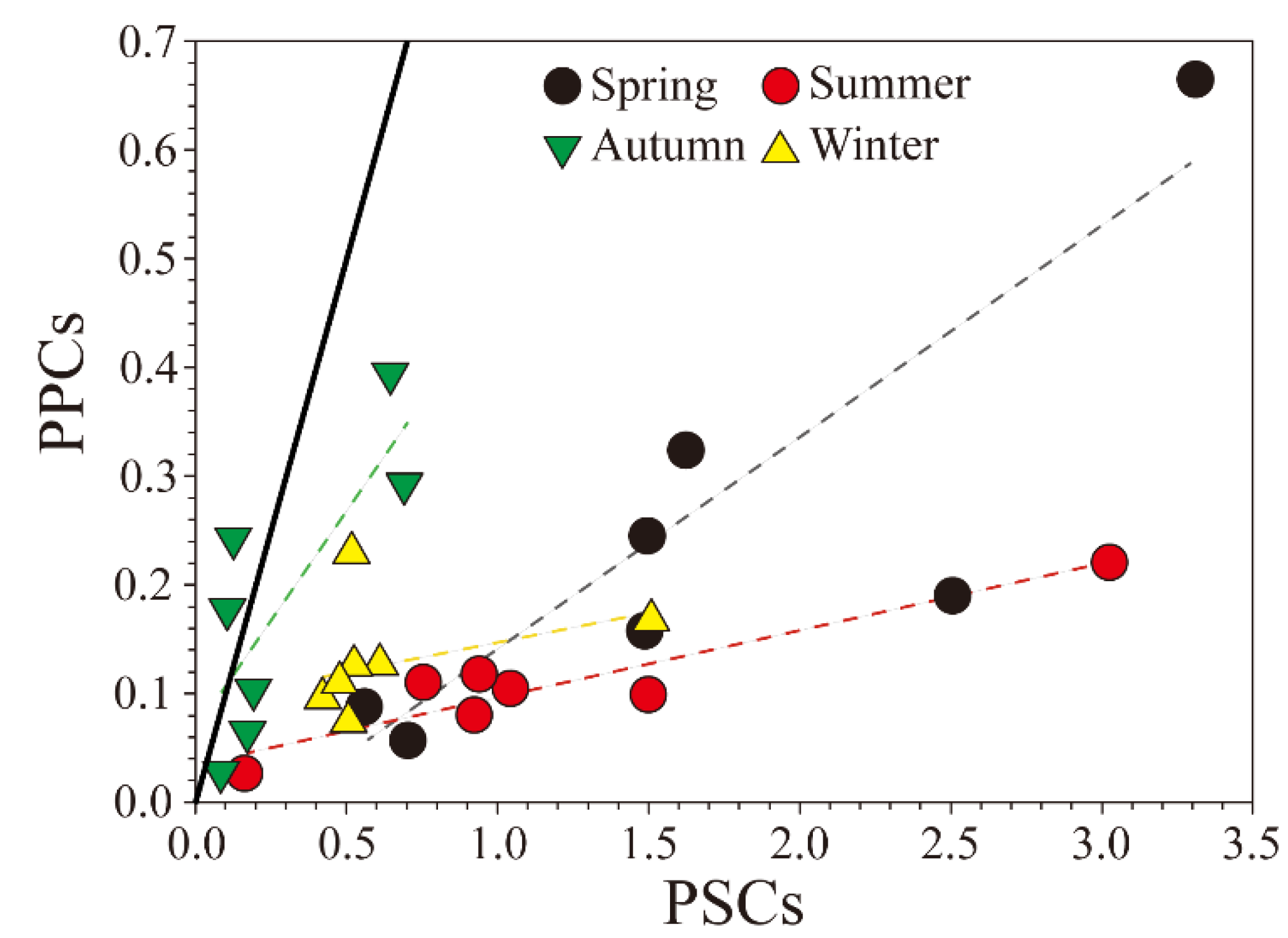
4.3. Influence of Environmental Factors on PPCs and PSCs
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cloern, J.E.; Jassby, A.D.; Schraga, T.S.; Nejad, E.; Martin, C. Ecosystem variability along the estuarine salinity gradient: Examples from long-term study of San Francisco Bay. Limnol. Oceanogr. 2017, 62, 272–291. [Google Scholar] [CrossRef] [Green Version]
- Muylaert, K.; Sabbe, K.; Vyverman, W. Spatial and temporal dynamics of phytoplankton communities in a freshwater tidal estuary (Schelde, Belgium). Estuar. Coast. Shelf. Sci. 2000, 50, 673–687. [Google Scholar] [CrossRef]
- Sheaves, M.; Baker, R.; Nagelkerken, I.; Connolly, R.M. True value of estuarine and coastal nurseries for fish: Incorporating complexity and dynamics. Estuar. Coast. 2015, 38, 401–414. [Google Scholar] [CrossRef] [Green Version]
- Jouenne, F.; Lefebvre, S.; Véron, B.; Lagadeuc, Y. Phytoplankton community structure and primary production in small intertidal estuarine-bay ecosystem (eastern English Channel, France). Mar. Biol. 2007, 151, 805–825. [Google Scholar] [CrossRef]
- Mendiguchía, C.; Moreno, C.; García-Vargas, M. Evaluation of natural and anthropogenic influences on the Guadalquivir River (Spain) by dissolved heavy metals and nutrients. Chemosphere 2007, 69, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Yegemova, S.; Kumar, R.; Abuduwaili, J.; Ma, L.; Samat, A.; Issanova, G.; Rodrigo-Comino, J. Identifying the key information and land management plans for water conservation under dry weather conditions in the Border areas of the Syr Darya River in Kazakhstan. Water 2018, 10, 1754. [Google Scholar] [CrossRef] [Green Version]
- Lehman, P.W. The influence of climate on phytoplankton community biomass in San Francisco Bay Estuary. Limnol. Oceanogr. 2000, 45, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Statham, P.J. Nutrients in estuaries—an overview and the potential impacts of climate change. Sci. Total Environ. 2012, 434, 213–227. [Google Scholar] [CrossRef]
- Wolanski, E.; Elliott, M. Estuarine Ecohydrology: An Introduction; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Aktan, Y.; Tüfekçi, V.; Tüfekçi, H.; Aykulu, G. Distribution patterns, biomass estimates and diversity of phytoplankton in Izmit Bay (Turkey). Estuar. Coast. Shelf. Sci. 2005, 64, 372–384. [Google Scholar] [CrossRef]
- Naik, R.K.; Anil, A.C.; Narale, D.D.; Chitari, R.R.; Kulkarni, V.V. Primary description of surface water phytoplankton pigment patterns in the Bay of Bengal. J. Sea. Res. 2011, 65, 435–441. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Mantoura, R.F.C.; Bjørnland, T. Data for the identification of 47 key phytoplankton pigments. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods. Monographs on Oceanographic Methodology; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO: Paris, French, 1997. [Google Scholar]
- Havskum, H.; Schlüter, L.; Scharek, R.; Berdalet, E.; Jacquet, S. Routine quantification of phytoplankton groups microscopy or pigment analyses? Mar. Ecol. Prog. Ser. 2004, 273, 31–42. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W. Pigment Markers for Phytoplankton Production Marine Organic Matter: Biomarkers, Isotopes and DNA; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Barlow, R.; Kyewalyanga, M.; Sessions, H.; Van den Berg, M.; Morris, T. Phytoplankton pigments, functional types, and absorption properties in the Delagoa and Natal Bights of the Agulhas ecosystem. Estuar. Coast. Shelf Sci. 2008, 80, 201–211. [Google Scholar] [CrossRef]
- Paerl, H.W.; Valdes, L.M.; Pinckney, J.L.; Piehler, M.F.; Dyble, J.; Moisander, P.H. Phytoplankton photopigments as indicators of estuarine and coastal eutrophication. BioScience 2003, 53, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Descy, J.P.; Higgins, H.W.; Mackey, D.J.; Hurley, J.P.; Frost, T.M. Pigment ratios and phytoplankton assessment in northern Wisconsin lakes. J. Phycol. 2000, 36, 274–286. [Google Scholar] [CrossRef]
- Seoane, S.; Garmendia, M.; Revilla, M.; Borja, Á.; Franco, J.; Orive, E.; Valencia, V. Phytoplankton pigments and epifluorescence microscopy as tools for ecological status assessment in coastal and estuarine waters, within the Water Framework Directive. Mar. Pollut. Bull. 2011, 62, 1484–1497. [Google Scholar] [CrossRef]
- Tett, P.; Carreira, C.; Mills, D.K.; Van Leeuwen, S.; Foden, J.; Bresnan, E.; Gowen, R.J. Use of a Phytoplankton Community Index to assess the health of coastal waters. ICES J. Mar. Sci. 2008, 65, 1475–1482. [Google Scholar] [CrossRef]
- Devlin, M.; Barry, J.; Painting, S.; Best, M. Extending the phytoplankton tool kit for the UK Water Framework Directive: Indicators of phytoplankton community structure. Hydrobiologia 2009, 633, 151–168. [Google Scholar] [CrossRef]
- Roy, R.; Pratihary, A.; Mangesh, G.; Naqvi, S.W.A. Spatial variation of phytoplankton pigments along the southwest coast of India. Estuar. Coast. Shelf. Sci. 2006, 69, 189–195. [Google Scholar] [CrossRef]
- Chai, C.; Jiang, T.; Cen, J.; Ge, W.; Lu, S. Phytoplankton pigments and functional community structure in relation to environmental factors in the Pearl River Estuary. Oceanologia 2016, 58, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Gibb, S.W.; Barlow, R.G.; Cummings, D.G.; Rees, N.W.; Trees, C.C.; Holligan, P.; Suggett, D. Surface phytoplankton pigment distributions in the Atlantic Ocean: An assessment of basin scale variability between 50 N and 50 S. Prog. Oceanogr. 2000, 45, 339–368. [Google Scholar] [CrossRef]
- Moreno, D.V.; Marrero, J.P.; Morales, J.; García, C.L.; Úbeda, M.G.V.; Rueda, M.J.; Llinás, O. Phytoplankton functional community structure in Argentinian continental shelf determined by HPLC pigment signatures. Estuar. Coast. Shelf. Sci. 2012, 100, 72–81. [Google Scholar] [CrossRef]
- Lee, M.; Park, B.S.; Baek, S.H. Tidal influences on biotic and abiotic factors in the Seomjin River Estuary and Gwangyang Bay, Korea. Estuaries Coast 2018, 41, 1977–1993. [Google Scholar] [CrossRef]
- Lee, D.I.; Park, C.K.; Cho, H.S. Ecological modeling for water quality management of Kwangyang Bay, Korea. J. Environ. Manag. 2005, 74, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Jang, D.; Kim, Y.H. Stratification and salt-wedge in the Seomjin river estuary under the idealized tidal influence. Ocean Sci. J. 2017, 52, 469–487. [Google Scholar] [CrossRef]
- Hong, S.H.; Kannan, N.; Yim, U.H.; Choi, J.W.; Shim, W.J. Polychlorinated biphenyls (PCBs) in a benthic ecosystem in Gwangyang Bay, South Korea. Mar. Pollut. Bull. 2011, 62, 2863–2868. [Google Scholar] [CrossRef]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Mackey, M.D.; Mackey, D.J.; Higgins, H.W.; Wright, S.W. CHEMTAX-a program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef] [Green Version]
- Riegman, R.; Kraay, G.W. Phytoplankton community structure derived from HPLC analysis of pigments in the Faroe-Shetland Channel during summer 1999: The distribution of taxonomic groups in relation to physical/chemical conditions in the photic zone. J. Plankton Res. 2001, 23, 191–205. [Google Scholar] [CrossRef]
- Zhang, J. Analysis between vegetation and environments I: CCA and DCCA. J. Shanxi Univ. 1992, 15, 182–189. [Google Scholar]
- Tester, P.A.; Geesey, M.E.; Guo, C.; Paerl, H.W.; Millie, D.F. Evaluating phytoplankton dynamics in the Newport River estuary (North Carolina, USA) by HPLC-derived pigment profiles. Mar. Ecol. Prog. Ser. 1995, 124, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.A.; Bidigare, R.R.; Keller, M.D.; Latasa, M. A comparison of HPLC pigment signatures and electron microscopic observations for oligotrophic waters of the North Atlantic and Pacific Oceans. Deep Sea Res. Part Ii Top. Stud. Oceanogr. 1996, 43, 517–537. [Google Scholar] [CrossRef]
- Suzuki, K.; Minami, C.; Liu, H.; Saino, T. Temporal and spatial patterns of chemotaxonomic algal pigments in the subarctic Pacific and the Bering Sea during the early summer of 1999. Deep Sea Res. Part Ii Top. Stud. Oceanogr. 2002, 49, 5685–5704. [Google Scholar] [CrossRef]
- Silva, A.; Mendes, C.; Palma, S.; Brotas, V. Short-time scale variation of phytoplankton succession in Lisbon bay (Portugal) as revealed by microscopy cell counts and HPLC pigment analysis. Estuar. Coast. Shelf. Sci. 2008, 79, 230–238. [Google Scholar] [CrossRef]
- Brewin, R.J.; Sathyendranath, S.; Hirata, T.; Lavender, S.J.; Barciela, R.M.; Hardman-Mountford, N.J. A three-component model of phytoplankton size class for the Atlantic Ocean. Ecol. Model. 2010, 221, 1472–1483. [Google Scholar] [CrossRef]
- Rodrıguez, F.; Pazos, Y.; Maneiro, J.; Zapata, M. Temporal variation in phytoplankton assemblages and pigment composition at a fixed station of the Rıa of Pontevedra (NW Spain). Estuar. Coast. Shelf. Sci. 2003, 58, 499–515. [Google Scholar] [CrossRef]
- Gameiro, C.; Cartaxana, P.; Brotas, V. Environmental drivers of phytoplankton distribution and composition in Tagus Estuary, Portugal. Estuar. Coast. Shelf. Sci. 2007, 75, 21–34. [Google Scholar] [CrossRef]
- Olenina, I. Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environ. Proc. 2006, 106, 1–144. [Google Scholar]
- Schlüter, L.; Møhlenberg, F.; Havskum, H.; Larsen, S. The use of phytoplankton pigments for identifying and quantifying phytoplankton groups in coastal areas: Testing the influence of light and nutrients on pigment/chlorophyll a ratios. Mar. Ecol. Prog. Ser. 2000, 192, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Lignell, R. Excretion of organic carbon by phytoplankton: Its relation to algal biomass, primary productivity and bacterial secondary productivity in the Baltic Sea. Mar. Ecol. Prog. Ser. 1990, 68, 85–99. [Google Scholar] [CrossRef]
- Huot, Y.; Babin, M.; Bruyant, F.; Grob, C.; Twardowski, M.S.; Claustre, H. Does chlorophyll a provide the best index of phytoplankton biomass for primary productivity studies? Biogeosci. Discuss. 2007, 4, 707–745. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, P.; Padisák, J.; Koschel, R.; Krienitz, L.; Gervais, F. Chlorophyll a concentration across a trophic gradient of lakes: An estimator of phytoplankton biomass? Limnologica 2008, 38, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Breton, E.; Brunet, C.; Sautour, B.; Brylinski, J.M. Annual variations of phytoplankton biomass in the Eastern English Channel: Comparison by pigment signatures and microscopic counts. J. Plankton Res. 2000, 22, 1423–1440. [Google Scholar] [CrossRef] [Green Version]
- Ediger, D.; Soydemir, N.; Kideys, A.E. Estimation of phytoplankton biomass using HPLC pigment analysis in the southwestern Black Sea. Deep Sea Res. Part Ii Top. Stud. Oceanogr. 2006, 53, 1911–1922. [Google Scholar] [CrossRef]
- Baek, S.H.; Kim, D.; Son, M.; Yun, S.M.; Kim, Y.O. Seasonal distribution of phytoplankton assemblages and nutrient-enriched bioassays as indicators of nutrient limitation of phytoplankton growth in Gwangyang Bay, Korea. Estuar. Coast. Shelf. Sci. 2015, 163, 265–278. [Google Scholar] [CrossRef]
- Park, T.G.; Lim, W.A.; Park, Y.T.; Lee, C.K.; Jeong, H.J. Economic impact, management and mitigation of red tides in Korea. Harmful algae 2013, 30, 131–143. [Google Scholar] [CrossRef]
- Baek, S.H.; Shin, K.; Son, M.; Bae, S.W.; Cho, H.; Na, D.H.; Kim, Y.O.; Kim, S.W. Algicidal effects of yellow clay and the thiazolidinedione derivative TD49 on the fish-killing dinoflagellate Cochlodinium polykrikoides in microcosm experiments. J. Appl. Phycol. 2014, 26, 2367–2378. [Google Scholar] [CrossRef]
- Tolotti, M.; Corradini, F.; Boscaini, A.; Calliari, D. Weather-driven ecology of planktonic diatoms in Lake Tovel (Trentino, Italy). Hydrobiologia 2007, 578, 147–156. [Google Scholar] [CrossRef]
- Hötzel, G.; Croome, R. Population dynamics of Aulacoseira granulata (EHR.) SIMONSON (Bacillariophyceae, Centrales), the dominant alga in the Murray River, Australia. Arch. Hydrobiol. 1996, 136, 191–215. [Google Scholar]
- Wang, C.; Li, X.; Lai, Z.; Tan, X.; Pang, S.; Yang, W. Seasonal variations of Aulacoseira granulata population abundance in the Pearl River Estuary. Estuar. Coast. Shelf Sci. 2009, 85, 585–592. [Google Scholar] [CrossRef]
- Na, J.-E.; Jung, M.-H.; Cho, I.-S.; Park, J.-H.; Hwang, K.-S.; Song, H.-J.; Lim, B.-J.; La, G.-H.; Kim, H.-W.; Lee, H.-Y. Phytoplankton community in reservoirs of Yeongsan and Seomjin River basins, Korea. Korean J. Environ. Biol. 2012, 30, 39–46. [Google Scholar]
- Yu, J.J.; Lee, H.J.; Lee, K.L.; Lyu, H.S.; Whang, J.W.; Shin, L.Y.; Chen, S.U. Relationship between distribution of the dominant phytoplankton species and water temperature in the Nakdong River, Korea. Korean J. Environ. Ecol. 2014, 47, 247–257. [Google Scholar] [CrossRef]
- Sommer, U. Comparison between steady state and non-steady state competition: Experiments with natural phytoplankton. Limnol. Oceanogr. 1985, 30, 335–346. [Google Scholar] [CrossRef]
- Klaveness, D. Biology and ecology of the Cryptophyceae: Status and challenges. Biol. Oceanog. 1989, 6, 257–270. [Google Scholar]
- Barone, R.; Naselli-Flores, L. Distribution and seasonal dynamics of Cryptomonads in Sicilian water bodies. Hydrobiologia 2003, 502, 325–329. [Google Scholar] [CrossRef]
- Baek, S.H.; Kim, D.; Kim, Y.O.; Son, M.; Kim, Y.-J.; Lee, M.; Park, B.S. Seasonal changes in abiotic environmental conditions in the Busan coastal region (South Korea) due to the Nakdong River in 2013 and effect of these changes on phytoplankton communities. Cont. Shelf Res. 2019, 175, 116–126. [Google Scholar] [CrossRef]
- Kaeriyama, H.; Katsuki, E.; Otsubo, M.; Yamada, M.; Ichimi, K.; Tada, K.; Harrison, P.J. Effects of temperature and irradiance on growth of strains belonging to seven Skeletonema species isolated from Dokai Bay, southern Japan. Eur. J. Phycol. 2011, 46, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Bidigare, R.R.; Ondrusek, M.E. Spatial and temporal variability of phytoplankton pigment distributions in the central equatorial Pacific Ocean. Deep Sea Res. Part Ii Top. Stud. Oceanogr. 1996, 43, 809–833. [Google Scholar] [CrossRef]
- Madhu, N.; Ullas, N.; Ashwini, R.; Meenu, P.; Rehitha, T.; Lallu, K. Characterization of phytoplankton pigments and functional community structure in the Gulf of Mannar and the Palk Bay using HPLC–CHEMTAX analysis. Cont. Shelf Res. 2014, 80, 79–90. [Google Scholar] [CrossRef]
- Wysocki, L.A.; Bianchi, T.S.; Powell, R.T.; Reuss, N. Spatial variability in the coupling of organic carbon, nutrients, and phytoplankton pigments in surface waters and sediments of the Mississippi River plume. Estuar. Coast. Shelf Sci. 2006, 69, 47–63. [Google Scholar] [CrossRef]
- Zhu, Z.-Y.; Ng, W.-M.; Liu, S.-M.; Zhang, J.; Chen, J.-C.; Wu, Y. Estuarine phytoplankton dynamics and shift of limiting factors: A study in the Changjiang (Yangtze River) Estuary and adjacent area. Estuar. Coast. Shelf Sci. 2009, 84, 393–401. [Google Scholar] [CrossRef]
- Cloern, J.E. Turbidity as a control on phytoplankton biomass and productivity in estuaries. Cont. Shelf Res. 1987, 7, 1367–1381. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviations | Pigment | Designation |
| Chl-a | Chlorophyll a | |
| Chl-c2 | Chlorophyll c2 | |
| Chl-b | Chlorophyll b | Chlorophytes |
| Allo | Alloxanthin | Cryptophytes |
| Diad | Diadinoxanthin | |
| Diato | Diatoxanthin | |
| Fuco | Fucoxathin | Diatoms (major) |
| Hex | 19’ Hexanoyloxy fucoxanthin | Prymnesiophytes |
| Peri | Peridinine | Dinoflagellates |
| Zea | Zeaxanthin | Cyanobacteria |
| Β-Car | β-Carotene | |
| But | 19’ Butanoyloxy fucoxanthin | Haptophytes (major) |
| Viol | Violaxanthin | |
| Lut | Lutein | |
| Neo | 9’-cis-neoxanthin | |
| Variable | Pigment sum | Formula |
| TChl-a | Total chlorophyll a | Chl a+DVChl a+Chlide a |
| PPCs | Photoprotective carotenoids | Allo+Diad+Vio+Zea+β−Car |
| PSCs | Photosynthetic carotenoids | But+Fuco+Hex+Per |
| Perid | 19butfu | Fuco | 19hexfu | Neo | Prasino | Viol | Allo | Lut | Zea | Chl b | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Prasino | 0 | 0 | 0 | 0 | 0.3768 | 0.1413 | 0.2165 | 0 | 0.0843 | 0 | 0.2807 |
| Dino | 0.7471 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Crypto | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1927 | 0 | 0 | 0 |
| Hapto_N | 0 | 0 | 0 | 1.7139 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hapto_S | 0 | 0.5076 | 0.8354 | 0.2225 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chloro | 0 | 0 | 0 | 0 | 0.0495 | 0 | 0.1185 | 0 | 0.1294 | 0.3262 | 0.0168 |
| Cyano | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.6795 | 0 |
| Diat | 0 | 0 | 1.0198 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Precipitation | Discharge | Temp. | Sal. | DO | NOx | NH4+ | DIP | DSi | |
|---|---|---|---|---|---|---|---|---|---|
| (mm/10days) | (m3 s−1) | (°C) | (psu) | (mg L−1) | (uM) | (uM) | (uM) | (uM) | |
| S1 | 17.7 | 5.5 | 20.16 | 33.05 | 6.88 | 2.99 | 3.58 | 0.41 | 14.04 |
| S2 | 21.24 | 31.34 | 6.65 | 4.32 | 5.31 | 0.65 | 20.53 | ||
| S3 | 21.45 | 31.75 | 6.32 | 4.77 | 5.86 | 0.69 | 20.18 | ||
| S4 | 22.36 | 27.32 | 6.92 | 15.54 | 6.49 | 0.88 | 36.96 | ||
| S5 | 23.20 | 20.81 | 6.90 | 28.89 | 5.72 | 0.86 | 53.05 | ||
| S6 | 23.50 | 18.44 | 6.58 | 35.54 | 5.60 | 0.87 | 64.08 | ||
| S7 | 23.76 | 12.74 | 7.05 | 39.89 | 3.74 | 0.74 | 64.61 | ||
| Sm1 | 12.8 | 17.5 | 23.25 | 32.03 | 5.74 | 0.96 | 0.39 | 0.35 | 17.63 |
| Sm2 | 25.15 | 30.03 | 6.44 | 1.08 | 2.27 | 0.46 | 18.22 | ||
| Sm3 | 25.68 | 28.74 | 5.96 | 1.33 | 3.58 | 0.53 | 19.49 | ||
| Sm4 | 26.52 | 25.30 | 6.33 | 0.84 | 3.81 | 0.41 | 21.54 | ||
| Sm5 | 26.52 | 21.57 | 6.70 | 2.95 | 4.33 | 0.33 | 32.23 | ||
| Sm6 | 26.60 | 17.08 | 6.76 | 7.15 | 5.27 | 0.34 | 45.88 | ||
| Sm7 | 26.91 | 17.28 | 6.38 | 5.70 | 5.53 | 0.34 | 46.07 | ||
| A1 | 15.4 | 38.5 | 18.23 | 31.76 | 7.58 | 2.34 | 5.69 | 0.94 | 14.48 |
| A2 | 18.05 | 29.88 | 7.90 | 6.07 | 7.30 | 1.06 | 25.13 | ||
| A3 | 17.69 | 27.81 | 7.88 | 6.70 | 7.40 | 1.08 | 24.27 | ||
| A4 | 16.95 | 22.65 | 7.64 | 12.06 | 7.03 | 1.10 | 36.14 | ||
| A5 | 16.45 | 15.30 | 8.20 | 22.78 | 6.15 | 0.98 | 55.81 | ||
| A6 | 16.27 | 14.67 | 8.33 | 23.49 | 5.77 | 0.77 | 48.61 | ||
| A7 | 15.90 | 9.90 | 8.92 | 28.75 | 5.23 | 0.84 | 62.48 | ||
| W1 | 46.9 | 69.0 | 7.88 | 31.71 | 10.26 | 2.71 | 0.88 | 0.31 | 8.17 |
| W2 | 8.00 | 30.25 | 10.11 | 7.77 | 1.90 | 0.35 | 16.95 | ||
| W3 | 7.96 | 27.68 | 10.30 | 10.41 | 2.09 | 0.36 | 20.23 | ||
| W4 | 7.25 | 22.20 | 10.88 | 28.37 | 2.37 | 0.28 | 45.98 | ||
| W5 | 6.88 | 14.69 | 11.47 | 44.51 | 1.86 | 0.26 | 62.95 | ||
| W6 | 7.11 | 15.20 | 11.50 | 45.17 | 2.01 | 0.27 | 63.89 | ||
| W7 | 6.36 | 9.80 | 11.80 | 58.48 | 1.58 | 0.23 | 74.19 |
| Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Station 6 | Station 7 | ||
|---|---|---|---|---|---|---|---|---|
| Spring | Total abundance | 9.05 | 2.01 | 2.68 | 1.85 | 2.40 | 2.50 | 7.11 |
| Diatoms (%) | 84.70 | 73.86 | 72.98 | 53.74 | 27.06 | 43.29 | 18.95 | |
| Dinoflagellates (%) | 0.53 | 2.90 | 0.31 | 0.93 | 0.00 | 0.61 | 2.15 | |
| Cryptophytes (%) | 14.41 | 23.24 | 26.40 | 44.86 | 72.61 | 56.10 | 78.78 | |
| Summer | Total abundance | 2.72 | 0.69 | 1.07 | 2.17 | 4.83 | 4.81 | 7.15 |
| Diatoms (%) | 62.86 | 76.73 | 81.12 | 84.24 | 89.28 | 85.89 | 83.77 | |
| Dinoflagellates (%) | 33.97 | 9.43 | 8.03 | 7.27 | 3.72 | 4.53 | 2.07 | |
| Cryptophytes (%) | 3.17 | 13.84 | 10.84 | 8.18 | 6.56 | 9.07 | 11.23 | |
| Autumn | Total abundance | 1.80 | 1.34 | 2.16 | 2.12 | 2.37 | 3.83 | 0.93 |
| Diatoms (%) | 10.19 | 9.52 | 4.92 | 1.33 | 5.41 | 5.81 | 3.89 | |
| Dinoflagellates (%) | 1.85 | 0.00 | 0.00 | 0.67 | 0.00 | 1.66 | 0.56 | |
| Cryptophytes (%) | 87.04 | 89.29 | 95.08 | 96.00 | 94.59 | 91.29 | 94.44 | |
| Winter | Total abundance | 5.22 | 2.20 | 2.31 | 2.36 | 1.56 | 2.67 | 2.93 |
| Diatoms (%) | 96.71 | 91.94 | 85.71 | 79.53 | 83.33 | 96.08 | 95.06 | |
| Dinoflagellates (%) | 0.00 | 0.81 | 0.00 | 0.00 | 0.00 | 0.00 | 1.23 | |
| Cryptophytes (%) | 2.74 | 4.84 | 11.43 | 19.69 | 15.56 | 3.27 | 2.47 |
| Station 1 | Station 2 | Station 3 | Station 4 | Station 5 | Station 6 | Station 7 | ||
|---|---|---|---|---|---|---|---|---|
| Spring | Chaetoceros | +++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Skeletonema | ++ | + | + | ++ | + | ++ | ++ | |
| Thalassionema | +++ | ++ | +++ | ++ | ++ | + | + | |
| Cryptomonas | +++ | ++ | ++ | ++ | +++ | +++ | +++ | |
| Summer | Aulacoseira | + | ++ | ++ | +++ | +++ | +++ | |
| Chaetoceros | ++ | ++ | ++ | ++ | ++ | ++ | ++ | |
| Skeletonema | ++ | + | ++ | ++ | ++ | ++ | ++ | |
| Cryptomonas | + | ++ | ++ | ++ | ++ | ++ | ++ | |
| Autumn | Amphora | + | + | + | ||||
| Navicular | ++ | + | + | + | + | + | ||
| Nitzschia | + | + | + | + | + | |||
| Cryptomonas | +++ | +++ | +++ | +++ | +++ | +++ | +++ | |
| Winter | Chaetoceros | +++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Skeletonema | +++ | ++ | +++ | +++ | ++ | +++ | +++ | |
| Thalassiosira | ++ | ++ | + | ++ | ++ | ++ | ++ | |
| Cryptomonas | ++ | ++ | ++ | ++ | ++ | + | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.; Won, N.-I.; Baek, S.H. Comparison of HPLC Pigment Analysis and Microscopy in Phytoplankton Assessment in the Seomjin River Estuary, Korea. Sustainability 2020, 12, 1675. https://doi.org/10.3390/su12041675
Lee M, Won N-I, Baek SH. Comparison of HPLC Pigment Analysis and Microscopy in Phytoplankton Assessment in the Seomjin River Estuary, Korea. Sustainability. 2020; 12(4):1675. https://doi.org/10.3390/su12041675
Chicago/Turabian StyleLee, Minji, Nam-Il Won, and Seung Ho Baek. 2020. "Comparison of HPLC Pigment Analysis and Microscopy in Phytoplankton Assessment in the Seomjin River Estuary, Korea" Sustainability 12, no. 4: 1675. https://doi.org/10.3390/su12041675
APA StyleLee, M., Won, N. -I., & Baek, S. H. (2020). Comparison of HPLC Pigment Analysis and Microscopy in Phytoplankton Assessment in the Seomjin River Estuary, Korea. Sustainability, 12(4), 1675. https://doi.org/10.3390/su12041675