1. Introduction
Among the extensive types of forest ecosystems worldwide, there are some that appear locally, with a restricted distribution in small areas mainly due to evolutionary processes. These ecosystems are considered to be inherently vulnerable because of their usually narrow environmental envelopes, their geographically restricted distribution, and the fact that many of them appear to be near climatic thresholds [
1,
2]. As regards global warming, the potential reduction in available moisture and extreme drought conditions could be the biggest future threat, either alone or combined with wildfires. Thus, special attention should be given to these issues by the scientific community in order to secure the existence and sustainability of these important ecosystems [
3].
Tree natural regeneration is an essential process in forest ecosystems to ensure the persistence and resilience of forest stands when subjected to various disturbances, especially fires, and should contribute to a gradual process of recovery of the structure, function, and composition of the pre-disturbance ecosystem [
4,
5]. Successful seed germination and seedling survival and growth are the major factors for in situ species conservation [
6]. However, when naturally regenerating forests are unlikely to recover or face difficulties in their self-renewing process, an active restoration approach with enrichment plantings may be required to increase the abundance of poorly dispersed species or to protect endangered species and secure their long-term conservation [
4,
7,
8,
9].
Abies cephalonica Loudon (commonly known as Greek fir) is an endemic, important fir species with extremely restricted distribution found only in the southern part of the Balkan peninsula, and specifically in Southern and Central Greece [
10,
11,
12,
13]. It is a Mediterranean mountainous tree species that grows in relatively high altitudes, usually between 800 and 2000 m a.s.l. [
11]. Greek fir is shade-tolerant, with relatively high demand for soil moisture, an environmental variable considered as the crucial limiting factor for the species’ growth and survival [
14]. Due to its abovementioned ecophysiological attitudes, its regeneration, as is the case with many other fir species, in forest management systems is obtained inside an interior forest environment, under the protection of canopy cover of mother trees, applying shelterwood cuttings [
15,
16,
17]. This partially shaded environment protects the sensitive young fir seedlings from intensive sunlight and additionally creates favorable microsoil conditions, such as soil moisture, for the recruitment and growth of young seedlings [
18].
Due to climate change and summer drought episodes that have increased in the relatively more humid and colder regions of Greece [
19], a shift of wildfires toward higher altitudinal ecosystems, including fir ecosystems, has been observed [
20,
21,
22]. This phenomenon increases fire danger and, hence, the displacement risk of these mountainous fir ecosystems since they have not developed fire-adapted mechanisms. Greek fir does not produce serotinous cones and does not maintain a seed bank to secure its conservation after a wildfire [
21]. Findings from previous studies have concluded that the post-fire regeneration of
A. cephalonica in Mt. Parnitha, three years after the wildfire, tends to be zero despite the existence of nearby unburned fir stands [
6]. However, Raftoyannis and Spanos [
23], who studied the post-fire regeneration of the Greek fir in a much more humid environment, found that it depended mainly on the distance from the seed source, i.e., from the remnant stands or the border of the unburned zone. Thus, in order to ensure the conservation and endurance of Greek fir forests following various disturbances, effective restoration techniques should be developed, following the general rules of reforestation and aiming at the optimization of carbon sequestration, biodiversity conservation, and livelihood benefits [
24]. These techniques concern the selection of suitable site-specific provenances, setting specific standards for seedling production, and appropriate site-preparation techniques, especially those addressing post-fire environmental conditions. Special care should also be given to the fact that Greek fir ecosystems are mostly distributed within protected areas (e.g., European ecological network Natura 2000). Thus, the provenance (origin) of the plant reproductive material intended for reforestation projects in protected areas is of high importance, in order to avoid any genetic degradation of the local fir populations. Taking into consideration the fact that wildfires were not a common phenomenon in fir ecosystems in the past, many of the above issues concerning post-fire restoration have not been studied at all. Scarce studies have examined the issues pertaining to Greek fir, such as the influence of seedling height and site parameters (shade, competitive vegetation, bedrock, weather conditions) on survival and growth of outplanted
A. cephalonica seedlings [
25,
26]. However, most of the factors affecting reforestation success in a post-fire environment have not been thoroughly studied. Although the diversification of genetic plant material in reforestations is considered a promising strategy to promote forest adaptation to climate change [
27], no research so far has studied the survival and growth of different provenances of
A. cephalonica in a Mediterranean post-fire environment.
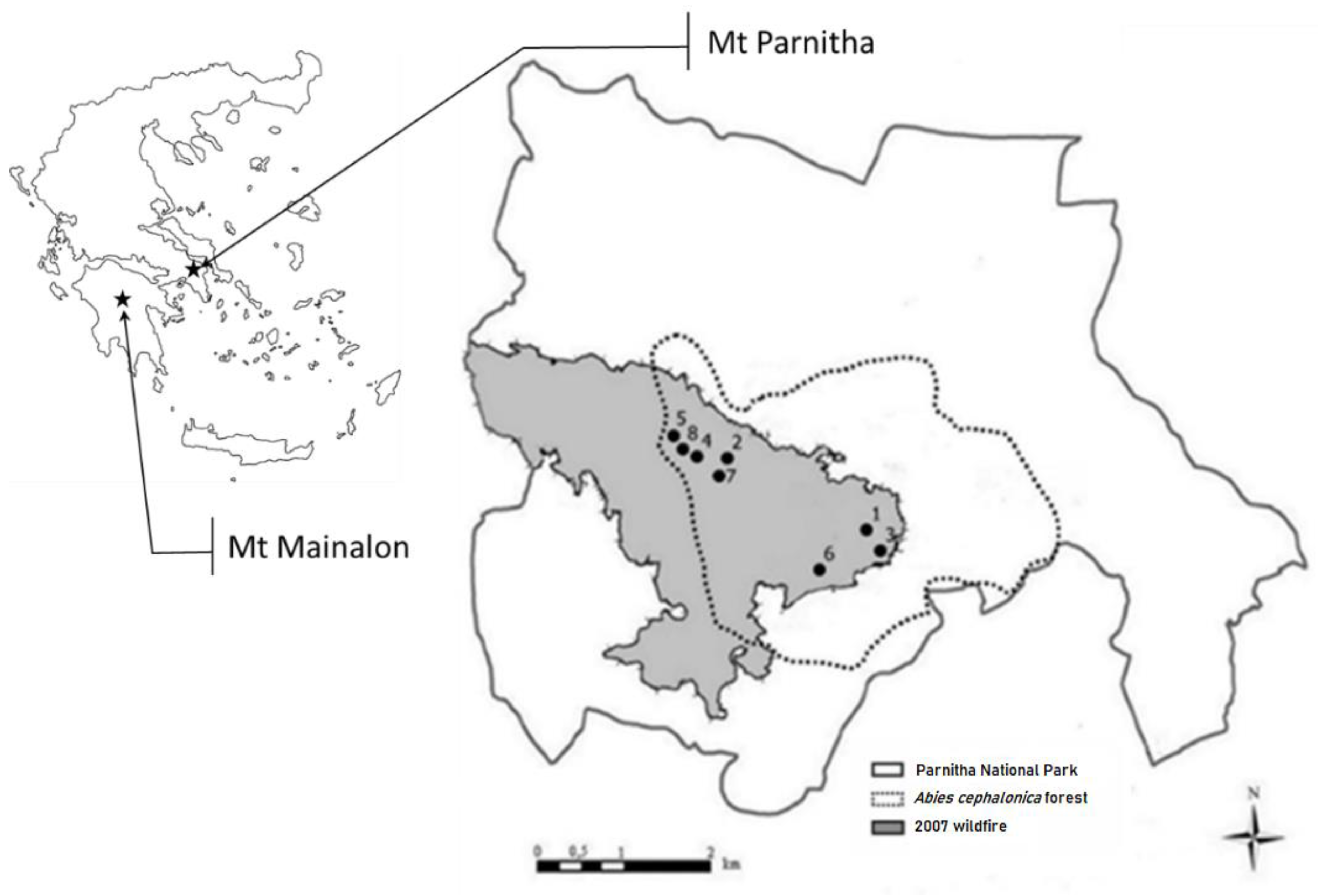
In the present study, we document, for the first time, the effect of seedling provenance as well as site heterogeneity (altitude, aspect, slope, bedrock–soil parent material) on a 3-year monitoring reforestation success (survival and growth) of Greek fir seedlings in the post-fire environment of Parnitha National Park (Attica, central Greece). Furthermore, seedling damage on crown architecture and apical bud, probably attributed to wildlife activity (local population of red deer, Cervus elaphus-Cervidae) in the post-fire environment was assessed.
4. Discussion
Our results suggest that the performance of Greek fir plantings in a post-fire environment, in the area of Parnitha National Park, were quite successful and presented an overall relatively high survival rate seven years after the first planting period (2008) and eight years after the fire event. Similarly, the seedlings presented satisfactory height and crown diameter growth compared to the baseline values (autumn 2012). A rather high percentage of seedlings having damaged branches and crowns was found, while the damage on seedling apical buds was much lower at the end of the study period. The above results interestingly indicate that although the seedlings were planted in a post-fire forest environment, without being protected by a forest canopy cover, their performance was quite satisfactory. This seems to be contradictory to the species’ ecophysiological attitudes, since it is characterized as shade-tolerant and semi-drought-tolerant [
39], and its regeneration in forest management systems is performed within an interior forest environment, under the mother stand canopy cover [
15,
16,
17]. The rather high field performance of the Greek fir seedlings could be attributed to the fact that these were watered during the post-planting period, especially in the first 3–4 years following reforestations and during the summer dry period. Therefore, a correlation analysis between growth or survival with climatic data was not attempted. Watering seems to play a crucial role in seedling survival and growth, by covering them with the necessary moisture for well-balanced water status and temperature fluctuation during the hot and dry summer months. Taking into consideration the fact that
A. cephalonica can grow in a sub-humid climate characterized by a relatively low annual precipitation of between 700 and 800 mm [
40], a watering regime during the crucial post-planting summer months should be considered necessary in forest practice for effective seedling survival and growth.
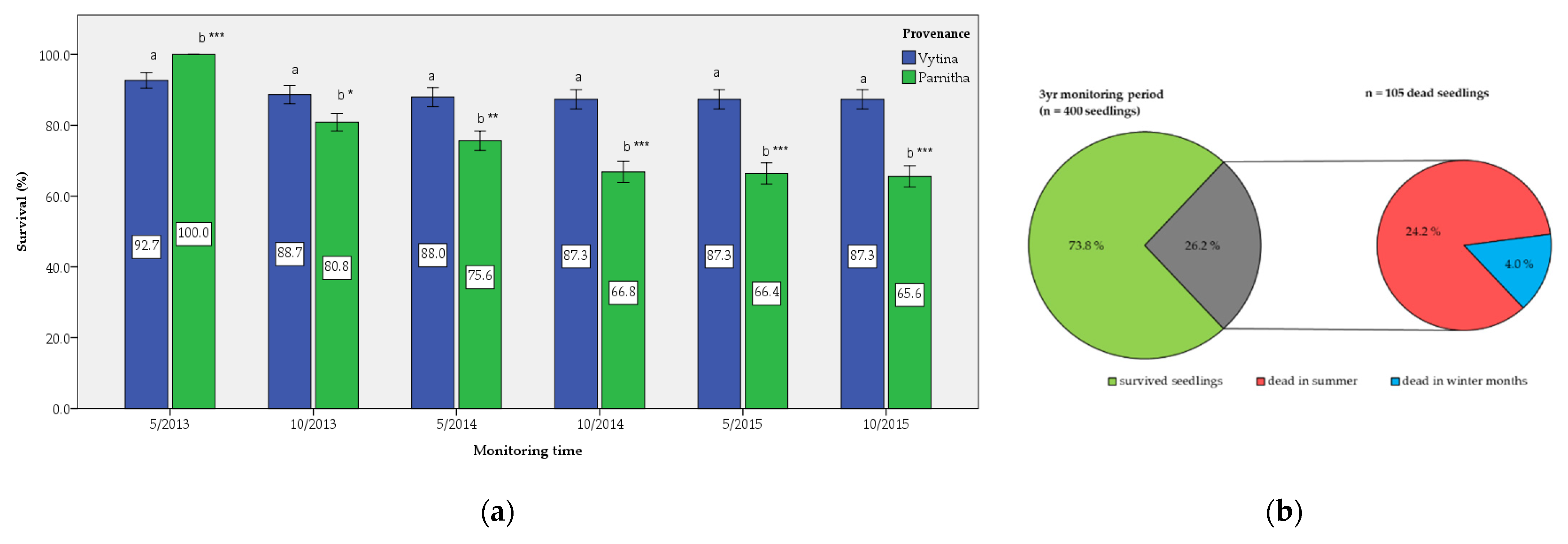
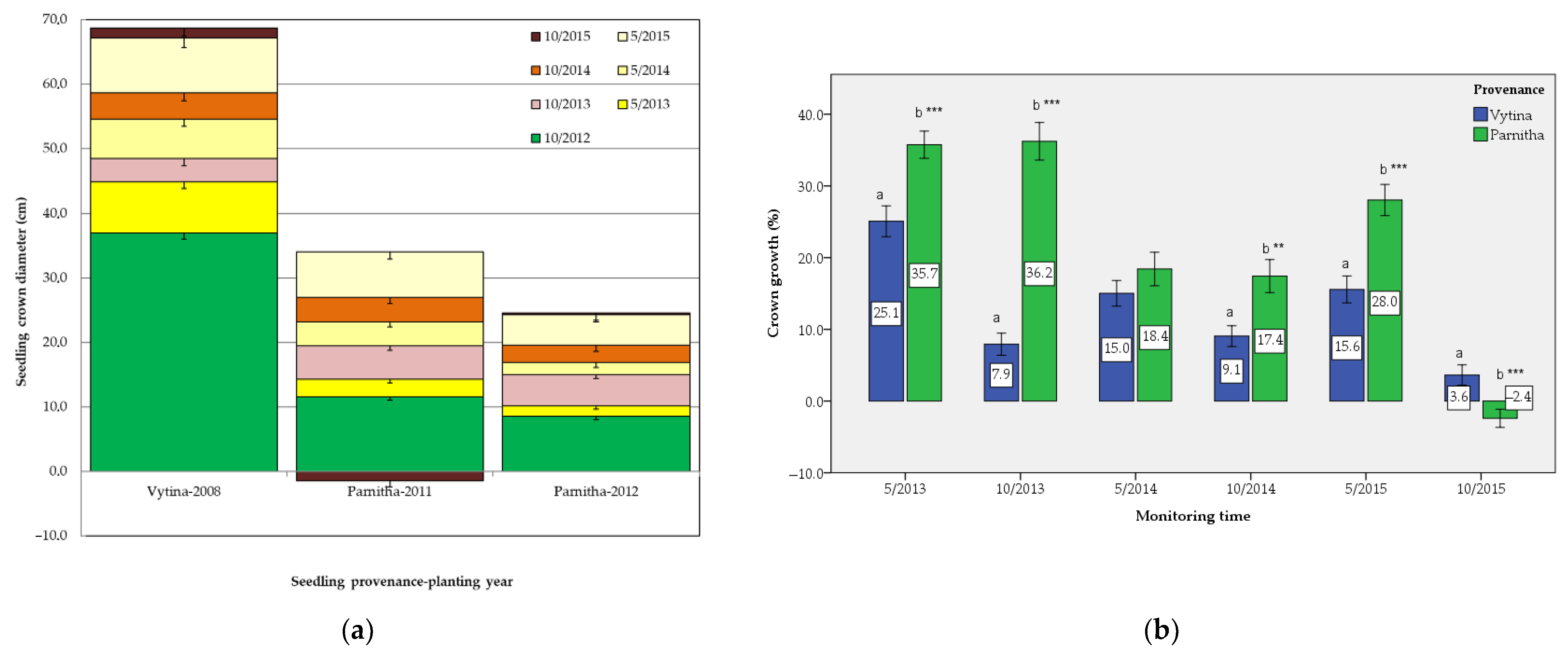
Overall, our results indicate that
A. cephalonica seedlings from the two different provenances (origins) respond quite similarly after reforestations. Higher survival rates were achieved by the seedlings originating from Vytina, while seedlings from Parnitha exhibited higher growth. This could be attributed to the fact that the summer drought periods affected the mortality of Parnitha seedlings more, while Vytina seedlings were slightly affected, since the latest provenance was mainly planted in favored sites and aspects. In addition, we can assume that the lower survival rate of Parnitha seedlings is interpreted in the reforestation period and year, 1–2 years before the onset of monitoring in the present study. According to Ganatsas et al. [
6], Vytina seedlings seemed to overcome stress 2 years after planting and adapted to the local post-fire conditions and were well established (with a mean survival rate of 65.3%). However, these results preceded the examined monitoring period, thus explaining the high and almost stable survival rate observed in this study. A similar survival rate (61.5%) was recorded by other researchers for the same provenance 2 years after the reforestation [
26]. Lower survival percentages were recorded 3 and 4 years after planting, 52.7% and 46.2%, respectively [
26].
For the Mediterranean forest plantations, surviving summer water stress after planting during the early years presents a major challenge [
41]. Detsis et al. [
25] also reported that water availability is among the basic factors affecting Greek fir seedlings’ performance in the area. This response to the drought period could be linked either to genetic adaptations of individual Greek fir provenances or the planting stock quality; planting stock from Vytina may have been more hardened before and at the time of planting. In addition, wildfire characteristics exert a strong influence on stand characteristics [
42], and the variability throughout the landscape of both fire intensity and severity produces heterogeneous post-fire environments [
43], thus affecting plant establishment [
42,
44,
45]. However, any possible effect of wildfire severity and intensity on seedlings’ performance was not studied, since there were no available spatial data for the wildfire, and the wildfire completely destroyed the forest in the burnt area, meaning that no spatial differentiations were observed.
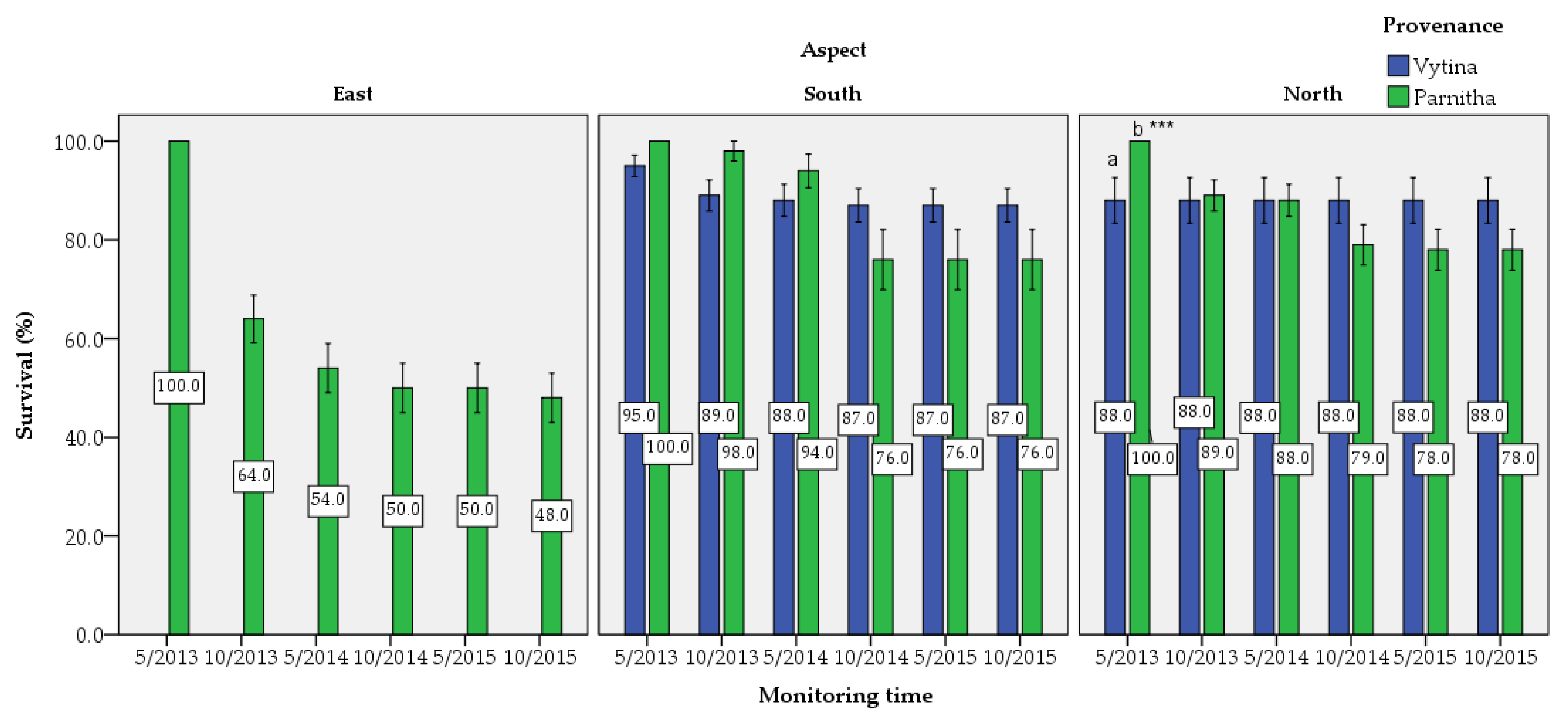
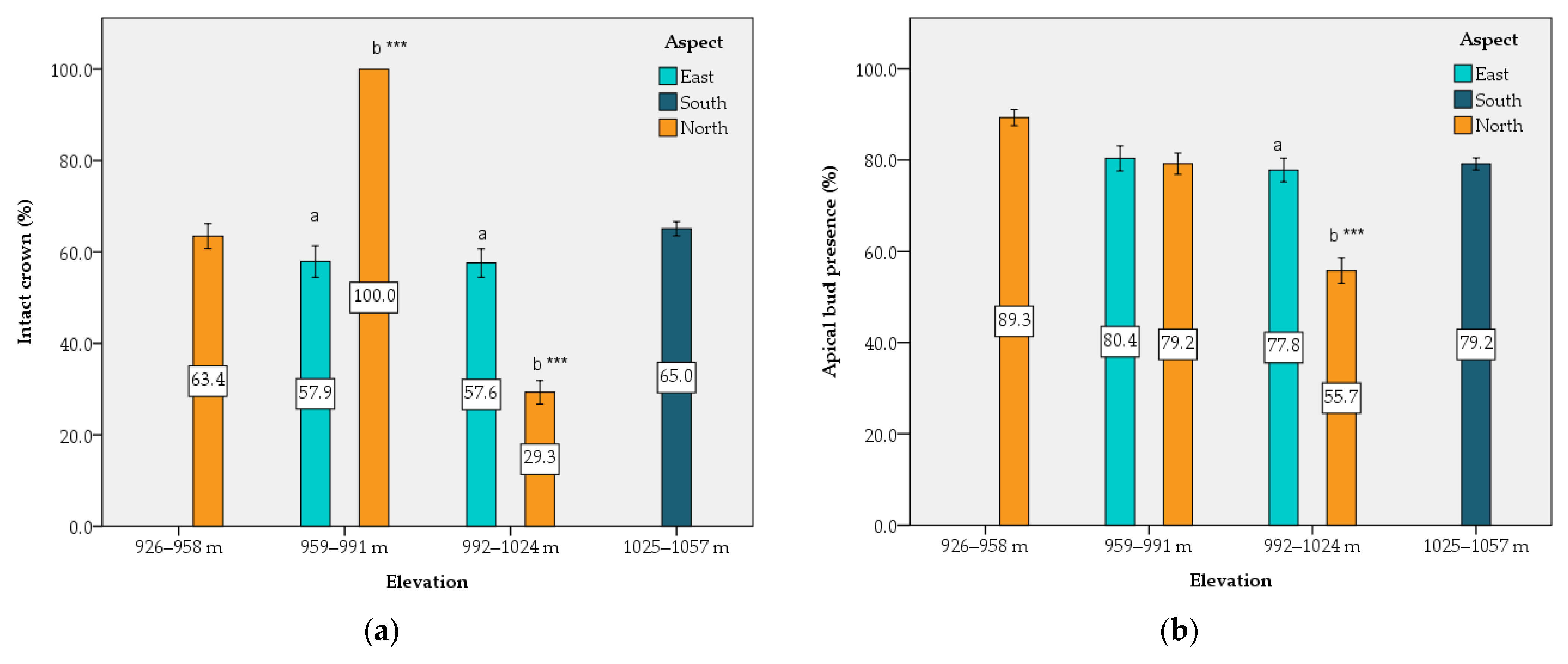
Furthermore, seedling survival was slightly higher in north-facing slopes followed by the south-facing ones, while it was lower in east-facing transects. This may be explained as planting was performed immediately after fire, and under the great pressure imposed by citizens for emergency reforestation. Planting started mainly in north-facing slopes, and thus, seedlings were well adapted to local microenvironments. Contrary to the common belief that northern aspects are much more favorable than southern ones, the transect aspect did not statistically influence seedling survival rate significantly, as had been reported by other studies [
46]. Moreover, elevation and aspect affect the variability of fire severity [
45], which in turn affects the survival of seedlings. In general, in divergent Mediterranean environments, aspect affects survival in varying ways, resulting in mixed conclusions [
47].
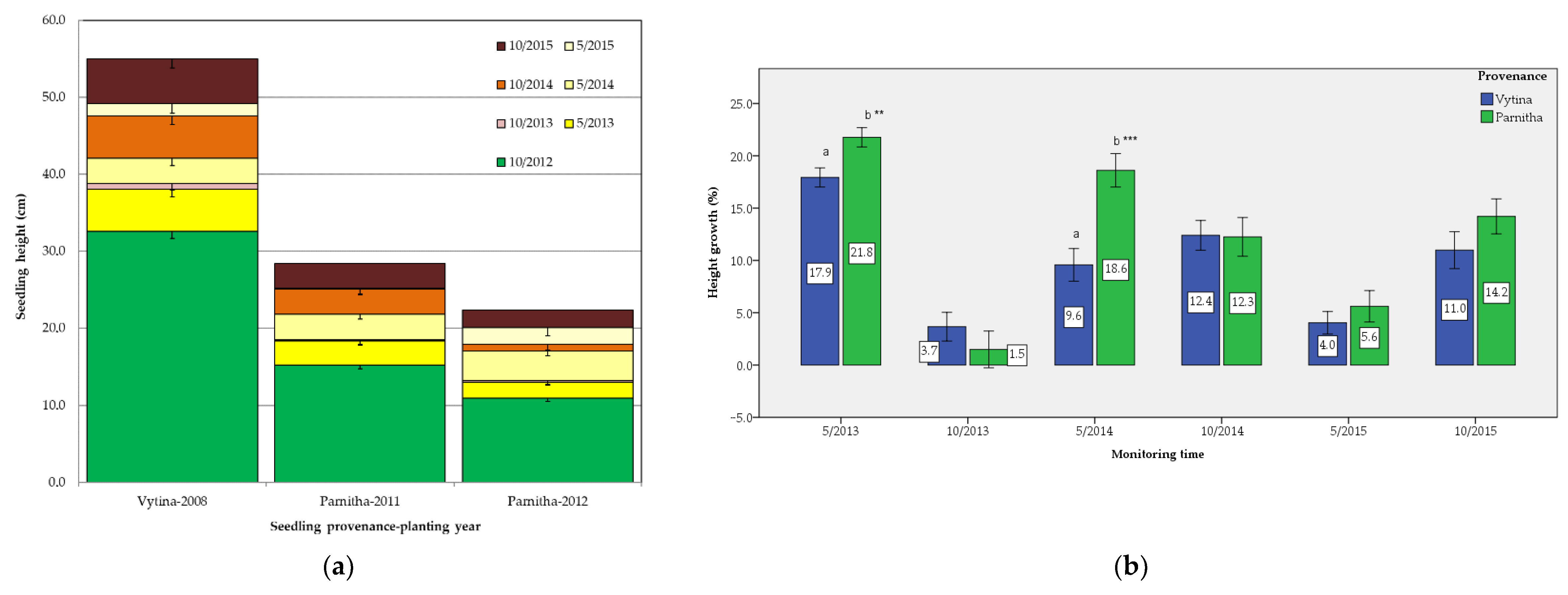
During the monitoring period, the height and crown diameter growth of Parnitha seedlings was significantly greater than that of Vytina’s and thus adapted better to the local site conditions. Species adaptation to local conditions is a well-known mechanism in plant ecology [
48,
49] which is highly taken into consideration during planting for ecosystem restoration [
50].
Site heterogeneity remarkably affected seedling performance [
51]. Among the studied environmental factors, the aspect and the altitude seemed to play an important role in seedling growth. At the end of the monitoring period, a greater growth of Greek fir seedlings, both in height and crown diameter, was observed in the north- and east-facing transects, respectively, and in medium altitudes (959–1024 m), probably due to their ability to create more favorable temperatures for plant growth [
52], especially under dry climatic conditions [
53]. This growth pattern is common in forest ecosystems in the northern hemisphere [
52,
54]; however, the interaction with other environmental variables should be considered. In general, seedling performance is affected by the microenvironments [
55]. By contrast, the slope and type of bedrock (soil parent material) did not affect seedling growth.
The percentage of intact crown and the apical bud presence in the seedlings were both highly affected by altitude and aspect. The greater percentage of seedlings with damaged crowns and the lower apical bud presence were recorded in north-facing transects and the medium altitudinal zone (992–1024 m). This can be explained as the local population of red deer thrives mainly in the north-facing slopes, since these sites are the most isolated from human presence. This isolation favors deer movement and activity.
Taking into consideration the negative impact of the current climate change, as predicted by several scenarios, it is expected that many forest species could be at high risk from its negative impact [
56]. Under these changing environmental conditions, endemic species, such as the studied Greek fir, may not be able to adjust to these changes and adapt and may therefore be the first to go extinct [
3]. For example, the inability of the species to regenerate naturally after fire may lead to a secondary ecological succession to other (degraded) plant communities [
6]. Native forests may change in composition, and some species may be entirely eliminated over large areas as a result of climatic change. Conservation of genetic resources will be necessary to restore declining forests; even in a seed source that is not adapted to the plantation site, some seedlings will survive and grow remarkably well [
57].
The early establishment and abundance of shade-tolerant conifer species, as was (partly) done in Parnitha National Park, contrasts with traditional stand development models which need significant time for monitoring, evaluation, resource availability administration, and knowledge and might have long-term impacts on biodiversity, landscapes, and livelihoods [
24]. Recruitment following large-scale disturbance is assumed to take decades, if not centuries [
58]. Due to the lack of an official and universal approach on forest restoration, imposing principles and standards can increase the effectiveness of restoration processes across different ecosystems, locally or globally [
59]. Long-term reforestation success in areas which are highly vulnerable to climate change depends on using plant material with appropriate levels of genetic diversity [
60], with local or regional genetic variation, which ensures the survival and resilience of a planted forest [
24]. Collecting seeds from native trees belonging to different local provenances across the parent population is an excellent practice [
61,
62,
63] that ensures adequate genetic diversity determined by the size of the parental population [
63] and collection rules [
64]. However, it may be prudent to include some germplasm of the same species from a predicted “future climate”, namely, a region with a climate similar to that predicted for the area being restored [
59]. In addition, population size is probably one of the most important criteria, useful for indicating both genetic erosion and genetic pollution [
65] as well as being an essential element for future evolution [
66]. Trees opt to make rapid adaptive changes while maintaining a high level of genetic diversity within the population. Their genetic diversity along with the environmental conditions influence the result of combined evolutionary forces [
67].
Thus, the conservation of biodiversity within the National Park of Parnitha enforces the need for planting A. cephalonica seedlings. The findings of the study aspire to mitigate the risk of the important endemic Mediterranean fir A. cephalonica extinction. However, further research data are urgently needed to secure species conservation and sustainability from a long-term perspective.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}