The Loss of Landscape Ecological Functionality in the Barcelona Province (1956–2009): Could Land-Use History Involve a Legacy for Current Biodiversity?


Abstract
:1. Introduction
2. Case Study and Methods
2.1. HANPP as a Measure of Farming Ecology Disturbance
2.2. Landscape Ecology Metrics to Assess Land Cover Diversity and Fragmentation
2.3. Landscape Ecology Metrics to Assess Ecological Connectivity
2.4. Statistical Analysis of the Relationship between Farming Disturbance and Landscape Ecology Metrics in 1956 and 2009 with Total Species Richness in 2009
3. Results
4. Discussion
4.1. Land Use and Cover Change (LUCC) from 1956 to 2009
4.2. Trends in the Net Primary Production and its Human Appropriation from 1956 to 2009
4.3. The Loss of Landscape Complex Mosaics from 1956 to 2009
4.4. Statistical Impact on Total Species Richness in 2009 of Landscape Homogenization and the Increase of Larger Forest Units from 1956 to 2009
4.5. The Impact of Past Conservation Policies on Landscape Functioning
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pagnutti, C.; Bauch, C.T.; Anan, M. Outlook on a worldwide forest transition. PLoS ONE 2013, 8, e75890. [Google Scholar] [CrossRef] [PubMed]
- Culas, R.J. REDD and forest transition: Tunneling through the environmental Kuznets curve. Ecol. Econ. 2012, 79, 44–51. [Google Scholar] [CrossRef]
- Barbier, E.B.; Burgess, J.C.; Grainger, A. The forest transition: Towards a more comprehensive theoretical framework. Land Use Policy 2010, 27, 98–107. [Google Scholar] [CrossRef]
- Rudel, T.K.; Coomes, O.T.; Moran, E.; Achard, F.; Angelsen, A.; Xu, J.; Lambin, E. Forest transitions: Towards a global understanding of land use change. Glob. Environ. Chang. 2005, 15, 23–31. [Google Scholar] [CrossRef]
- Rudel, T.K.; Schneider, L.; Uriarte, M. Forest transitions: An introduction. Land Use Policy 2010, 27, 95–97. [Google Scholar] [CrossRef]
- EEA. Landscapes in Transition. An Account of 25 Years of Land Cover Change in Europe; European Environmental Agency Report Number 10/2017; Office for Official Publications of the European Communities: Luxembourg, 2017; Available online: https://www.eea.europa.eu/publications/landscapes-in-transition (accessed on 27 January 2020).
- Gerard, F.; Petit, S.; Smith, G.; Thomson, A.; Brown, N.; Manchester, S.; Wadsworth, R.; Bugar, G.; Halada, L.; Bezak, P.; et al. Land cover change in Europe between 1950 and 2000 determined employing aerial photography. Prog. Phys. Geog. 2010, 34, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, C.; Beilin, R.; Folke, C.; Lindborg, R. Farmland abandonment: Threat or opportunity for biodiversity conservation? A global review. Front. Ecol. Environ. 2014, 12, 288–296. [Google Scholar] [CrossRef]
- Plieninger, T.; Hui, C.; Gaertner, M.; Huntsinger, L. The Impact of Land Abandonment on Species Richness and Abundance in the Mediterranean Basin: A Meta-Analysis. PLoS ONE 2014, 9, e9835. [Google Scholar] [CrossRef]
- Navarro, L.M.; Pereira, H.M. (Eds.) Rewilding Abandoned Landscapes in Europe; Springer Open: Cham, Switzerland, 2015. [Google Scholar]
- Cervera, T.; Pino, J.; Marull, J.; Padró, R.; Tello, E. Understanding the long-term dynamics of forest transition: From deforestation to afforestation in a Mediterranean landscape (Catalonia, 1868–2005). Land Use Policy 2016, 80, 318–331. [Google Scholar] [CrossRef]
- Otero, I.; Boada, M.; Tàbara, J.D. Social-ecological heritage and the conservation of Mediterranean landscapes under global change. A case study in Olzinelles (Catalonia). Land Use Policy 2013, 30, 25–37. [Google Scholar] [CrossRef]
- Otero, I.; Marull, J.; Tello, E.; Diana, G.L.; Pons, M.; Coll, F.; Boada, M. Land abandonment, landscape, and biodiversity: Questioning the restorative character of the forest transition in the Mediterranean. Ecol. Soc. 2015, 20, 7. [Google Scholar] [CrossRef] [Green Version]
- Tello, E.; Valldeperas, E.; Ollés, N.; Marull, J.; Coll, F.; Warde, P.; Wilcox, P.T. Looking backwards into a Mediterranean edge environment: Landscape changes in El Congost Valley (Catalonia) 1850–2005. Environ. Hist.-UK 2014, 20, 347–384. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Hartel, T.; Kuemmerle, T. Conservation policy in traditional farming landscapes. Conserv. Lett. 2012, 5, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Perfecto, I.; Vandermeer, J. The agroecological matrix as alternative to the land-sparing/agriculture intensification model. Proc. Natl. Acad. Sci. USA 2010, 107, 5786–5791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matson, P.A.; Vitousek, P.M. Agricultural Intensification: Will Land Spared from Farming be Land Spared for Nature? Conserv. Biol. 2006, 20, 709–710. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.A.; Parton, W.J.; Power, A.G.; Swift, M.J. Agricultural Intensification and Ecosystem Properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [Green Version]
- Gliessman, S.R.; Engles, E.W. Agroecology: The Ecology of Sustainable Food Systems, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 150, 53–59. [Google Scholar] [CrossRef]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes—Eigth hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef]
- Tilman, D. Competition and Biodiversity in Spatially Structured Habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Roxburgh, S.H.; Shea, K.; Wilson, J.B. The intermediate disturbance hypothesis: Patch dynamics and mechanisms of species coexistence. Ecology 2004, 85, 359–371. [Google Scholar] [CrossRef]
- Harper, K.A.; MacDonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.A.R.; Jaiteh, M.S.; Esseen, P.A.; et al. Edge Influence on Forest Structure and Composition in Fragmented Landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Loreau, M.; Mouquet, N.; Gonzalez, A. Biodiversity as spatial insurance in heterogeneous landscapes. Proc. Natl. Acad. Sci. USA 2003, 100, 12765–12770. [Google Scholar] [CrossRef] [Green Version]
- Marull, J.; Tello, E.; Bagaria, R.; Font, X.; Cattaneo, C.; Pino, J. Exploring the links between social metabolism and biodiversity distribution across landscape gradients: A regional-scale contribution to the land-sharing versus land-sparing debate. Sci. Total Environ. 2018, 619–620, 1272–1285. [Google Scholar] [CrossRef] [Green Version]
- Marull, J.; Herrando, S.; Brotons, L.; Melero, Y.; Pino, J.; Cattaneo, C.; Pons, M.; Llobet, J.; Tello, E. Building on Margalef: Testing the links between landscape structure, energy and information flows driven by farming and biodiversity. Sci. Total Environ. 2019, 674, 603–614. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Mooney, H.A.; Agard, J.; Capistrano, D.; DeFries, R.S.; Díaz, S.; Dietz, T.; Duraiappah, A.K.; Oteng-Yeboah, A.; Pereira, H.M.; et al. Science for managing ecosystem services: Beyond the Millennium Ecosystem Assessment. Proc. Natl. Acad. Sci. USA 2009, 106, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.M.; Levine, S.H. A model for behavioral regulation of metapopulation dynamics. Ecol. Model. 2003, 183, 411–423. [Google Scholar] [CrossRef]
- Isbell, F.; Tilman, D.; Polasky, S.; Loreau, M. The biodiversity-dependent ecosystem service debt. Ecol. Lett. 2015, 18, 119–134. [Google Scholar] [CrossRef]
- Başnou, C.; Álvarez, E.; Bagaria, G.; Guardiola, M.; Isern, R.; Vicente, P.; Pino, J. Spatial Patterns of Land Use Changes Across a Mediterranean Metropolitan Landscape: Implications for Biodiversity Management. Environ. Manag. 2013, 52, 971–980. [Google Scholar] [CrossRef]
- Agnoletti, M. Rural landscapes, nature conservation and culture. Some notes on research trends and management approaches from a (southern) European perspective. Landsc. Urban Plan. 2014, 126, 66–73. [Google Scholar] [CrossRef]
- Phalan, B.; Onial, M.; Balmford, A.; Green, R.E. Reconciling Food Production and Biodiversity Conservation: Land Sharing and Land Sparing Compared. Science 2011, 333, 1289–1291. [Google Scholar] [CrossRef]
- Winqvist, C.; Bengtsson, J.; Aavik, T.; Berendse, F.; Clement, L.W.; Eggers, S.; Fischer, C.; Flohre, A.; Geiger, F.; Liira, J.; et al. Mixed effects of organic farming and landscape complexity on farmland biodiversity and biological control potential across Europe. J. Appl. Ecol. 2011, 48, 570–579. [Google Scholar] [CrossRef]
- Geri, F.; Amici, V.; Rocchini, D. Human activity impact on the heterogeneity of a Mediterranean landscape. Appl. Geogr. 2010, 30, 370–379. [Google Scholar] [CrossRef]
- Swift, M.J.; Izac, A.-M.N.; van Noordwijk, M. Biodiversity and ecosystems services in agricultural landscapes—Are we asking the right questions? Agric. Ecosyst. Environ. 2004, 104, 113–134. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Altieri, M. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Inger, R.; Gregory, R.; Duffy, J.P.; Stott, I.; Voříšek, P.; Gaston, K.J. Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecol. Lett. 2015, 18, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Hamer, T.L.; Flather, C.H.; Noon, B.R. Factors associated with grassland bird species richness: The relative roles of grassland area, landscape structure, and prey. Landsc. Ecol. 2006, 21, 569–583. [Google Scholar] [CrossRef]
- Donald, P.F.; Green, R.E.; Heath, M.F. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc. R. Soc. Lond. B 2001, 268, 25–29. [Google Scholar] [CrossRef]
- Marull, J.; Tello, E.; Wilcox, P.T.; Coll, F.; Pons, M.; Warde, P.; Valldeperas, N.; Ollés, A. Recovering the land-use history behind a Mediterranean edge environment: The importance of agroforestry systems in biological conservation. Appl. Geogr. 2014, 54, 1–17. [Google Scholar] [CrossRef]
- Marull, J.; Otero, I.; Stefanescu, C.; Tello, E.; Miralles, M.; Coll, F.; Pons, M.; Diana, G.L. Exploring the links between forest transition and landscape changes in the Mediterranean: Can forest recovery lead to lower landscape quality? Agrofor. Syst. 2015, 89, 705–719. [Google Scholar] [CrossRef] [Green Version]
- Van der Ploeg, J.D. Peasants and the Art of Farming: A Chayanovian Manifesto; Practical Action Publishing: Rugby, UK, 2014. [Google Scholar]
- Fischer, J.; Brosi, B.; Daily, G.C.; Ehrlich, P.R.; Goldman, R.; Goldstein, J.; Lindenmayer, D.B.; Manning, A.D.; Mooney, H.A.; Pejchar, L.; et al. Should agricultural policies encourage land sparing or wildlife-friendly farming? Front. Ecol. Environ. 2008, 6, 380–385. [Google Scholar] [CrossRef]
- Shea, K.; Roxburgh, S.H.; Rauschert, E.S.L. Moving from pattern to process: Coexistence mechanisms under intermediate disturbance regimes. Ecol. Lett. 2004, 7, 491–508. [Google Scholar] [CrossRef]
- Pierce, S. Implications for biodiversity conservation of the lack of consensus regarding the humped-back model of species richness and biomass production. Funct. Ecol. 2014, 28, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Marull, J.; Delgadillo, O.; Cattaneo, C.; La Rota, M.J.; Krausmann, F. Socioecological transition in the Cauca river valley, Colombia (1943–2010): Towards an energy–landscape integrated analysis. Reg. Environ. Chang. 2018, 18, 1073–1087. [Google Scholar] [CrossRef]
- Marull, J.; Font, C.; Tello, E.; Fullana, N.; Domene, E.; Pons, M.; Galán, E. Towards an energy-landscape integrated analysis? Exploring the links between socio-metabolic disturbance and landscape ecology performance (Mallorca Island, Spain, 1956–2011). Landsc. Ecol. 2016, 31, 317–336. [Google Scholar] [CrossRef] [Green Version]
- Marull, J.; Font, C.; Padró, R.; Tello, E.; Panazzolo, A. Energy–landscape integrated analysis: A proposal for measuring complexity in internal agroecosystem processes (Barcelona metropolitan region, 1860–2000). Ecol. Indic. 2016, 66, 30–46. [Google Scholar] [CrossRef] [Green Version]
- Marull, J.; Tello, E.; Fullana, N.; Murray, I.; Jover, G.; Font, C.; Coll, F.; Domene, E.; Leoni, V.; Decolli, T. Long-term bio-cultural heritage: Exploring the intermediate disturbance hypothesis in agro-ecological landscapes (Mallorca, c. 1850–2012). Biodivers. Conserv. 2015, 24, 3217–3251. [Google Scholar] [CrossRef] [Green Version]
- Marull, J.; Pino, J.; Tello, E.; Cordobilla, M.J. Social metabolism, landscape change and land-use planning in the Barcelona Metropolitan Region. Land Use Policy 2010, 27, 497–510. [Google Scholar] [CrossRef]
- Foster, D.; Swanson, F.; Aber, J.; Burke, I.; Brokaw, N.; Tilman, D.; Knapp, A. The importance of land-use legacies to ecology and conservation. Bioscience 2003, 53, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Basnou, C.; Vicente, P.; Espelta, J.M.; Pino, J. Of niche differentiation, dispersal ability and historical legacies: What drives woody community assembly in recent Mediterranean forests? Oikos 2016, 125, 107–116. [Google Scholar] [CrossRef]
- Le Provost, G.; Badenhausser, I.; Le Bagousse-Pinguet, Y.; Clough, Y.; Henckel, L.; Violle, C.; Bretagnolle, V.; Roncoroni, M.; Manning, P.; Gross, N. Land-use history impacts functional diversity across multiple trophic groups. Proc. Natl. Acad. Sci. USA 2020. [Google Scholar] [CrossRef]
- Santos, K.C.; Pino, J.; Rodà, F.; Guirado, M.; Ribas, J. Beyond the reserves: The role of non-protected rural areas for avifauna conservation in the area of Barcelona (NE of Spain). Landsc. Urban Plan. 2008, 84, 140–151. [Google Scholar] [CrossRef]
- Hilbe, J.M. Negative Binomial Regression; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Haberl, H.; Erb, K.H.; Krausmann, F. Human Appropriation of Net Primary Production: Patterns, Trends, and Planetary Boundaries. Annu. Rev. Env. Resour. 2014, 39, 363–391. [Google Scholar] [CrossRef]
- Guzmán, G.; Aguilera, E.; Soto, D.; Cid, A.; Infante, J.; Ruiz, R.G.; Herrera, A.; Villa, I.; de Molina, M.G. Methodology and Conversion Factors to Estimate the Net Primary Productivity of Historical and Contemporary Agroecosystems. Sociedad Española de Historia Agraria, Working Paper DT-SEHA 1407. 2014. Available online: http://repositori.uji.es/xmlui/bitstream/handle/10234/91670/DT-SEHA%201407.pdf?sequence=3574 (accessed on 27 January 2020).
- Jaeger, J.A.G. Landscape division, splitting index, and effective mesh size: New measures of landscape fragmentation. Landsc. Ecol. 2000, 15, 115–130. [Google Scholar] [CrossRef]
- Marull, J.; Mallarach, J.M. A GIS methodology for assessing ecological connectivity: Application to the Barcelona Metropolitan Area. Landsc. Urban Plan. 2005, 71, 243–262. [Google Scholar] [CrossRef]
- Moser, B.; Jaeger, J.A.G.; Tappeiner, U.; Tasser, E.; Eiselt, B. Modification of the effective mesh size for measuring landscape fragmentation to solve the boundary problem. Landsc. Ecol. 2007, 22, 447–459. [Google Scholar] [CrossRef]
- Pino, J.; Marull, J. Ecological networks: Are they enough for connectivity conservation? A case study in the Barcelona Metropolitan Region (NE Spain). Land Use Policy 2012, 29, 684–690. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Parcerisas, L.; Marull, J.; Pino, J.; Tello, E.; Coll, F.; Basnou, C. Land use changes, landscape ecology and their socioeconomic driving forces in the Spanish Mediterranean coast (El Maresme County, 1850–2005). Environ. Sci. Policy 2012, 23, 123–132. [Google Scholar] [CrossRef]



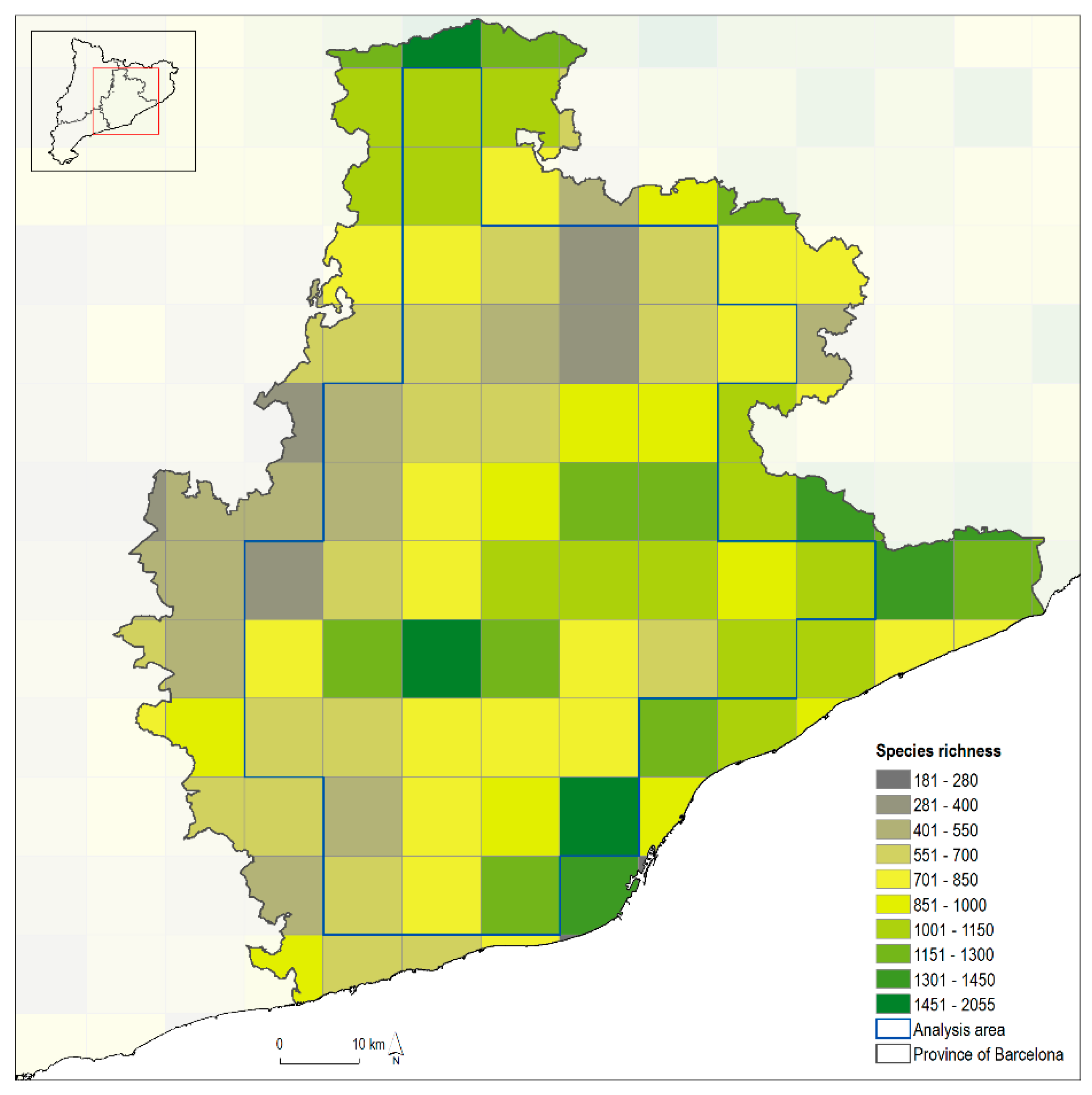{kind=link}
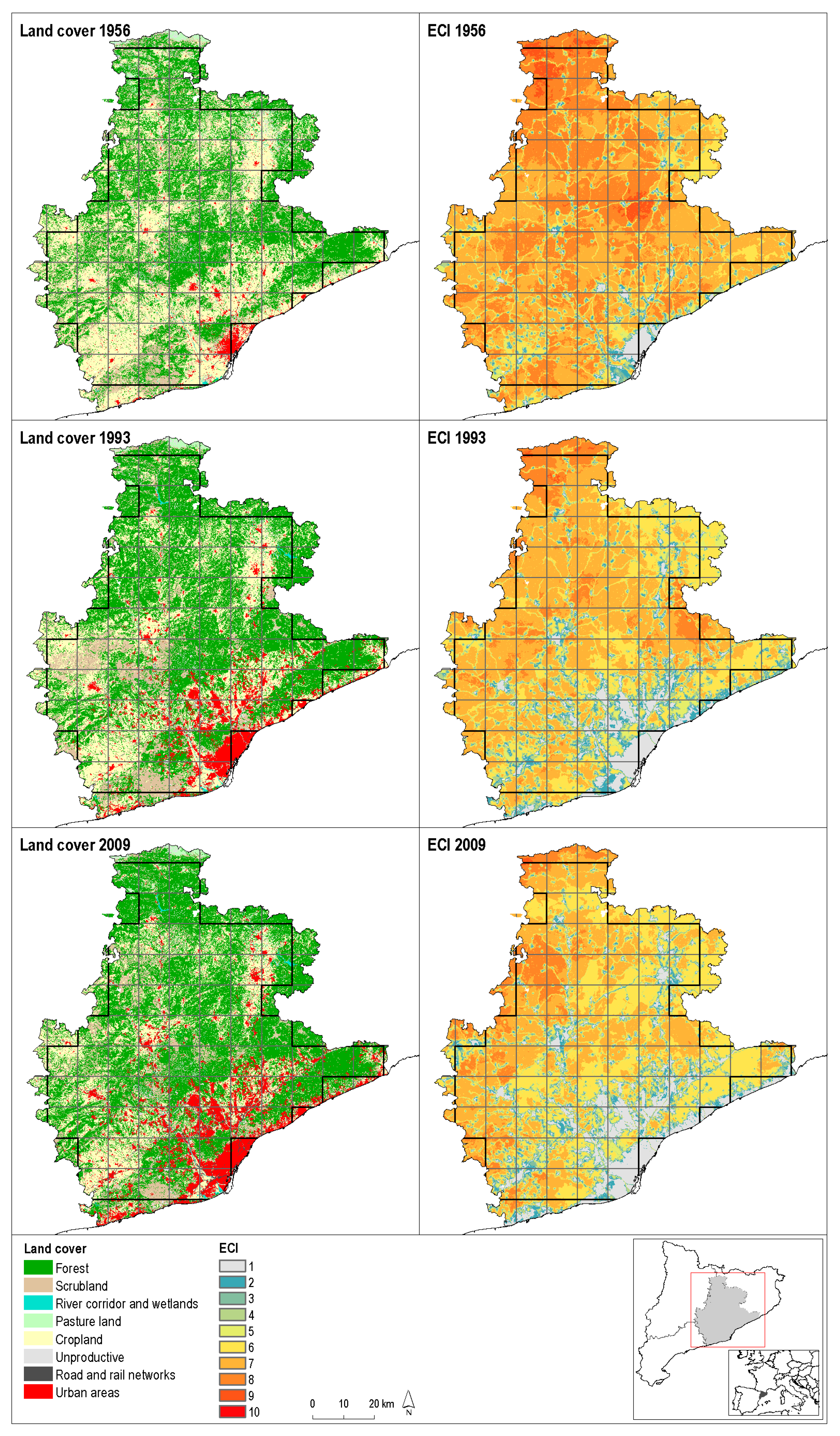{kind=link}
| Land Covers | 1956 | 1993 | 2009 | |||||
|---|---|---|---|---|---|---|---|---|
| ha | % | ha | % | 1956 = 100 | Ha | % | 1956 = 100 | |
| Forest | 195,526.4 | 40.7 | 223,060.6 | 46.5 | 114.1 | 233,357.3 | 48.6 | 119.3 |
| Scrubland and pastures | 88,710.9 | 18.5 | 69,751.0 | 14.5 | 78.6 | 68,691.5 | 14.3 | 77.4 |
| River corridor and wetlands | 1,759.7 | 0.4 | 1,755.3 | 0.4 | 99.7 | 2,097.0 | 0.4 | 119.2 |
| Cropland | 173,140.0 | 36.1 | 135,288.2 | 28.2 | 78.1 | 104,359.8 | 21.7 | 60.3 |
| Unproductive | 8,246.7 | 1.7 | 10,688.9 | 2.2 | 129.6 | 10,106.3 | 2.1 | 122.5 |
| Road and rail networks | 2,246.6 | 0.5 | 3,509.4 | 0.7 | 156.2 | 7,487.2 | 1.6 | 333.3 |
| Urban area | 10,369.7 | 2.2 | 35,946.7 | 7.5 | 346.7 | 53,900.8 | 11.2 | 519.8 |
| Total | 480,000.0 | 100.00 | 480,000.2 | 100.0 | 480,000.0 | 100.0 | ||
| Landscape Ecology Metric and HANPP Values | 1956 (A) | 1993 (B) | 2009 (C) |
|---|---|---|---|
| Polygon Density—PD (number of polygons) | 3,081.19 - | - | 3,786.98 A |
| Edge Density—ED (km) | 31.51 - | - | 31.58 - |
| Largest Patch Index—LPI (ha) | 1,964.82 - | - | 2,096.68 - |
| Effective Mesh Size—EMS (km2) | 256.49 C | - | 121.51 - |
| Shannon-Wiener Index (H’) | 0.589 C | - | 0.504 - |
| Ecological Connectivity Index—ECI | 6.81 BC | 5.65 - | 5.23 - |
| Net Primary Production actual—NPPact (TM C year−1 ha−1) | 69.08 - | 92.69 A | 86.04 A |
| Net Primary Production harvested—NPPharv (TM C year−1 ha−1) | 24.97 - | 35.82 AC | 25.73 - |
| Human Appropriation of NPP—HANPP (%) | 60.87 BC | 48.78 - | 50.42 - |
| Total Biodiversity | IRR | Std. Err. | Z | P > |z| |
|---|---|---|---|---|
| Δ HANPP | 1.132 | 0.064 | 2.20 | 0.028 (**) |
| Δ H’ | 0.802 | 0.047 | −3.73 | 0.000 (*) |
| Δ LPI | 1.127 | 0.055 | 2.45 | 0.014 (**) |
| Δ PD | 1.197 | 0.130 | 1.65 | 0.099 |
| Δ ED | 0.996 | 0.151 | −0.03 | 0.977 |
| Δ EMS | 1.058 | 0.038 | 1.60 | 0.111 |
| Δ ECI | 0.990 | 0.045 | −0.22 | 0.827 |
| cons. | 813.297 | 27.236 | 200.10 | 0.000 |
| Surface Area under Conservation Figures | Pearson Correlation | P > |z| |
|---|---|---|
| Δ HANPP | −0.029 | 0.845 |
| Δ H’ | −0.195 | 0.184 |
| Δ LPI | 0.370 | 0.010 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tello, E.; Marull, J.; Padró, R.; Cattaneo, C.; Coll, F. The Loss of Landscape Ecological Functionality in the Barcelona Province (1956–2009): Could Land-Use History Involve a Legacy for Current Biodiversity? Sustainability 2020, 12, 2238. https://doi.org/10.3390/su12062238
Tello E, Marull J, Padró R, Cattaneo C, Coll F. The Loss of Landscape Ecological Functionality in the Barcelona Province (1956–2009): Could Land-Use History Involve a Legacy for Current Biodiversity? Sustainability. 2020; 12(6):2238. https://doi.org/10.3390/su12062238
Chicago/Turabian StyleTello, Enric, Joan Marull, Roc Padró, Claudio Cattaneo, and Francesc Coll. 2020. "The Loss of Landscape Ecological Functionality in the Barcelona Province (1956–2009): Could Land-Use History Involve a Legacy for Current Biodiversity?" Sustainability 12, no. 6: 2238. https://doi.org/10.3390/su12062238
APA StyleTello, E., Marull, J., Padró, R., Cattaneo, C., & Coll, F. (2020). The Loss of Landscape Ecological Functionality in the Barcelona Province (1956–2009): Could Land-Use History Involve a Legacy for Current Biodiversity? Sustainability, 12(6), 2238. https://doi.org/10.3390/su12062238