
Pro197Thr Substitution in Ahas Gene Causing Resistance to Pyroxsulam Herbicide in Rigid Ryegrass (Lolium Rigidum Gaud.)

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
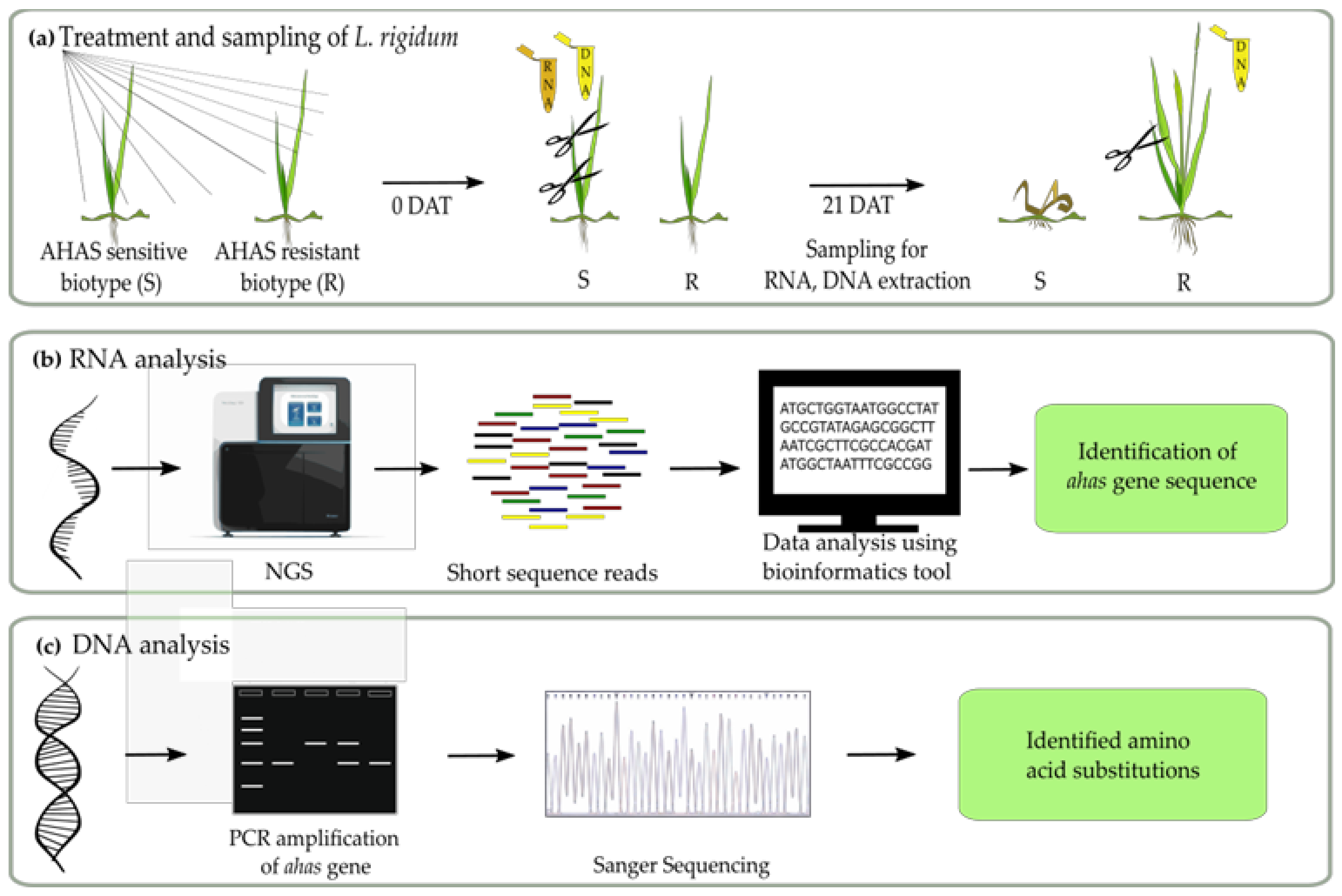
2.1. Plant Materials and Growth Conditions
2.2. De Novo Transcriptome Sequencing and Assembly
2.3. Identification of Ahas mRNA in L. Rigidum
2.4. Dose-Response Analysis
2.5. Ahas Gene of L. Rigidum Amplification and Sequencing
3. Results
3.1. Transcriptome Reconstruction and Identification of Ahas Cds
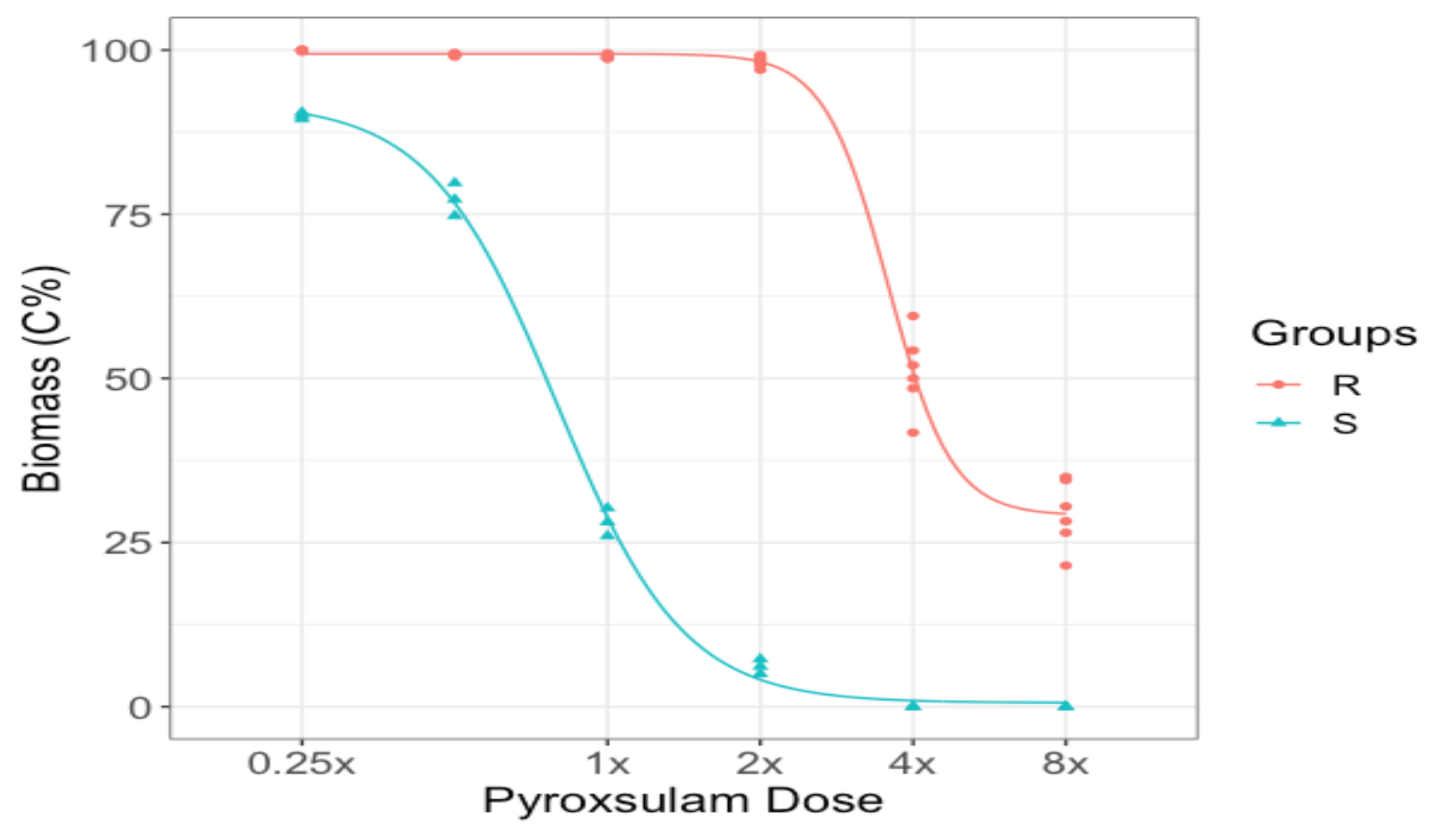
3.2. Dose–Response Experiment
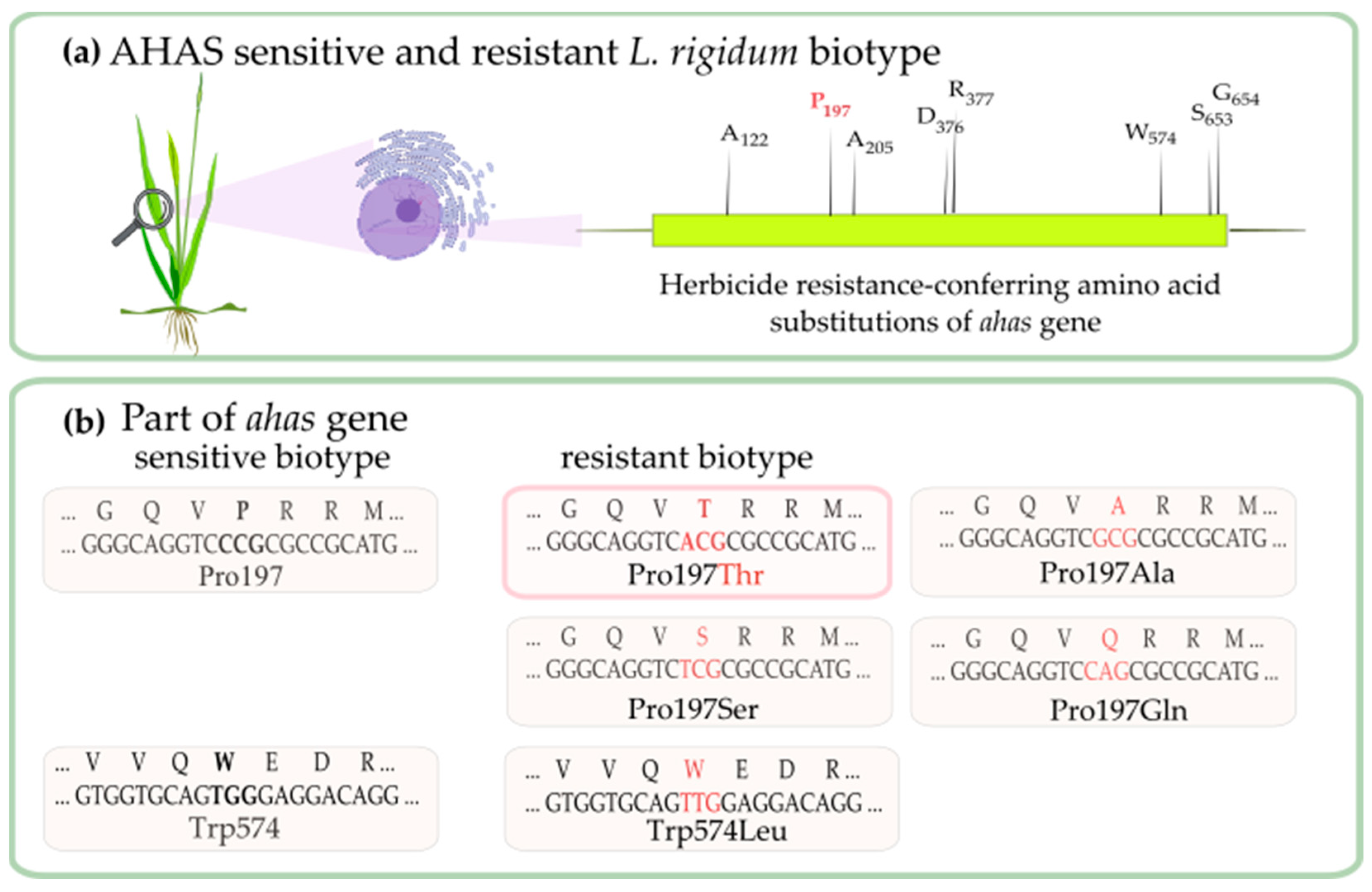
3.3. Determination of Mutation Points of Ahas Gene in L. Rigidum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowden, W.M. Chromosome numbers and taxonomic notes on northern grasses: iii. twenty-five genera. Can. J. Bot. 1960, 38, 541–557. [Google Scholar] [CrossRef]
- Castellanos-Frías, E.; De León, D.G.; Bastida, F.; Gonzalez-Andujar, J.L. Predicting global geographical distribution of Lolium rigidum (rigid ryegrass) under climate change. J. Agric. Sci. 2016, 154, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Preston, C.; Wakelin, A.M.; Dolman, F.C.; Bostamam, Y.; Boutsalis, P. A Decade of Glyphosate-Resistant Lolium around the World: Mechanisms, Genes, Fitness, and Agronomic Management. Weed Sci. 2009, 57, 435–441. [Google Scholar] [CrossRef]
- Niknam, S.; Moerkerk, M.; Cousens, R. Weed seed contamination in cereal and pulse crops. In Proceedings of the 13rd Australian Weeds Conference, Perth, Australia, 8–13 September 2002; Jacob, H.S., Dodd, J., Moore, J.H., Eds.; Plant Protection Society of Western Australia Inc.: Victoria, Australia, 2002; pp. 59–62. [Google Scholar]
- Preston, C.; Tardif, F.J.; Christopher, J.T.; Powles, S.B. Multiple Resistance to Dissimilar Herbicide Chemistries in a Biotype of Lolium rigidum Due to Enhanced Activity of Several Herbicide Degrading Enzymes. Pestic. Biochem. Physiol. 1996, 54, 123–134. [Google Scholar] [CrossRef]
- Beckie, H. Herbicide-Resistant Weeds: Management Tactics and Practices. Weed Technol. 2006, 20, 793–814. [Google Scholar] [CrossRef]
- Christopher, J.T.; Powles, S.B.; Liljegren, D.R.; Holtum, J.A. Cross-resistance to herbicides in annual ryegrass (Lolium rigidum): II. Chlorsulfuron resistance involves a wheat-like detoxification system. Plant Physiol. 1991, 95, 1036–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, M.J.; Walsh, M.J.; Llewellyn, R.S.; Powles, S.B. Widespread occurrence of multiple herbicide resistance in Western Australian annual ryegrass (Lolium rigidum) populations. Aust. J. Agric. Res. 2007, 58, 711–718. [Google Scholar] [CrossRef]
- Han, H.; Yu, Q.; Owen, M.J.; Cawthray, G.R.; Powles, S.B. Widespread occurrence of both metabolic and target-site herbicide resistance mechanisms in Lolium rigidum populations. Pest Manag. Sci. 2016, 72, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Powles, S.B.; Matthews, J.M. Multiple herbicide resistance in annual ryegrass (Lolium rigidum): A driving force for the adoption of integrated weed management. In Resistance’91: Achievements and Developments in Combating Pesticide Resistance; Springer: Berlin/Heidelberg, Germany, 1992; pp. 75–87. [Google Scholar]
- Burnet, M.W.M.; Hart, Q.; Holtum, J.A.M.; Powles, S.B. Resistance to Nine Herbicide Classes in a Population of Rigid Ryegrass (Lolium rigidum). Weed Sci. 1994, 42, 369–377. [Google Scholar] [CrossRef]
- Torra, J.; Montull, J.M.; Taberner, A.; Onkokesung, N.; Boonham, N.; Edwards, R. Target-Site and Non-target-Site Resistance Mechanisms Confer Multiple and Cross- Resistance to ALS and ACCase Inhibiting Herbicides in Lolium rigidum From Spain. Front. Plant Sci. 2021, 12, 88. [Google Scholar] [CrossRef] [PubMed]
- Owen, M.J.; Martinez, N.J.; Powles, S.B. Multiple herbicide-resistant Lolium rigidum (annual ryegrass) now dominates across the Western Australian grain belt. Weed Res. 2014, 54, 314–324. [Google Scholar] [CrossRef]
- Neve, P.; Powles, S.B. High survival frequencies at low herbicide use rates in populations of Lolium rigidum result in rapid evolution of herbicide resistance. Heredity 2005, 95, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the evolution of herbicide resistance in weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [Green Version]
- Harker, K.N.; O’Donovan, J.T. Recent Weed Control, Weed Management, and Integrated Weed Management. Weed Technol. 2013, 27, 1–11. [Google Scholar] [CrossRef]
- Kutasy, B.; Farkas, Z.; Kolics, B.; Decsi, K.; Hegedűs, G.; Kovács, J.; Taller, J.; Tóth, Z.; Kálmán, N.; Kazinczi, G.; et al. Detection of Target-Site Herbicide Resistance in the Common Ragweed: Nucleotide Polymorphism Genotyping by Targeted Amplicon Sequencing. Diversity 2021, 13, 118. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2021. Available online: www.weedscience.org (accessed on 10 June 2021).
- Gill, G. Development of herbicide resistance in annual ryegrass populations (Lolium rigidum Gaud.) in the cropping belt of Western Australia. Aust. J. Exp. Agric. 1995, 35, 67–72. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Powles, S.B. Mutations of the ALS gene endowing resistance to ALS-inhibiting herbicides in Lolium rigidum populations. Pest Manag. Sci. 2008, 64, 1229–1236. [Google Scholar] [CrossRef]
- Tranel, P.J.; Wright, T.R. Resistance of weeds to ALS-inhibiting herbicides: What have we learned? Weed Sci. 2002, 50, 700–712. [Google Scholar] [CrossRef]
- Délye, C.; Causse, R.; Michel, S. Genetic basis, evolutionary origin and spread of resistance to herbicides inhibiting acetolactate synthase in common groundsel (Senecio vulgaris). Pest Manag. Sci. 2016, 57, 1119–1126. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S.B. Resistance to AHAS inhibitor herbicides: Current understanding. Pest Manag. Sci. 2014, 70, 1340–1350. [Google Scholar] [CrossRef] [PubMed]
- Massa, D.; Krenz, B.; Gerhards, R. Target-site resistance to ALS-inhibiting herbicides in Apera spica-venti populations is conferred by documented and previously unknown mutations. Weed Res. 2011, 51, 294–303. [Google Scholar] [CrossRef]
- Menegat, A.; Bailly, G.C.; Aponte, R.; Heinrich, G.M.T.; Sievernich, B.; Gerhards, R. Acetohydroxyacid synthase (AHAS) amino acid substitution Asp376Glu in Lolium perenne: Effect on herbicide efficacy and plant growth. J. Plant Dis. Prot. 2016, 123, 145–153. [Google Scholar] [CrossRef]
- Scarabel, L.; Panozzo, S.; Loddo, D.; Mathiassen, S.K.; Kristensen, M.; Kudsk, P.; Gitsopoulos, T.; Travlos, I.; Tani, E.; Chachalis, D.; et al. Diversified Resistance Mechanisms in Multi-Resistant Lolium spp. in Three European Countries. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Anthimidou, E.; Ntoanidou, S.; Madesis, P.; Eleftherohorinos, I. Mechanisms of Lolium rigidum multiple resistance to ALS- and ACCase-inhibiting herbicides and their impact on plant fitness. Pestic. Biochem. Physiol. 2020, 164, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 10 June 2021).
- Nagy, E.; Hegedűs, G.; Taller, J.; Kutasy, B.; Virág, E. Illumina sequencing of the chloroplast genome of common ragweed (Ambrosia artemisiifolia L.). Data Brief 2017, 15, 606–611. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Seefeldt, S.S.; Jensen, J.E.; Fuerst, E.P. Log-Logistic Analysis of Herbicide Dose-Response Relationships. Weed Technol. 1995, 9, 218–227. [Google Scholar] [CrossRef]
- Knezevic, S.Z.; Streibig, J.C.; Ritz, C. Utilizing R Software Package for Dose-Response Studies: The Concept and Data Analysis. Weed Technol. 2007, 21, 840–848. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J. LA rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Westwood, J.H.; Charudattan, R.; Duke, S.O.; Fennimore, S.A.; Marrone, P.; Slaughter, D.C.; Swanton, C.; Zollinger, R. Weed Management in 2050: Perspectives on the Future of Weed Science. Weed Sci. 2018, 66, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Bajwa, A.; Mahajan, G.; Chauhan, B.S. Nonconventional Weed Management Strategies for Modern Agriculture. Weed Sci. 2015, 63, 723–747. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Li, M.; Purba, E.; Walsh, M.J.; Powles, S.B. Resistance evaluation for herbicide resistance-endowing acetolactate synthase (ALS) gene mutations using Raphanus raphanistrum populations homozygous for specific ALS mutations. Weed Res. 2012, 52, 178–186. [Google Scholar] [CrossRef]
- Saari, L.L.; Cotterman, J.C.; Primiani, M.M. Mechanism of Sulfonylurea Herbicide Resistance in the Broadleaf Weed, Kochia scoparia. Plant Physiol. 1990, 93, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, W.; Pan, L.; Li, J.; Wang, Q.; Feng, Y.; Dong, L. Molecular basis of ALS- and/or ACCase-inhibitor resistance in shortawn foxtail (Alopecurus aequalis Sobol.). Pestic. Biochem. Physiol. 2015, 122, 76–80. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutasy, B.; Takács, Z.; Kovács, J.; Bogaj, V.; Razak, S.A.; Hegedűs, G.; Decsi, K.; Székvári, K.; Virág, E. Pro197Thr Substitution in Ahas Gene Causing Resistance to Pyroxsulam Herbicide in Rigid Ryegrass (Lolium Rigidum Gaud.). Sustainability 2021, 13, 6648. https://doi.org/10.3390/su13126648
Kutasy B, Takács Z, Kovács J, Bogaj V, Razak SA, Hegedűs G, Decsi K, Székvári K, Virág E. Pro197Thr Substitution in Ahas Gene Causing Resistance to Pyroxsulam Herbicide in Rigid Ryegrass (Lolium Rigidum Gaud.). Sustainability. 2021; 13(12):6648. https://doi.org/10.3390/su13126648
Chicago/Turabian StyleKutasy, Barbara, Zsolt Takács, Judit Kovács, Verëlindë Bogaj, Syafiq A. Razak, Géza Hegedűs, Kincső Decsi, Kinga Székvári, and Eszter Virág. 2021. "Pro197Thr Substitution in Ahas Gene Causing Resistance to Pyroxsulam Herbicide in Rigid Ryegrass (Lolium Rigidum Gaud.)" Sustainability 13, no. 12: 6648. https://doi.org/10.3390/su13126648
APA StyleKutasy, B., Takács, Z., Kovács, J., Bogaj, V., Razak, S. A., Hegedűs, G., Decsi, K., Székvári, K., & Virág, E. (2021). Pro197Thr Substitution in Ahas Gene Causing Resistance to Pyroxsulam Herbicide in Rigid Ryegrass (Lolium Rigidum Gaud.). Sustainability, 13(12), 6648. https://doi.org/10.3390/su13126648