
Total Replacement of Fishmeal by Spirulina (Arthrospira platensis) and Its Effect on Growth Performance and Product Quality of African Catfish (Clarias gariepinus)

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Clearance
2.2. Rearing of Experimental Fish
2.3. Sampling of Experimental Fish
2.4. Color Measurements of Fish
2.5. Sampling of Experimental Fish
2.6. Fatty Acids
2.6.1. Chemical and Reagents
2.6.2. Fish Samples Treatment
2.6.3. Feed Samples Treatment
2.6.4. GC-FID Analysis of FAMEs
2.7. Statistical Analysis
3. Results
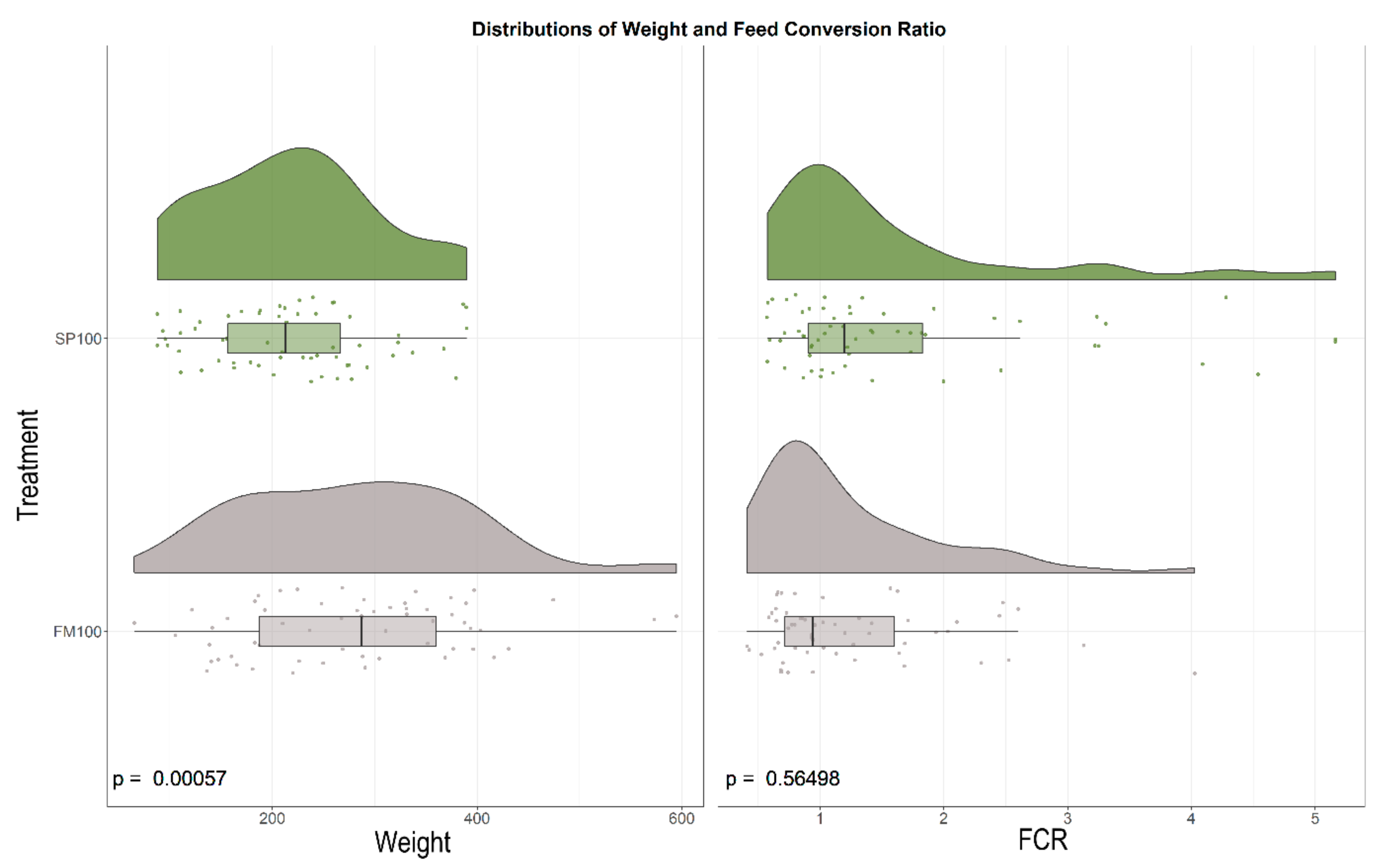
3.1. Growth Parameters
3.2. Color Parameters and Cooking Loss
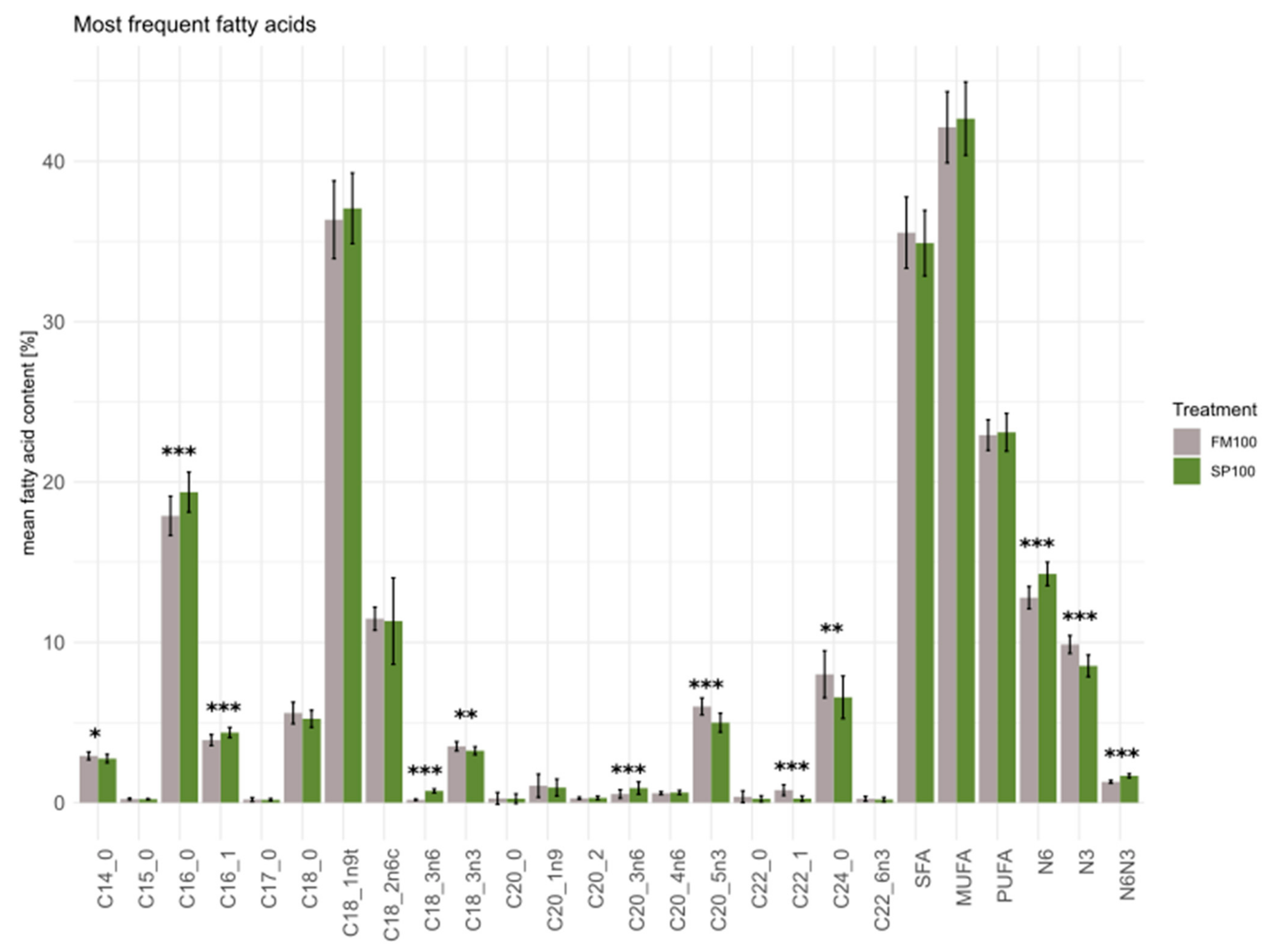
3.3. Fatty Acids
3.4. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Weight [g] | Length [mm] | Carcass Weight [g] | Approximate Feed Conversion Rate (FCR) | Daily Weight Gain [g] |
|---|---|---|---|---|---|
| FM100 | 280.84 ± 110.80 | 315.85 ± 38.84 | 256.67 ± 99.91 | 0.97 ± 1.89 | 3.31 ± 1.59 |
| SP100 | 218.66 ± 82.24 | 291.33 ± 33.31 | 199.61 ± 72.87 | 1.16 ± 1.16 | 2.41 ± 1.18 |
| df | 108.74 | 115.32 | 107.93 | 97.921 | 108.74 |
| t-Statistics | 3.550 | 3.711 | 3.617 | −0.577 | 3.550 |
| p-Value | <0.001 | <0.001 | <0.001 | 0.565 | <0.001 |
| Weeks | 0–2 | 2–6 | 6–10 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Diet | Weight [g] | FCR | Daily Weight Gain [g] | Weight [g] | FCR | Daily Weight Gain [g] | Weight [g] | FCR | Daily Weight Gain [g] |
| FM100 | 74.47 ± 0.64 | 0.67 ± 0.03 | 1.714 ± 0.08 | 169.33 ± 8.81 | 0.66 ± 0.05 | 3.388 ± 0.25 | 280.84 ± 13.19 | 1.33 ± 0.19 | 3.982 ± 0.60 |
| SP100 | 71.07 ± 1.06 | 0.75 ± 0.03 | 1.526 ± 0.07 | 147.95 ± 5.87 | 0.78 ± 0.04 | 2.756 ± 0.16 | 218.66 ± 41.57 | 1.72 ± 0.28 | 2.526 ± 0.44 |
| df | 2.606 | 4.000 | 3.753 | 3.588 | 3.986 | 3.439 | 2.399 | 3.561 | 3.671 |
| t-Statistics | 3.396 | −3.084 | 3.053 | 3.506 | −3.245 | 3.738 | 2.469 | −1.991 | 3.412 |
| p-Value | 0.053 | 0.037 | 0.041 | 0.030 | 0.032 | 0.0264 | 0.111 | 0.126 | 0.031 |
| Fatty Acid | FM100 | SP100 | p-Value |
|---|---|---|---|
| C6:0 | 0.00 ± 0.00 | 0.00 ± 0.00 | NA |
| C8:0 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.586 |
| C10:0 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.690 |
| C11:0 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.622 |
| C12:0 | 0.07 ± 0.02 | 0.07 ± 0.01 | 0.957 |
| C13:0 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.700 |
| C14:0 | 2.92 ± 0.24 | 2.75 ± 0.26 | 0.026 |
| C14:1 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.247 |
| C15:0 | 0.23 ± 0.06 | 0.22 ± 0.03 | 0.535 |
| C15:1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.834 |
| C16:0 | 17.89 ± 1.22 | 19.37 ± 1.25 | <0.001 |
| C16:1 | 3.90 ± 0.34 | 4.37 ± 0.32 | <0.001 |
| C17:0 | 0.20 ± 0.12 | 0.18 ± 0.07 | 0.639 |
| C17:1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.155 |
| C18:0 | 5.59 ± 0.67 | 5.23 ± 0.53 | 0.061 |
| C18:1n9 | 36.36 ± 2.42 | 37.07 ± 2.20 | 0.312 |
| C18:2n6c | 11.48 ± 0.72 | 11.33 ± 0.64 | 0.811 |
| C18:3n6 | 0.18 ± 0.05 | 0.73 ± 0.13 | <0.001 |
| C18:3n3 | 3.52 ± 0.29 | 3.24 ± 0.25 | 0.001 |
| C20:0 | 0.27 ± 0.37 | 0.25 ± 0.30 | 0.806 |
| C20:1n9 | 1.06 ± 0.72 | 0.95 ± 0.53 | 0.580 |
| C20:2 | 0.27 ± 0.07 | 0.30 ± 0.11 | 0.389 |
| C20:3n6 | 0.54 ± 0.27 | 0.91 ± 0.39 | <0.001 |
| C21:0 | 0.00 ± 0.01 | 0.00 ± 0.00 | 0.223 |
| C20:4n6 | 0.59 ± 0.10 | 0.63 ± 0.13 | 0.195 |
| C20:3n3 | 0.11 ± 0.02 | 0.10 ± 0.03 | 0.264 |
| C20:5n3 | 6.00 ± 0.53 | 4.99 ± 0.59 | <0.001 |
| C22:0 | 0.37 ± 0.36 | 0.24 ± 0.18 | 0.099 |
| C22:1 | 0.78 ± 0.34 | 0.26 ± 0.17 | <0.001 |
| C22:2 | 0.00 ± 0.00 | 0.00 ± 0.00 | NA |
| C23:0 | 0.00 ± 0.00 | 0.00 ± 0.00 | NA |
| C24:0 | 8.01 ± 1.47 | 6.58 ± 1.33 | 0.001 |
| C24:1n9 | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.324 |
| C22:6n3 | 0.24 ± 0.15 | 0.20 ± 0.13 | 0.277 |
| SFA | 35.55 ± 2.22 | 34.90 ± 2.04 | 0.299 |
| MUFA | 42.12 ± 2.22 | 42.66 ± 2.29 | 0.428 |
| PUFA | 22.93 ± 0.95 | 23.10 ± 1.18 | 0.604 |
| Total n-6 | 12.79 ± 0.70 | 14.27 ± 0.73 | <0.001 |
| Total n-3 | 9.87 ± 0.56 | 8.53 ± 0.68 | <0.001 |
| n-6/n-3 | 1.30 ± 0.09 | 1.68 ± 0.12 | <0.001 |
| Fatty Acid | FM100 | SP100 |
|---|---|---|
| C6:0 | 0.00 ± 0.00 | 0.15 ± 0.00 |
| C8:0 | 0.02 ± 0.02 | 0.10 ± 0.02 |
| C10:0 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| C11:0 | 0.00 ± 0.00 | 0.28 ± 0.00 |
| C12:0 | 0.06 ± 0.02 | 0.11 ± 0.02 |
| C13:0 | 0.01 ± 0.00 | 0.05 ± 0.00 |
| C14:0 | 3.65 ± 0.33 | 4.59 ± 0.33 |
| C14:1 | NA | NA |
| C15:0 | 0.24 ± 0.02 | 0.54 ± 0.02 |
| C15:1 | NA | NA |
| C16:0 | 12.60 ± 0.35 | 19.03 ± 0.35 |
| C16:1 | 3.93 ± 0.26 | 4.85 ± 0.26 |
| C17:0 | 0.20 ± 0.39 | 0.45 ± 0.39 |
| C17:1 | 0.09 ± 0.00 | 0.09 ± 0.00 |
| C18:0 | 2.61 ± 0.18 | 3.73 ± 0.18 |
| C18:1n9 | 36.80 ± 2.25 | 44.75 ± 2.25 |
| C18:2n6c | 15.77 ± 0.53 | 11.81 ± 0.16 |
| C18:3n6 | 0.12 ± 0.02 | 1.19 ± 0.05 |
| C18:3n3 | 4.83 ± 0.36 | 1.34 ± 0.04 |
| C20:0 | 0.58 ± 0.04 | 1.80 ± 0.03 |
| C20:1n9 | 1.87 ± 0.13 | 1.96 ± 0.05 |
| C20:2 | 0.10 ± 0.01 | 0.06 ± 0.01 |
| C20:3n6 | 0.07 ± 0.02 | 0.06 ± 0.00 |
| C21:0 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| C20:4n6 | 0.56 ± 0.06 | 0.12 ± 0.01 |
| C20:3n3 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| C20:5n3 | 8.58 ± 0.63 | 0.62 ± 0.05 |
| C22:0 | 0.22 ± 0.06 | 0.59 ± 0.28 |
| C22:1 | 1.24 ± 0.15 | 0.50 ± 0.28 |
| C22:2 | NA | NA |
| C23:0 | NA | NA |
| C24:0 | 5.18 ± 0.24 | 0.61 ± 0.03 |
| C24:1n9 | 0.00 ± 0.00 | 0.00 ± 0.01 |
| C22:6n3 | 0.67 ± 0.16 | 0.58 ± 0.12 |
| SFA | 25.37 ± 0.54 | 32.05 ± 0.28 |
| MUFA | 43.93 ± 1.97 | 52.16 ± 0.39 |
| PUFA | 30.60 ± 1.48 | 15.73 ± 0.17 |
| Total n-6 | 16.51 ± 0.60 | 13.19 ± 0.20 |
| Total n-3 | 14.09 ± 0.88 | 2.54 ± 0.12 |
| n-6/n-3 | 1.17 ± 0.30 | 5.21 ± 0.03 |
References
- Lovejoy, T.E.; Hannah, L.J.; Ainley, D.G.; Wilson, E.O. Biodiversity and Climate Change: Transforming the Biosphere; Yale University Press: London, UK, 2019; ISBN 9780300206111. [Google Scholar]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Mooney, H.; Larigauderie, A.; Cesario, M.; Elmquist, T.; Hoegh-Guldberg, O.; Lavorel, S.; Mace, G.M.; Palmer, M.; Scholes, R.; Yahara, T. Biodiversity, climate change, and ecosystem services. Curr. Opin. Environ. Sustain. 2009, 1, 46–54. [Google Scholar] [CrossRef]
- Rijnsdorp, A.D.; Peck, M.A.; Engelhard, G.H.; Möllmann, C.; Pinnegar, J.K. Resolving the effect of climate change on fish populations. ICES J. Mar. Sci. 2009, 66, 1570–1583. [Google Scholar] [CrossRef]
- Cheung, W.W.L.; Lam, V.W.Y.; Sarmiento, J.L.; Kearney, K.; Watson, R.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Booth, D.J.; Poloczanska, E.; Donelson, J.M.; Molinos, J.G.; Burrows, M. Biodiversity and Climate Change in the Oceans. In Climate Change Impacts on Fisheries and Aquaculture; Phillips, B.F., Pérez-Ramírez, M., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 63–89. [Google Scholar]
- Lawrence, A.J.; Arukwe, A.; Moore, M.; Sayer, M.; Thain, J. Molecular/Cellular Processes and the Physiological Response to Pollution. In Effects of Pollution on Fish; Lawrence, A., Hemingway, K., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; pp. 83–133. [Google Scholar]
- Austin, B. The effects of pollution on fish health. J. Appl. Microbiol. 1998, 85 (Suppl. 1), 234–242. [Google Scholar] [CrossRef]
- Heath, A.G. Water Pollution and Fish Physiology, 2nd ed.; CRC: Boca Raton, FL, USA, 2018; ISBN 9780203718896. [Google Scholar]
- Khoshnood, Z. Effects of Environmental Pollution on Fish: A Short Review. Transylv. Rev. Syst. Ecol. Res. 2017, 19, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Little, D.C.; Edwards, P. Integrated Livestock-Fish Farming Systems; FAO: Rome, Italy, 2003; ISBN 92-5-105055-4. [Google Scholar]
- García, S.M. The Ecosystem Approach to Fisheries: Issues, Terminology, Principles, Institutional Foundations, Implementation and Outlook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2003; ISBN 92-5-104960-2. [Google Scholar]
- Taelman, S.E.; de Meester, S.; Roef, L.; Michiels, M.; Dewulf, J. The environmental sustainability of microalgae as feed for aquaculture: A life cycle perspective. Bioresour. Technol. 2013, 150, 513–522. [Google Scholar] [CrossRef]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Dernekbasi, S.; Unal, H.; Karayucel, I.; Aral, O. Effect of Dietary Supplementation of Different Rates of Spirulina (Spirulina platensis) on Growth and Feed Conversion in Guppy (Poecilia reticulate Peters, 1860). J. Anim. Vet. Adv. 2010, 9, 1395–1399. [Google Scholar]
- Velasquez, S.F.; Chan, M.A.; Abisado, R.G.; Traifalgar, R.F.M.; Tayamen, M.M.; Maliwat, G.C.F.; Ragaza, J.A. Dietary Spirulina (Arthrospira platensis) replacement enhances performance of juvenile Nile tilapia (Oreochromis niloticus). J. Appl. Phycol. 2016, 28, 1023–1030. [Google Scholar] [CrossRef]
- Teimouri, M.; Amirkolaie, A.K.; Yeganeh, S. The effects of dietary supplement of Spirulina platensis on blood carotenoid concentration and fillet color stability in rainbow trout (Oncorhynchus mykiss). Aquaculture 2013, 414–415, 224–228. [Google Scholar] [CrossRef]
- Sirakov, I.; Velichkova, K.; Nikolov, G. The effect of algae meal (Spirulina) on the growth performance and carcass parameters of rainbow trout (Oncorhynchus mykiss). J. BioSci. Biotech. 2012, 151–156. Available online: https://www.researchgate.net/publication/271326438_The_effect_of_algae_meal_Spirulina_on_the_growth_performance_and_carcass_parameters_of_rainbow_trout_Oncorhynchus_mykiss (accessed on 29 September 2020).
- Nandeesha, M.C.; Gangadhara, B.; Manissery, J.K.; Venkataraman, L.V. Growth performance of two Indian major carps, catla (Catlacatla) and rohu (Labeorohita) fed diets containing different levels of Spirulina platensis. Bioresour. Technol. 2001, 80, 117–120. [Google Scholar] [CrossRef]
- Raji, A.A.; Alaba, P.A.; Yusuf, H.; Abu Bakar, N.H.; Mohd Taufek, N.; Muin, H.; Alias, Z.; Milow, P.; Abdul Razak, S. Fishmeal replacement with Spirulina Platensis and Chlorella vulgaris in African catfish (Clarias gariepinus) diet: Effect on antioxidant enzyme activities and haematological parameters. Vet. Sci. Res. J. 2018, 119, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Promya, J.; Chitmanat, C. The effects of Spirulina platensis and Cladophora Algae on the Growth Performance, Meat Quality and Immunity Stimulating Capacity of the African Sharptooth Catfish (Clarias gariepinus). Int. J. Agric. Biol. 2011, 13, 77–82. [Google Scholar]
- Meng-Umphan, K. Growth performance, sex hormone levels and maturation ability of Pla Pho (Pangasius bocourti) fed with spirulina supplementary pellet and hormone application. Int. J. Agric. Biol. 2009, 11, 458–462. [Google Scholar]
- Roy, S.S.; Pal, R. Microalgae in Aquaculture: A Review with Special References to Nutritional Value and Fish Dietetics. Proc. Zool. Soc. 2015, 68, 1–8. [Google Scholar] [CrossRef]
- Miki, W.; Yamaguchi, K.; Konosu, S. Chemistry and utilization of plankton. VIII. Carotenoid composition of Spirulina maxima. Nippon Suisan Gakkaishi 1986, 52, 1225–1227. [Google Scholar] [CrossRef]
- Mathew, B.; Sankaranarayanan, R.; Nair, P.P.; Varghese, C.; Somanathan, T.; Amma, B.P.; Amma, N.S.; Nair, M.K. Evaluation of chemoprevention of oral cancer with Spirulina fusiformis. Nutr. Cancer 1995, 24, 197–202. [Google Scholar] [CrossRef]
- Hata, M.; Hata, M. Studies on Astaxanthin Formation in some Fresh-water Fishes. Tohoku J. Agric. Res. 1974, 24, 192–196. [Google Scholar]
- Kop, A.; Durmaz, Y. The effect of synthetic and natural pigments on the colour of the cichlids (Cichlasoma severum sp., Heckel 1840). Aquac. Int. 2008, 16, 117–122. [Google Scholar] [CrossRef]
- Roohani, A.M.; Abedian Kenari, A.; Fallahi Kapoorchali, M.; Borani, M.S.; Zoriezahra, S.J.; Smiley, A.H.; Esmaeili, M.; Rombenso, A.N. Effect of spirulina Spirulina platensis as a complementary ingredient to reduce dietary fish meal on the growth performance, whole-body composition, fatty acid and amino acid profiles, and pigmentation of Caspian brown trout (Salmo trutta caspius) juveniles. Aquac. Nutr. 2019, 25, 633–645. [Google Scholar] [CrossRef]
- Vílchez, C.; Forján, E.; Cuaresma, M.; Bédmar, F.; Garbayo, I.; Vega, J.M. Marine carotenoids: Biological functions and commercial applications. Mar. Drugs 2011, 9, 319–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welker, C.; de Negro, P.; Sarti, M. Green algal carotenoids and yellow pigmentation of rainbow trout fish. Aquac. Int. 2001, 9, 87–93. [Google Scholar] [CrossRef]
- Tapiero, H.; Nguyen Ba, G.; Couvreur, P.; Tew, K.D. Polyunsaturated fatty acids (PUFA) and eicosanoids in human health and pathologies. Biomed. Pharmacother. 2002, 56, 215–222. [Google Scholar] [CrossRef]
- Nakagawa, H. Effect of dietary algae on improvement of lipid metabolism in fish. Biomed. Pharmacother. 1997, 51, 345–348. [Google Scholar] [CrossRef]
- Jafari, S.M.A.; Rabbani, M.; Emtyazjoo, M.; Piryaei, F. Effect of dietary Spirulina platensis on fatty acid composition of rainbow trout (Oncorhynchus mykiss) fillet. Aquac. Int. 2014, 22, 1307–1315. [Google Scholar] [CrossRef]
- Teimouri, M.; Yeganeh, S.; Amirkolaie, A.K. The effects of Spirulina platensis meal on proximate composition, fatty acid profile and lipid peroxidation of rainbow trout (Oncorhynchus mykiss) muscle. Aquac. Nutr. 2016, 22, 559–566. [Google Scholar] [CrossRef]
- FAO. FAO Fisheries & Aquaculture—Species Fact Sheets—Clarias Gariepinus (Burchell, 1822). Available online: http://www.fao.org/fishery/species/2982/en (accessed on 29 September 2020).
- Dietz, C.; Sünder, A.; Liebert, F. Does Genetic Background of Rainbow Trout Impact on Growth and Feed Utilization Following Fishmeal Substitution by Partly Defatted Insect Meal (Hermetia illucens) or Microalgae Powder (Arthrospira platensis)?: 74rd Conference 3–5 March 2020 in Göttingen; DLG-Verlag: Frankfurt, Germany, 2020; ISBN 978-3-7690-4113-2. [Google Scholar]
- National Research Council. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-47322-4. [Google Scholar]
- Iwona, C.-S.; Skiepko, N.; Pomianowski, J.F.; Kubiak, M.S.; Baryczka, M. Gender differences in the chemical composition and selected properties of African Catfish (Clarias Gariepinus Burchell, 1822) Meat. Ital. J. Food Sci. 2016, 28, 2016–2391. [Google Scholar]
- Du, M.; Ahn, D.U.; Sell, J.L. Effects of dietary conjugated linoleic acid and linoleic:linolenic acid ratio on polyunsaturated fatty acid status in laying hens. Poult. Sci. J. 2000, 79, 1749–1756. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Germany, 2016; ISBN 9783319242774. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 29 September 2020).
- Allen, M.; Poggiali, D.; Whitaker, K.; Marshall, T.R.; Kievit, R.A. Raincloud plots: A multi-platform tool for robust data visualization. Wellcome Open Res. 2019, 4, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The State of World Fisheries and Aquaculture: Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018; ISBN 9789251305621. [Google Scholar]
- Olvera-Novoa, M.A.; Domínguez-Cen, L.J.; Olivera-Castillo, L.; Martínez-Palacios, C.A. Effect of the use of the microalga Spirulina maxima as fish meal replacement in diets for tilapia, Oreochromis mossambicus (Peters), fry. Aquac. Res. 1998, 29, 709–715. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Evaluation of soybean meal, spirulina meal and chicken offal meal as protein sources for silver seabream (Rhabdosargus sarba) fingerlings. Aquaculture 1994, 127, 169–176. [Google Scholar] [CrossRef]
- Chavez, P.D.L.C.; Bolviar, R.B. Spirulina platensis as an alternative protein source for the African catfish Clarias gariepinus. AACL Bioflux 2018, 11, 481–494. [Google Scholar]
- Raji, A.A.; Junaid, O.Q.; Milow, P.; Mohd. Taufek, N.; Fada, A.M.; Kolawole, A.A.; Alias, Z.; Razak, S.A. Partial replacement of fishmeal with Spirulina platensis and Chlorella vulgaris and its effect on growth and body composition of African catfish Clarias gariepinus (Burchell 1822). Indian J. Fish. 2019, 66, 100–111. [Google Scholar] [CrossRef]
- Choubert, G. Tenative utilization of Spirulina algae as a source of carotenoid pigments for Rainbow Trout. Aquaculture 1979, 18, 135–143. [Google Scholar] [CrossRef]
- Peterson, D.H.; Jäger, H.K.; Savage, G.M.; Washburn, G.N.; Westers, H. Natural Coloration of Trout Using Xanthophylls. Trans. Am. Fish. Soc. 1966, 95, 408–414. [Google Scholar] [CrossRef]
- Young, J.A.; Smith, A.P.; Muir, J.F. Market analysis using Multiple Correspondence Analysis: The case of consumer acceptance of African catfish, Clarias gariepinus (Burchell, 1822). Aquac. Res. 1996, 27, 283–291. [Google Scholar] [CrossRef]
- Jäger, G.-L. Entwicklung einer Marketing-Konzeption für Süßwasserfische aus Warmwasser-Kreislaufanlagen: Dargestellt am Beispiel Afrikanischer Wels. Ph.D. Thesis, Universität Göttingen, Göttingen, Germany, 1991. [Google Scholar]
- Li, Y.; Liu, S.; Cline, D.; Chen, S.; Wang, Y.; Bell, L.N. Chemical treatments for reducing the yellow discoloration of channel catfish (Ictalurus punctatus) fillets. J. Food Sci. 2013, 78, S1609–S1613. [Google Scholar] [CrossRef]
- Larsson, T.; Koppang, E.O.; Espe, M.; Terjesen, B.F.; Krasnov, A.; Moreno, H.M.; Rørvik, K.-A.; Thomassen, M.; Mørkøre, T. Fillet quality and health of Atlantic salmon (Salmo salar L.) fed a diet supplemented with glutamate. Aquaculture 2014, 426–427, 288–295. [Google Scholar] [CrossRef]
- Morkore, T.; Einen, O. Relating Sensory and Instrumental Texture Analyses of Atlantic Salmon. J. Food Sci. 2003, 68, 1492–1497. [Google Scholar] [CrossRef]
- Solomon, R.J. Proximate Analysis and Nutritional Value of African Catfish (Clarias gariepinus) Fed with Local (Telferia occidentales and Moringa olefera) and Industrial Feed (Coppens). J. Fish. Livest. Prod. 2018, 6, 1–6. [Google Scholar] [CrossRef]
- Chukwu, O.; Shaba, M.I. Effects of Drying Methods on Proximate Compositions of Catfish (Clarias gariepinus). World J. Agric. Sci. 2009, 5, 114–116. [Google Scholar]
- Akpambang, V.O.E. Proximate composition of some tropical fish species. Chem. Sin. 2015, 6, 125–129. [Google Scholar]
- Rosa, R.; Bandarra, N.M.; Nunes, M.L. Nutritional quality of African catfish Clarias gariepinus (Burchell 1822): A positive criterion for the future development of the European production of Siluroidei. Int. J. Food. Sci. Technol. 2007, 42, 342–351. [Google Scholar] [CrossRef]
- Offer, G.; Knight, P.; Jeacocke, R.; Almond, R.; Cousins, T.; Elsey, J.; Parsons, N.; Sharp, A.; Starr, R.; Purslow, P. The Structural Basis of the Water-Holding, Appearance and Toughness of Meat and Meat Products. Food Struct. 1989, 8, 151–170. [Google Scholar]
- Wedekind, H. Untersuchungen zur Produktqualität Afrikanischer Welse (Clarias gariepinus) in Abhängigkeit von genetischer Herkunft, Fütterung, Geschlecht und Schlachtalter. Ph.D. Thesis, Universität Göttingen, Göttingen, Germany, 1991. [Google Scholar]
- Palmegiano, G.B.; Agradi, E.; Forneris, G.; Gai, F.; Gasco, L.; Rigamonti, E.; Sicuro, B.; Zoccarato, I. Spirulina as a nutrient source in diets for growing sturgeon (Acipenser baeri). Aquac. Res. 2005, 36, 188–195. [Google Scholar] [CrossRef]
- Chainapong, T.; Traichaiyaporn, S.; Deming, R.L. Effect of dietary spirulina platensis on the fatty acid composition in flesh and ovary of walking catfish (Clarias macrocephalus). Chiang Mai J. Sci. 2018, 45, 129–135. [Google Scholar]
- Oboh, A.; Betancor, M.B.; Tocher, D.R.; Monroig, O. Biosynthesis of long-chain polyunsaturated fatty acids in the African catfish Clarias gariepinus: Molecular cloning and functional characterisation of fatty acyl desaturase (fads2) and elongase (elovl2) cDNAs7. Aquaculture 2016, 462, 70–79. [Google Scholar] [CrossRef]
- Passi, S.; Cataudella, S.; Di Marco, P.; de Simone, F.; Rastrelli, L. Fatty acid composition and antioxidant levels in muscle tissue of different Mediterranean marine species of fish and shellfish. J. Agric. Food Chem. 2002, 50, 7314–7322. [Google Scholar] [CrossRef] [PubMed]
- Elvevoll, E.O.; James, D.G. Potential benefits of fish for maternal, foetal and neonatal nutrition: A review of the literature. Food Nutr. Agric. 2000, 27, 28–39. [Google Scholar]




| Ingredient (% Dry Matter) | FM100 | SP100 |
|---|---|---|
| Fishmeal 1 | 20.00 | 0.00 |
| Spirulina 2 | 0.00 | 20.00 |
| Fish oil | 10.70 | 10.70 |
| Wheat meal | 14.00 | 12.50 |
| Wheat gluten | 20.00 | 21.50 |
| Soy protein concentrate 3 | 20.00 | 20.00 |
| Rapeseed oil | 10.70 | 10.70 |
| Vit./min. premix | 1.00 | 1.00 |
| CaHPO4 | 1.00 | 1.00 |
| CMC (Binder) | 1.29 | 1.08 |
| TiO2 (Marker) | 0.50 | 0.50 |
| Fe3O4—black (dye) | 0.07 | 0.07 |
| L-Lysin (HCL-Lys, 78% Lys) | 0.70 | 0.90 |
| D,L-Methionine | 0.01 | 0.04 |
| L-Tryptophan | 0.03 | 0.01 |
| Approximate Composition (%) | FM100 | SP100 |
|---|---|---|
| Dry matter | 94.6 | 94.0 |
| Crude protein (Nx6.25) | 45.4 | 45.7 |
| Crude lipids | 24.6 | 23.9 |
| N-free extracts | 17.5 | 19.0 |
| Crude ash | 7.1 | 5.4 |
| Gross energy [MJ/kg] | 23.4 | 23.5 |
| Digestible energy [MJ/kg] | 20.0 | 20.0 |
| Skin (Dorsal Back Muscle) | |||||
| Diet | L* | a* | b* | C* | h° |
| FM100 | 30.42 ± 4.92 | 1.17 ± 0.46 | 1.97 ± 1.42 | 2.40 ± 1.29 | 67.29 ± 63.92 |
| SP100 | 29.81 ± 4.63 | 0.77 ± 0.46 | 4.18 ± 2.04 | 4.30 ± 2.00 | 78.14 ± 17.43 |
| p-Value | 0.455 | <0.001 | <0.001 | <0.001 | 0.091 |
| Skin (Abdomen) | |||||
| Diet | L* | a* | b* | C* | h° |
| FM100 | 71.19 ± 8.28 | 1.56 ± 1.74 | 2.49 ± 2.53 | 3.53 ± 2.37 | 107.89 ± 100.11 |
| SP100 | 68.60 ± 7.45 | 0.61 ± 2.35 | 11.25 ± 3.95 | 11.46 ± 4.09 | 87.34 ± 10.46 |
| p-Value | 0.058 | 0.002 | <0.001 | <0.001 | 0.056 |
| Raw Fillet | |||||
| Diet | L* | a* | b* | C* | h° |
| FM100 | 48.66 ± 5.05 | 0.35 ± 1.69 | 9.15 ± 2.25 | 9.32 ± 2.23 | 89.90 ± 13.58 |
| SP100 | 47.98 ± 5.52 | 2.32 ± 2.17 | 13.17 ± 3.55 | 13.52 ± 3.66 | 82.00 ± 11.83 |
| p-Value | 0.247 | <0.001 | <0.001 | <0.001 | <0.001 |
| Cooked Fillet | |||||
| Diet | ΔL* | Δa* | Δb* | ΔC* | Δh° |
| FM100 | 24.36 ± 3.68 | 2.22 ± 1.28 | 8.34 ± 2.61 | 7.83 ± 2.41 | 16.58 ± 8.65 |
| SP100 | 24.19 ± 3.37 | 1.80 ± 1.09 | 8.83 ± 2.96 | 8.81 ± 2.96 | 7.19 ± 4.17 |
| p-Value | 0.690 | 0.003 | 0.142 | 0.037 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosenau, S.; Oertel, E.; Dietz, C.; Wessels, S.; Tetens, J.; Mörlein, D.; Ciulu, M. Total Replacement of Fishmeal by Spirulina (Arthrospira platensis) and Its Effect on Growth Performance and Product Quality of African Catfish (Clarias gariepinus). Sustainability 2021, 13, 8726. https://doi.org/10.3390/su13168726
Rosenau S, Oertel E, Dietz C, Wessels S, Tetens J, Mörlein D, Ciulu M. Total Replacement of Fishmeal by Spirulina (Arthrospira platensis) and Its Effect on Growth Performance and Product Quality of African Catfish (Clarias gariepinus). Sustainability. 2021; 13(16):8726. https://doi.org/10.3390/su13168726
Chicago/Turabian StyleRosenau, Simon, Elisa Oertel, Carsten Dietz, Stephan Wessels, Jens Tetens, Daniel Mörlein, and Marco Ciulu. 2021. "Total Replacement of Fishmeal by Spirulina (Arthrospira platensis) and Its Effect on Growth Performance and Product Quality of African Catfish (Clarias gariepinus)" Sustainability 13, no. 16: 8726. https://doi.org/10.3390/su13168726
APA StyleRosenau, S., Oertel, E., Dietz, C., Wessels, S., Tetens, J., Mörlein, D., & Ciulu, M. (2021). Total Replacement of Fishmeal by Spirulina (Arthrospira platensis) and Its Effect on Growth Performance and Product Quality of African Catfish (Clarias gariepinus). Sustainability, 13(16), 8726. https://doi.org/10.3390/su13168726