Natural Protected Areas as Providers of Ecological Connectivity in the Landscape: The Case of the Iberian Lynx

 and
and
Abstract
:1. Introduction
2. Materials and Methods
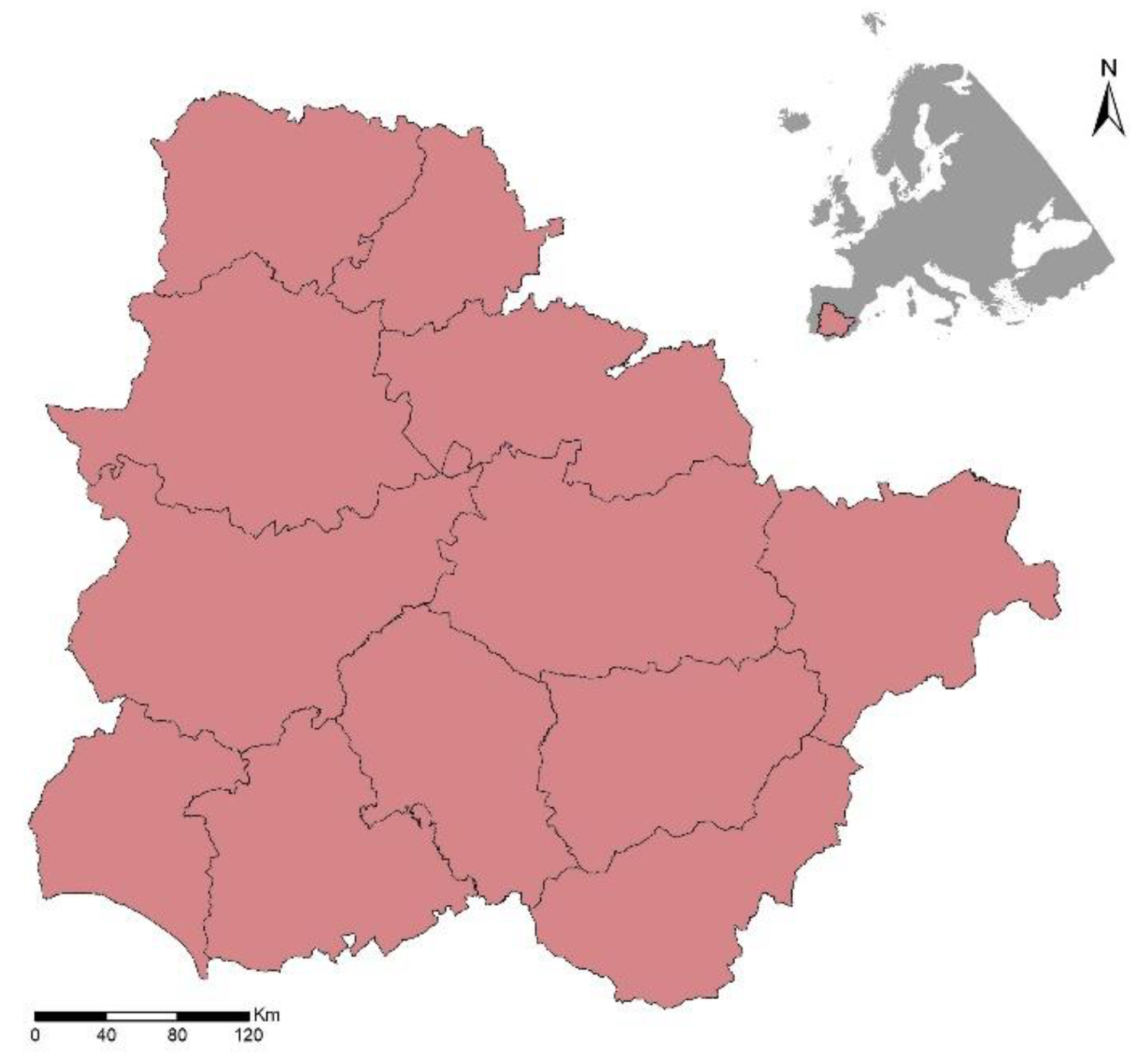
2.1. Study Area
2.2. Study Species
2.3. Habitat Suitability Modelling
2.3.1. Species Data
2.3.2. Environmental Data
2.3.3. Modelling Process with MaxEnt
2.4. The Design of Ecological Corridors
2.4.1. Selection of Core Areas
2.4.2. Resistance Raster
2.4.3. Corridor Design
2.5. Corridor Evaluation
Prioritization of Landscape Elements for Conservation and Restoration
3. Results
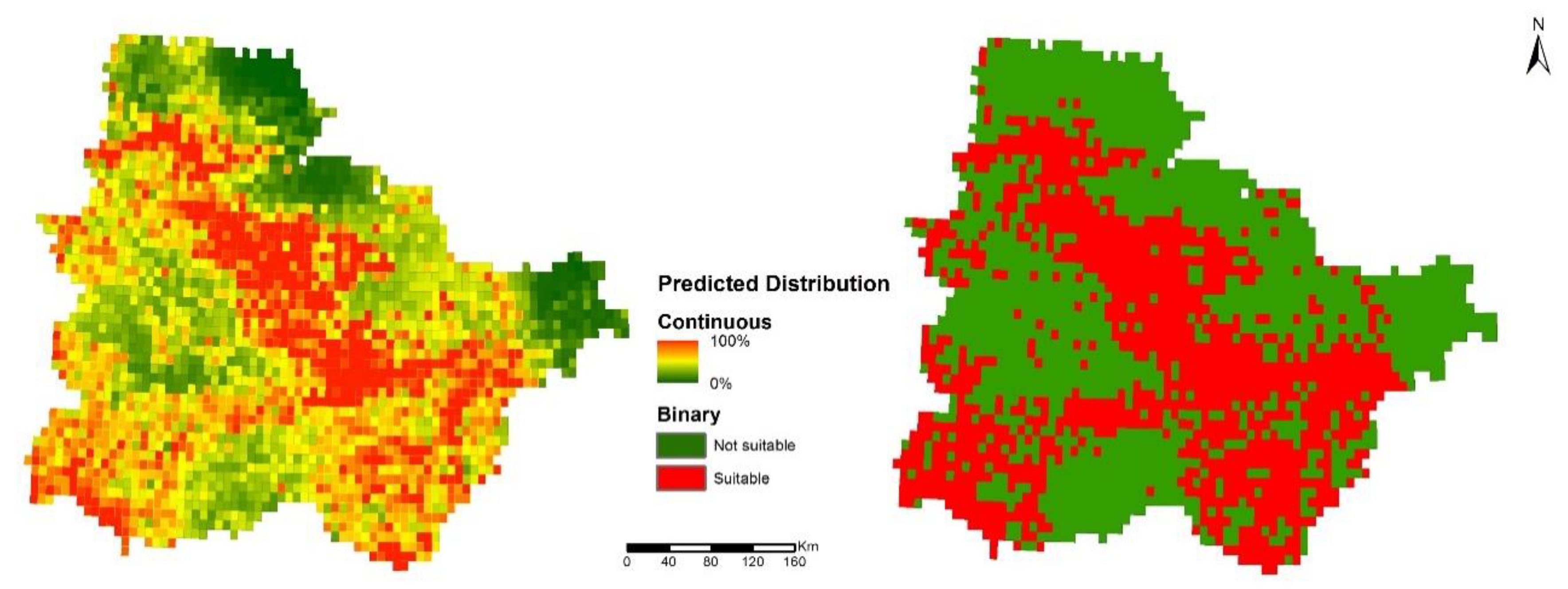
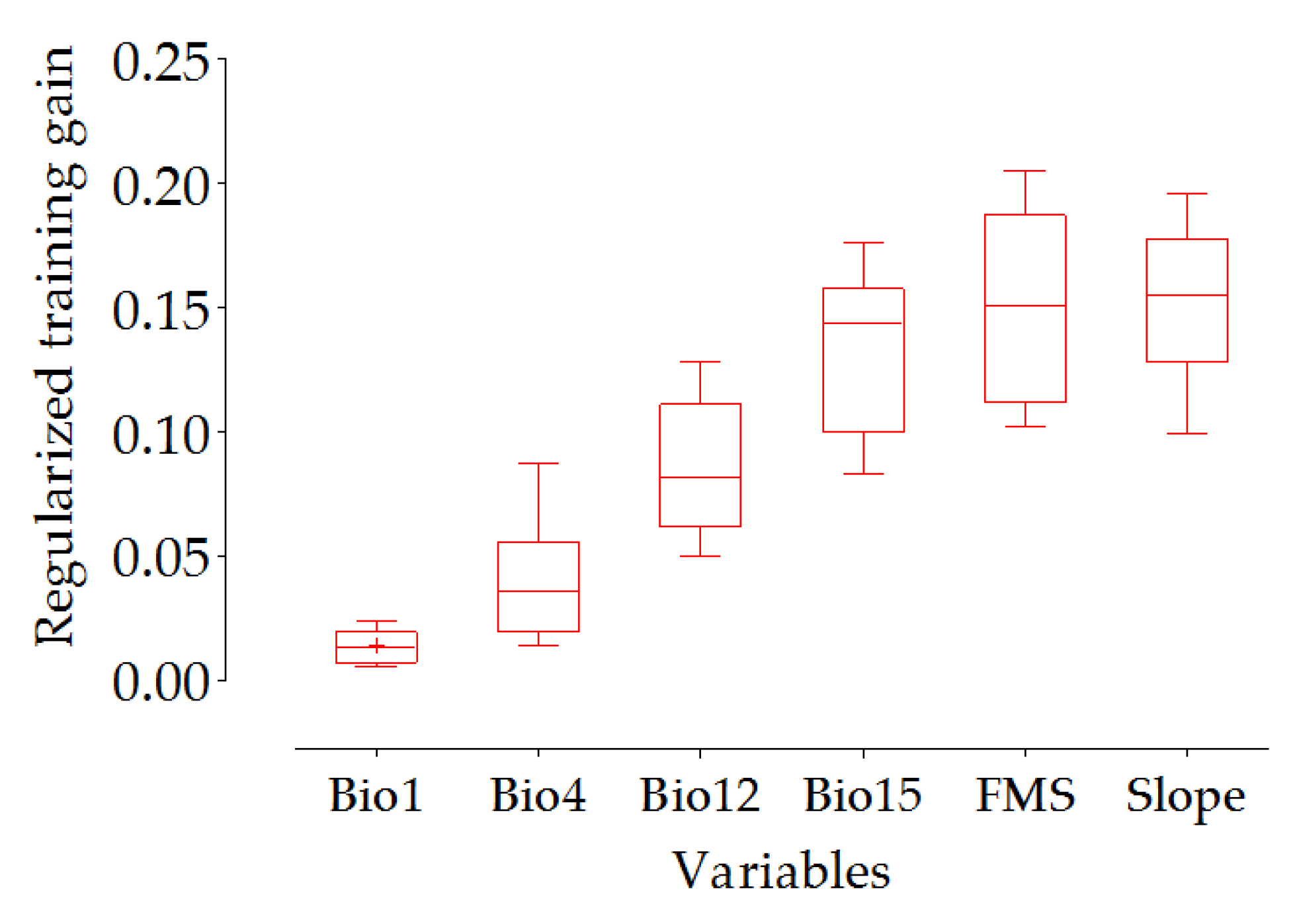
3.1. Evaluation of the Suitability Model
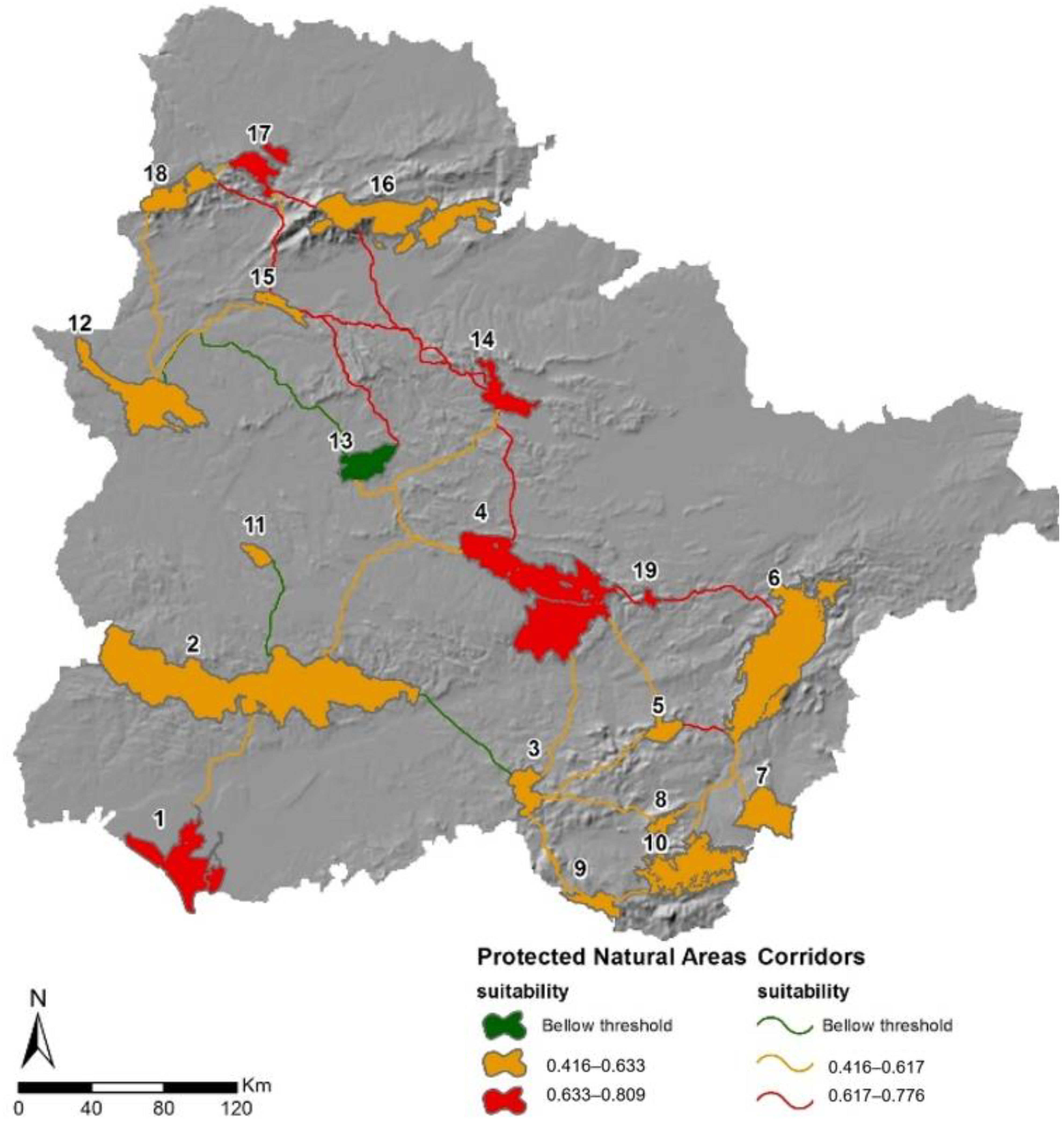
3.2. Suitability of Natural Protected Areas
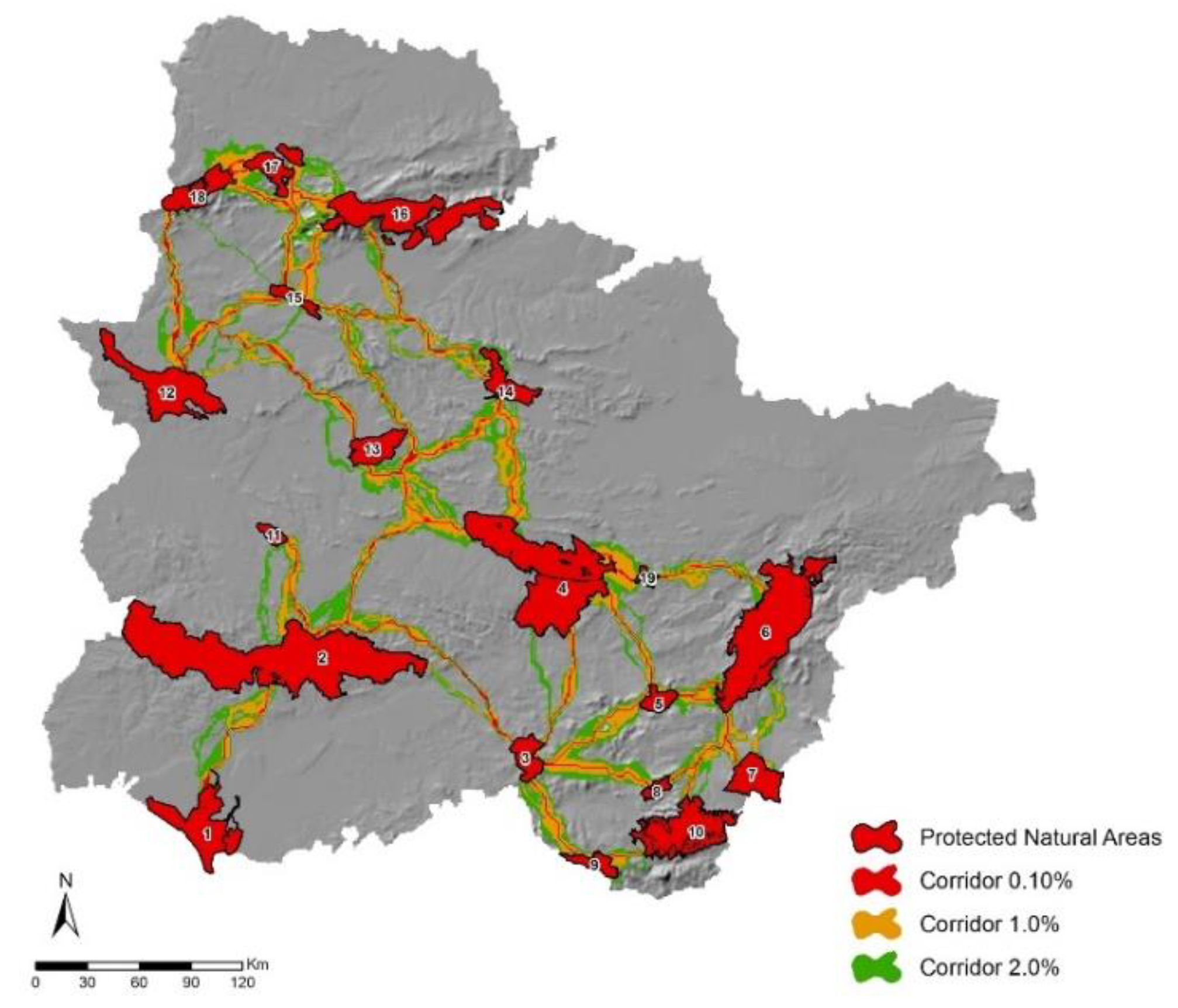
3.3. Corridor Design
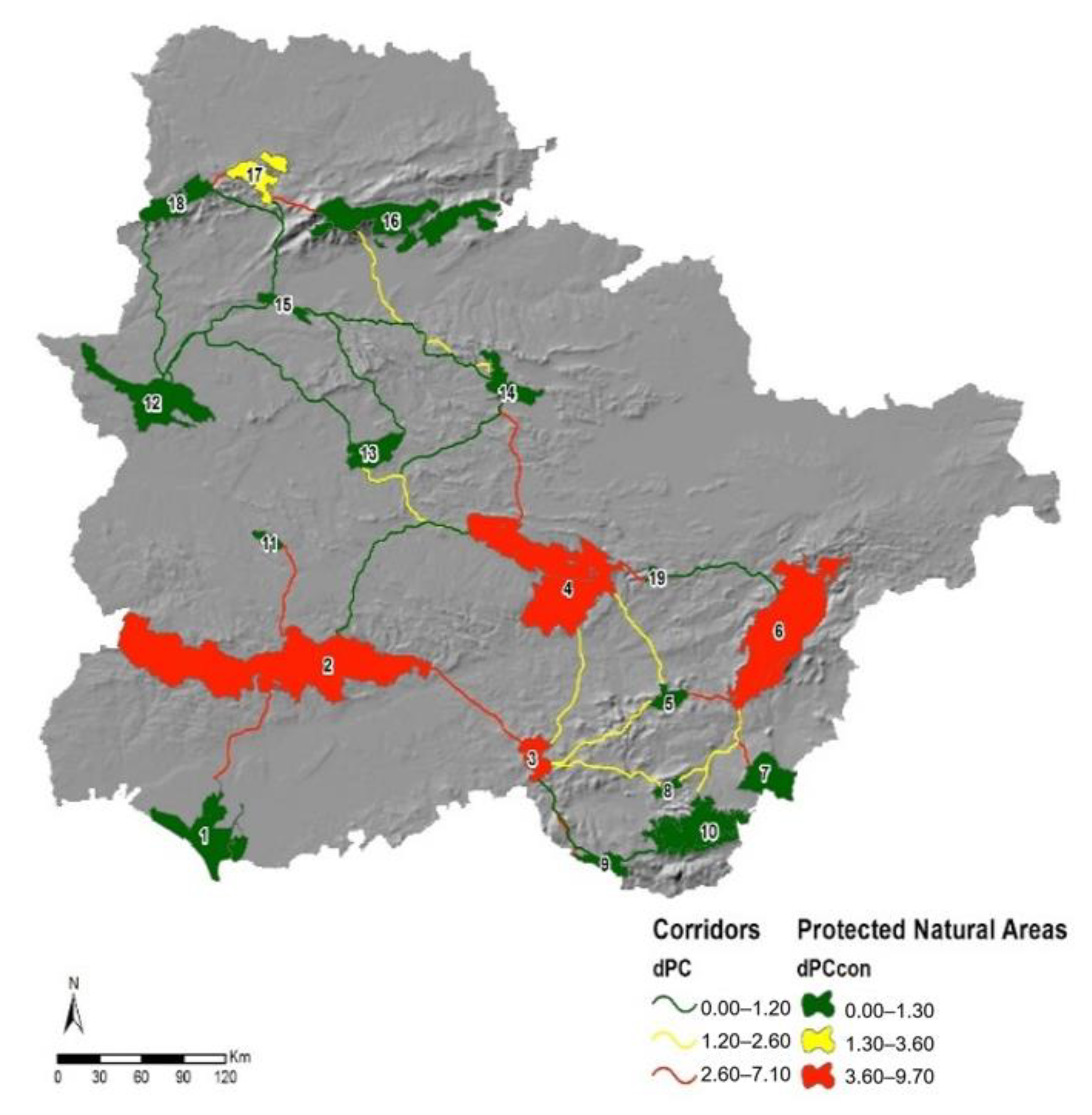
3.4. Corridors Evaluation
4. Discussion
4.1. Connectivity Studies in Environmental Assessments. The Practical Approach
4.2. Quality of Natural Protected Areas
4.3. Connectivity between NPAs. The Important Role of Sierra Morena
4.4. The Importance of Corridor Width
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zalasiewicz, J.; Williams, M.; Smith, A.; Barry, T.L.; Coe, A.L.; Bown, P.R.; Brenchley, P.; Cantrill, D.; Gale, A.; Gibbard, P.; et al. Are we now living in the Anthropocene? Gsa Today 2008, 18, 4. [Google Scholar] [CrossRef]
- Zalasiewicz, J.; Williams, M.; Steffen, W.; Crutzen, P. The new world of the Anthropocene. Environ. Sci. Technol. 2010, 44, 2228–2231. [Google Scholar] [CrossRef]
- Lovejoy, T.E.; Bierregaard, R.O., Jr.; Rylands, A.B.; Malcolm, J.R.; Quintela, C.E.; Harper, L.H.; Brown, K.S., Jr.; Powell, A.H.; Powell, G.V.N.; Schubart, H.O.R. Edge and Other Effects of Isolation on Amazon Forest Fragments; Sinauer Associates: Sunderland, MA, USA, 1986. [Google Scholar]
- Crooks, K.R.; Sanjayan, M. Connectivity Conservation: Maintaining Connections for Nature; Conservation Biology; Cambridge University Press: Cambridge, UK, 2006; Volume 14. [Google Scholar]
- Beier, P.; Penrod, K.L.; Luke, C.; Spencer, W.D.; Cabañero, C. South Coast Missing Linkages: Restoring connectivity to wildlands in the largest metropolitan area in the USA. In Connectivity Conservation: Maintaining Connections for Nature; Crooks, K.R., Sanjayan, M., Eds.; Conservation Biology; Cambridge University Press: Cambridge, UK, 2006; pp. 555–586. ISBN 9780521673815. [Google Scholar]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.A. Metapopulation dynamics and landscape ecology. In Metapopulation Biology; Academic Press: New York, NY, USA, 1997; pp. 43–62. [Google Scholar]
- Tischendorf, L.; Fahrig, L. On the usage and measurement of landscape connectivity. Oikos 2000, 90, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Moilanen, A.; Hanski, I. On the use of connectivity measures in spatial ecology. Oikos 2001, 95, 147–151. [Google Scholar] [CrossRef]
- Nelson, M.; Allen, J.; Ailing, A.; Dempster, W.F.; Silverstone, S. Earth applications of closed ecological systems: Relevance to the development of sustainability in our global biosphere. Adv. Space Res. 2003, 31, 1649–1655. [Google Scholar] [CrossRef]
- Beier, P.; Spencer, W.; Baldwin, R.F.; McRae, B.H. Toward best practices for developing regional connectivity maps. Conserv. Biol. 2011, 25, 879–892. [Google Scholar] [CrossRef]
- Schmitz, S. La géographie humaine et ses revues «internationales»: Globalisation ou fragmentation? Ann. Géo 2003, 632, 402–411. [Google Scholar] [CrossRef]
- Múgica, M.; De Lucio, J.V.; Martínez, C.; Sastre, P.; Atauri-Mezquida, J.A.; Montes, C. Integración Territorial de Espacios Naturales Protegidos y Conectividad Ecológica en Paisajes Mediterráneos; RENPA, Junta de Andalucía: Sevilla, Spain, 2002. [Google Scholar]
- Gurrutxaga, M.; Lozano, P.J.; del Barrio, G. GIS-based approach for incorporating the connectivity of ecological networks into regional planning. J. Nat. Conserv. 2010, 18, 318–326. [Google Scholar] [CrossRef]
- Clergeau, P.; Burel, F. The role of spatio-temporal patch connectivity at the landscape level: An example in a bird distribution. Landsc. Urban Plan. 1997, 38, 37–43. [Google Scholar] [CrossRef]
- Flather, C.H.; Bevers, M. Patchy reaction-diffusion and population abundance: The relative importance of habitat amount and arrangement. Am. Nat. 2002, 159, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Delibes, M. Current range and status of the Iberian lynx Felis pardina Temminck, 1824 in Spain. Biol. Conserv. 1992, 61, 189–196. [Google Scholar] [CrossRef]
- Rodríguez, A.; Delibes, M. Internal structure and patterns of contraction in the geographic range of the Iberian lynx. Ecography 2002, 25, 314–328. [Google Scholar] [CrossRef]
- Life + IBERLINCE. LIFE+ Iberlince project (LIFE10NAT/ES/570): Recuperación de la Distribución Histórica del Lince Ibérico (Lynx pardinus) en España y Portugal. Available online: www.iberlince.eu (accessed on 10 November 2019).
- Puerto, A.; Muñoz, J.C. Red de conectores ecológicos para el lince ibérico en la provincia de Huelva. In Tecnologías de la Información Geográfica: La Información Geográfica al servicio de los ciudadanos, Proceedings of the Congreso Nacional de Tecnologías de la Información Geográfica; Secretariado de Publicaciones de la Universidad de Sevilla: Seville, Spain, 2010; pp. 1028–1038. [Google Scholar]
- Blazquez-Cabrera, S.; Ciudad, C.; Gastón, A.; Simón, M.Á.; Saura, S. Identification of strategic corridors for restoring landscape connectivity: Application to the Iberian lynx. Anim. Conserv. 2019, 22, 210–219. [Google Scholar] [CrossRef]
- Illanas, S.; Gastón, A.; Blázquez-Cabrera, S.; Simón, M.A.; Saura, S. Selección del hábitat y permeabilidad del territorio para el lince ibérico (Lynx pardinus) en Andalucía: Influencia del estado de comportamiento y de la resolución cartográfica. Cuad. Soc. Española Cienc. For. 2017, 43, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Elith, J.; Burgman, M.A. Predictions and their validation: Rare plants in the Central Highlands, Victoria, Australia. In Predictions Species Occurences: Issues of Accuracy and Scale; Scott, J.M., Heglund, P.J., Morrison, M.L., Raphael, M.G., Wall, W.A., Samson, F.B., Eds.; Island Press: Covelo, CA, USA, 2002; pp. 303–314. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 14. [Google Scholar] [CrossRef]
- Schumaker, N.H. A Users Guide to the PATCH Model; U.S. Environmental Protection Agency, Environmental Research Laboratory: Corvallis, OR, USA, 1998.
- Urban, D.; Keitt, T. Landscape connectivity: A graph-theoretic perspective. Ecology 2001, 82, 1205–1218. [Google Scholar] [CrossRef]
- Adriaensen, F.; Chardon, J.P.; De Blust, G.; Swinnen, E.; Villalba, S.; Gulinck, H.; Matthysen, E. The application of ‘least-cost’modelling as a functional landscape model. Landsc. Urban Plan. 2003, 64, 233–247. [Google Scholar] [CrossRef]
- Minor, E.S.; Urban, D.L. Graph theory as a proxy for spatially explicit population models in conservation planning. Ecol. Appl. 2007, 17, 1771–1782. [Google Scholar] [CrossRef] [PubMed]
- Harary, F. Some historical and intuitive aspects of graph theory. SIAM Rev. 1960, 2, 123–131. [Google Scholar] [CrossRef]
- Saura, S.; Rubio, L. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography 2010, 33, 523–537. [Google Scholar] [CrossRef]
- Bunn, A.; Urban, D.; Keitt, T. Landscape connectivity: A conservation application of graph theory. J. Environ. Manag. 2000, 59, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, E.J. Quantifying Landscape Spatial Pattern: What Is the State of the Art? Ecosystems 1998, 1, 143–156. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Rodríguez, A. Lince ibérico—Lynx pardinus (Temminck, 1827). In Enciclopedia Virtual de los Vertebrados Españoles; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2004. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 10 November 2020).
- Rodríguez, A.; Calzada, J. Lynx pardinus (Iberian Lynx), 2015. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/species/12520/174111773 (accessed on 10 November 2020).
- Rodríguez, A.; Delibes, M. El Lince Ibérico (Lynx pardina) en España: Distribución y Problemas de Conservación; Instituto Nacional para la Conservación de la Naturaleza: Madrid, Spain, 1990. [Google Scholar]
- Palomares, F.; Delibes, M.; Ferreras, P.; Fedriani, J.M.; Calzada, J.; Revilla, E. Iberian lynx in a fragmented landscape: Predispersal, dispersal, and postdispersal habitats. Conserv. Biol. 2000, 14, 809–818. [Google Scholar] [CrossRef]
- Ferreras, P. Landscape structure and asymmetrical inter-patch connectivity in a metapopulation of the endangered Iberian lynx. Biol. Conserv. 2001, 100, 125–136. [Google Scholar] [CrossRef]
- Palomares, F. Vegetation structure and prey abundance requirements of the Iberian lynx: Implications for the design of reserves and corridors. J. Appl. Ecol. 2001, 38, 9–18. [Google Scholar] [CrossRef]
- Delibes, M.; Rodriguez, A.; Ferreras, P. Action Plan for the Conservation of the Iberian Lynx (Lynx pardinus) in Europe; Council of Europe: Strasbourg, France, 2000. [Google Scholar]
- Ferreras, P.; Beltrán, J.F.; Aldama, J.J.; Delibes, M. Spatial organization and land tenure system of the endangered Iberian lynx (Lynx pardinus). J. Zool. 1997, 243, 163–189. [Google Scholar] [CrossRef]
- GBIF: The Global Biodiversity Information Facility. Available online: https://www.gbif.org/es/ (accessed on 10 November 2019).
- ESRI. ArcMap 10.5; ESRI: Redlands, CA, USA, 2016. [Google Scholar]
- IGN Modelo Digital del Terreno con Paso de Malla de 200 m (MDT200). Datum ETRS89, Huso 29 y 30. Available online: http://centrodedescargas.cnig.es/CentroDescargas/index.jsp (accessed on 25 August 2018).
- MAPAMA. Mapa Forestal de España 1:50,000 (MFE). Available online: https://www.mapama.gob.es/es/biodiversidad/servicios/banco-datos-naturaleza/informacion-disponible/mfe50.aspx (accessed on 16 July 2018).
- Fick, S.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Schloerke, B.; Crowley, J.; Cook, D.; Briatte, F.; Marbach, M.; Thoen, E.; Elberg, A.; Larmarange, J. GGally: Extension to ‘ggplot2′’. R Package, Version 1.4.0. Available online: https://CRAN.R-project.org/package=GGally’ (accessed on 10 December 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780521700023. [Google Scholar]
- Hernández-Lambraño, R.; de la Cruz, D.R.; Sánchez-Agudo, J. Spatial oak decline models to inform conservation planning in the Central-Western Iberian Peninsula. For. Ecol. Manag. 2019, 441, 115–126. [Google Scholar] [CrossRef]
- Crespo-Luengo, G.; Hernández-Lambraño, R.E.; Barbero-Bermejo, I.; Sánchez-Agudo, J.Á. Analysis of Spatio-Temporal Patterns of Red Kite Milvus milvus Electrocution. Ardeola 2020, 67, 247–268. [Google Scholar] [CrossRef]
- Cantor, S.B.; Sun, C.C.; Tortolero-Luna, G.; Richards-Kortum, R.; Follen, M. A comparison of C/B ratios from studies using receiver operating characteristic curve analysis. J. Clin. Epidemiol. 1999, 52, 885–892. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Beier, P.; Majka, D.R.; Spencer, W.D. Forks in the road: Choices in procedures for designing wildland linkages. Conserv. Biol. 2008, 22, 836–851. [Google Scholar] [CrossRef]
- Majka, D.; Jenness, J.; Beier, P. ArcGIS Tools for Designing and Evaluating Corridors, 2007. CorridorDesign. Available online: http://corridordesign.org (accessed on 21 December 2020).
- WCS; CIESIN. Last of the Wild Project, Version 2, (LWP-2): Global Human Footprint Dataset (Geographic), 2005. Palisades. Available online: http://dx.doi.org/10.7927/H4M61H5F (accessed on 10 November 2019).
- Beier, P.; Majka, D.R.; Newell, S.L. Uncertainty analysis of least-cost modeling for designing wildlife linkages. Ecol. Appl. 2009, 19, 2067–2077. [Google Scholar] [CrossRef] [Green Version]
- Saura, S.; Torné, J. Conefor Sensinode 2.2: A software package for quantifying the importance of habitat patches for landscape connectivity. Environ. Model. Softw. 2009, 24, 135–139. [Google Scholar] [CrossRef]
- Jenks, G.F. The Data Model Concept in Statistical Mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Clauzel, C.; Bannwarth, C.; Foltete, J.-C. Integrating regional-scale connectivity in habitat restoration: An application for amphibian conservation in eastern France. J. Nat. Conserv. 2015, 23, 98–107. [Google Scholar] [CrossRef] [Green Version]
- de la Fuente, B.; Beck, P.S.A. Invasive Species May Disrupt Protected Area Networks: Insights from the Pine Wood Nematode Spread in Portugal. Forests 2018, 9, 282. [Google Scholar] [CrossRef] [Green Version]
- Duflot, R.; Avon, C.; Roche, P.; Bergès, L. Combining habitat suitability models and spatial graphs for more effective landscape conservation planning: An applied methodological framework and a species case study. J. Nat. Conserv. 2018, 46, 38–47. [Google Scholar] [CrossRef]
- Bodin, Ö.; Saura, S. Ranking individual habitat patches as connectivity providers: Integrating network analysis and patch removal experiments. Ecol. Model. 2010, 221, 2393–2405. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Gragera, F. Nuevos datos sobre la distribución pasada y actual del Lince ibérico (Felis pardina T.) en la provincia de Badajoz. Aegypius 1993, 11, 77–79. [Google Scholar]
- Oreja, J.A.G.; Vázquez, J.G.G. Situación del lince ibérico en Sierra de Gata. Doñana Acta Vertebr. 1996, 23, 91–98. [Google Scholar]
- Oreja, J.A.G. Non-natural mortality of the Iberian lynx in the fragmented population of Sierra de Gata (W Spain). Miscel Lània Zoològica 1998, 21, 31–35. [Google Scholar]
- Ordiz, A.; Llaneza, L. Situación del lince ibérico Lynx pardinus en la Sierra de Gata y aledaños. Galemys: Boletín informativo de la Sociedad Española para la conservacion y estudio de los mamiferos 2004, 16, 15–23. [Google Scholar]
- Sastre, P.; de Lucio, J.V.; Martínez, C. Modelos de conectividad del paisaje a distintas escalas. Ejemplos de aplicación en la Comunidad de Madrid. Ecosistemas 2002, 11, 1–10. [Google Scholar]
- Colino-Rabanal, V.J.; Lizana, M.; Peris, S.J. Factors influencing wolf Canis lupus roadkills in Northwest Spain. Eur. J. Wildl. Res. 2011, 57, 399–409. [Google Scholar] [CrossRef]
- Malo, J.E.; Suárez, F.; Díez, A. Can we mitigate animal-vehicle accidents using predictive models? J. Appl. Ecol. 2004, 41, 701–710. [Google Scholar] [CrossRef]
- Pascual-Hortal, L.; Saura, S. Comparison and development of new graph-based landscape connectivity indices: Towards the priorization of habitat patches and corridors for conservation. Landsc. Ecol. 2006, 21, 959–967. [Google Scholar] [CrossRef]
- Chester, C.C. Yellowstone to Yukon: Transborder conservation across a vast international landscape. Environ. Sci. Policy 2015, 49, 75–84. [Google Scholar] [CrossRef]
- Geldmann, J.; Barnes, M.; Coad, L.; Craigie, I.D.; Hockings, M.; Burgess, N.D. Effectiveness of terrestrial protected areas in reducing habitat loss and population declines. Biol. Conserv. 2013, 161, 230–238. [Google Scholar] [CrossRef]
- Gray, C.L.; Hill, S.L.L.; Newbold, T.; Hudson, L.N.; Börger, L.; Contu, S.; Hoskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7, 12306. [Google Scholar] [CrossRef] [Green Version]
- Andreassen, H.P.; Hertzberg, K.; Ims, R.A. Space-use responses to habitat fragmentation and connectivity in the root vole Microtus oeconomus. Ecology 1998, 79, 1223–1235. [Google Scholar] [CrossRef]
- Whitcomb, S.D.; Servello, F.A.; O’Connell, A.F., Jr. Patch occupancy and dispersal of spruce grouse on the edge of its range in Maine. Can. J. Zool. 1996, 74, 1951–1955. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description | Source |
|---|---|---|
| Slope | Gradient of the land and inclination | As of DEM [48] |
| FMS | Forest Map of Spain (vegetation cover) | [49] |
| Bio1 | Average annual temperature | [50] |
| Bio4 | Seasonal temperature (standard deviation * 100) | [50] |
| Bio12 | Annual precipitation | [50] |
| Bio15 | Seasonal precipitation (variation coefficient) | [50] |
| Cod. | NameNPAs | Protection Level | Surface Area (km2) | Suitability | HII | CP |
|---|---|---|---|---|---|---|
| 1 | Doñana | NP/NatP | 1034.76 | 65.50 | 35.30 | Y |
| 2a | Sierra de Aracena | NatP | 1868.01 | 46.10 | 30.35 | N |
| 2b | Sierra Norte de Sevilla | NatP | 1774.83 | 43.40 | 30.24 | N |
| 2c | Sierra de Hornachuelos | NatP | 600.31 | 46.20 | 25.19 | N |
| 3 | Sierras Subbéticas | NatP | 320.56 | 46.80 | 40.43 | N |
| 4a | Sierra de Cardeña | NatP | 384.49 | 66.70 | 26.51 | Y |
| 4b | Sierra de Andújar | NatP | 747.75 | 81.60 | 28.70 | Y |
| 4c | Valle de Alcudia | NatP | 1488.20 | 72.10 | 21.79 | Y |
| 5 | Sierra Mágina | NatP | 199.61 | 58.00 | 28.63 | N |
| 6a | Sierra de Cazorla | NatP | 2100.66 | 63.40 | 33.07 | N |
| 6b | Sierra de Castril | NatP | 126.95 | 86.30 | 36.33 | N |
| 7 | Sierra de Baza | NatP | 535.98 | 55.40 | 31.66 | N |
| 8 | Sierra de Huétor | NatP | 121.29 | 53.60 | 38.57 | N |
| 9 | Sierras de Tejeda | NatP | 213.22 | 52.10 | 28.48 | N |
| 10 | Sierra Nevada | NP/NatP | 1195.71 | 44.50 | 33.80 | N |
| 11 | Sierra Grande de Hornachos | ZRI | 121.90 | 54.20 | 37.95 | N |
| 12 | Sierra de San Pedro | ZRI | 1151.77 | 45.40 | 29.98 | N |
| 13 | Embalse Orellana | ZRI | 425.95 | 31.40 | 37.71 | Y |
| 14 | Cabañeros | NP | 409.07 | 67.56 | 18.60 | Y |
| 15 | Monfragüe | NP | 180.07 | 59.30 | 22.76 | N |
| 16 | Sierra de Gredos | RegP | 863.82 | 50.50 | 27.11 | N |
| 17a | Batuecas-Sierra Francia | NatP | 315.45 | 75.70 | 28.73 | N |
| 17b | Quilamas | SCZ | 106.50 | 67.97 | 27.74 | N |
| 18 | El Rebollar | SCZ | 496.36 | 62.00 | 27.41 | N |
| 19 | Despeñaperros | NatP | 76.49 | 81.80 | 27.54 | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbero-Bermejo, I.; Crespo-Luengo, G.; Hernández-Lambraño, R.E.; Rodríguez de la Cruz, D.; Sánchez-Agudo, J.Á. Natural Protected Areas as Providers of Ecological Connectivity in the Landscape: The Case of the Iberian Lynx. Sustainability 2021, 13, 41. https://doi.org/10.3390/su13010041
Barbero-Bermejo I, Crespo-Luengo G, Hernández-Lambraño RE, Rodríguez de la Cruz D, Sánchez-Agudo JÁ. Natural Protected Areas as Providers of Ecological Connectivity in the Landscape: The Case of the Iberian Lynx. Sustainability. 2021; 13(1):41. https://doi.org/10.3390/su13010041
Chicago/Turabian StyleBarbero-Bermejo, Iván, Gabriela Crespo-Luengo, Ricardo Enrique Hernández-Lambraño, David Rodríguez de la Cruz, and José Ángel Sánchez-Agudo. 2021. "Natural Protected Areas as Providers of Ecological Connectivity in the Landscape: The Case of the Iberian Lynx" Sustainability 13, no. 1: 41. https://doi.org/10.3390/su13010041
APA StyleBarbero-Bermejo, I., Crespo-Luengo, G., Hernández-Lambraño, R. E., Rodríguez de la Cruz, D., & Sánchez-Agudo, J. Á. (2021). Natural Protected Areas as Providers of Ecological Connectivity in the Landscape: The Case of the Iberian Lynx. Sustainability, 13(1), 41. https://doi.org/10.3390/su13010041