
Dynamic Role and Importance of Multi-Kingdom Communities in Mediterranean Wood-Pastures


 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Literature Review
3. Mediterranean Ecosystems
3.1. General Characteristics
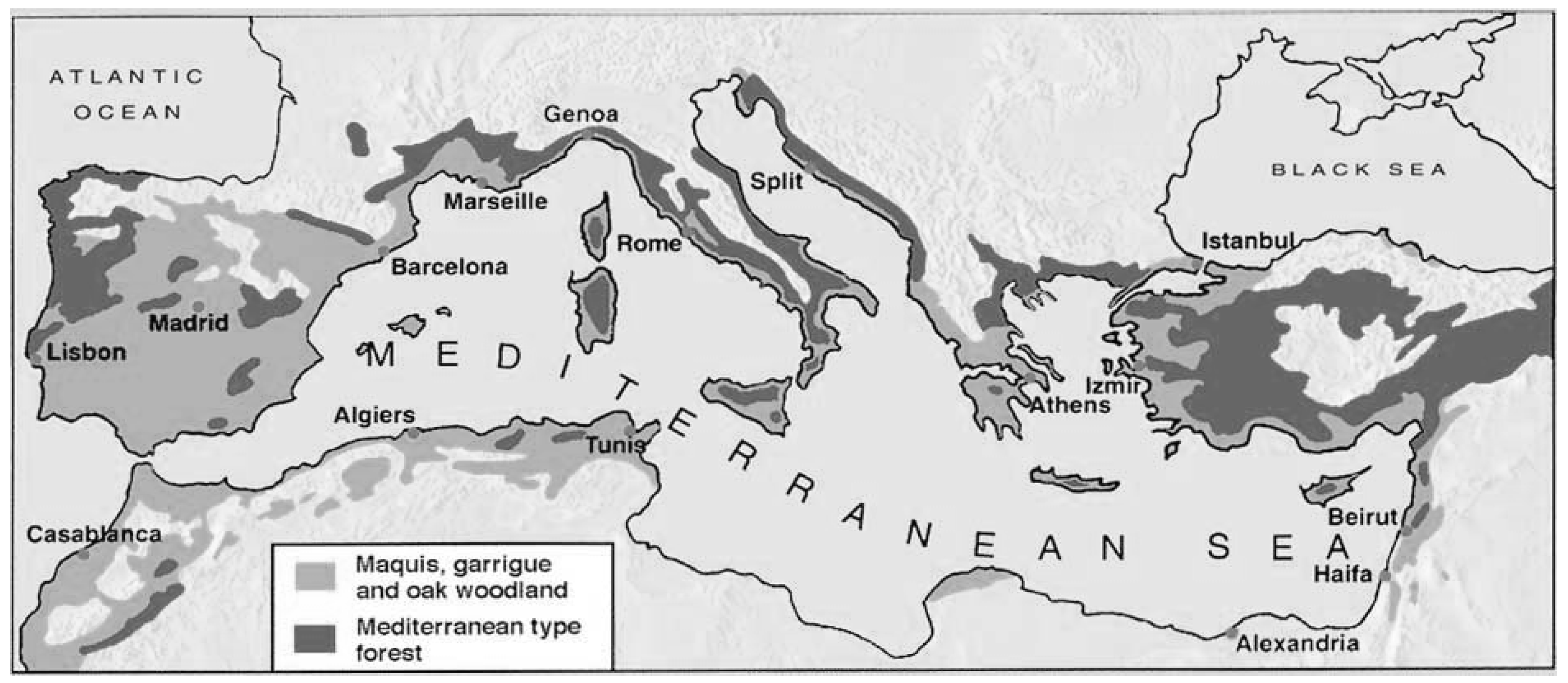
3.2. Mediterranean Basin
3.3. Dynamics of the Communities in the Wood-Pastures of Mediterranean Landscapes
3.4. Important Plant Families in Meadows
4. Grazing Management in Mediterranean Wood-Pastures
4.1. Role of Grazing in Plant Diversity of Mediterranean Landscapes
4.2. The importance of Fungi in Ecosystem Processes
4.2.1. The Dynamics of Fungal Communities in Mediterranean Wood-Pasture Landscapes
4.2.2. The Effect of Grazing on Fungal Biodiversity in Mediterranean Wood-Pastures
4.3. Effect of Wood Land Grazing on Ruminants
Products and PDO
- Meat
- Cheese
- Success stories of mountain ruminant farming
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Jalón, S.G.; Graves, A.; Moreno, G.; Palma, J.H.N.; Crous-Durán, J.; Kay, S.; Burgess, P.J. Forage-SAFE: A Model for Assessing the Impact of Tree Cover on Wood Pasture Profitability. Ecol. Model. 2018, 372, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Plieninger, T.; Levers, C.; Mantel, M.; Costa, A.; Schaich, H.; Kuemmerle, T. Patterns and drivers of scattered tree loss in agricultural landscapes: Orchard meadows in Germany (1968–2009). PLoS ONE 2015, 10, e0126178. [Google Scholar] [CrossRef]
- Solomou, A.D.; Sfougaris, A. Predicting Woody Plant Diversity as Key Component of Ecosystems: A Case Study in Central Greece. IJAEIS 2019, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Solomou, A.D.; Sfougaris, A. Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea Europaea L.) in the Mediterranean Landscape. Agronomy 2021, 11, 161. [Google Scholar] [CrossRef]
- Plieninger, T.; Höchtl, F.; Spek, T. Traditional land-use and nature conservation in European rural landscapes. Environ. Sci. Policy 2006, 9, 317–321. [Google Scholar] [CrossRef]
- Stoate, C.; Báldi, A.; Beja, P.; Boatman, N.D.; Herzon, I.; van Doorn, A.; de Snoo, G.R.; Rakosy, L.; Ramwell, C. Ecological impacts of early 21st century agricultural change in Europe—A review. J. Environ. Manag. 2009, 91, 22–46. [Google Scholar] [CrossRef] [PubMed]
- Sollen-Norrlin, M.; Ghaley, B.B.; Rintoul, N.L.J. Agroforestry Benefits and Challenges for Adoption in Europe and Beyond. Sustainability 2020, 12, 7001. [Google Scholar] [CrossRef]
- Hartel, T.; Plieninger, T. European Wood-Pastures in Transition Asocial-Ecological Approach; Hartel, T., Plieninger, T., Eds.; Fish Books Routledge: London, UK; New York, NY, USA, 2014. [Google Scholar]
- Garbarino, M.; Lingua, E.; Subirà, M.M.; Motta, R. The larch wood pasture: Structure and dynamics of a cultural landscape. Eur. J. For. Res. 2011, 130, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Bergmeier, E.; Petermann, J.; Schröder, E. Geobotanical survey of wood-pasture habitats in Europe: Diversity, threats and conservation. Biodivers. Conserv. 2010, 19, 2995–3014. [Google Scholar] [CrossRef] [Green Version]
- Ferraz-de-Oliveira, M.I.; Azeda, C.; Pinto-Correia, T. Management of Montados and Dehesas for High Nature Value: An Interdisciplinary Pathway. Agrofor. Syst. 2016, 90, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bernues, A.; Rodríguez-Ortega, T.; Ripoll-Bosch, R.; Alfnes, F. Socio-Cultural and Economic Valuation of Ecosystem Services Provided by Mediterranean Mountain Agroecosystems. PLoS ONE 2014, 9, e102479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Clara, M.I.E.; de Almeida Ribeiro, N.M.C. Decline of Mediterranean Oak Trees and Its Association with Phytophthora Cinnamomi: A Review. Eur. J. For. 2013, 132, 411–432. [Google Scholar] [CrossRef]
- Charvalas, G.; Solomou, A.D.; Giannoulis, K.D.; Skoufogianni, E.; Bartzialis, D.; Emmanouil, C.; Danalatos, N.G. Determination of Heavy Metals in the Territory of Contaminated Areas of Greece and Their Restoration through Hyperaccumulators. Environ. Sci. Pollut. Res. 2021, 28, 3858–3863. [Google Scholar] [CrossRef] [PubMed]
- Di Castri, F. Mediterranean-type shrublands of the world. In Ecosystems of the World: Mediterranean-Type Shrublands; Di Castri, F., Goodall, D.W., Specht, L.R., Eds.; Elsevier: Amsterdam, The Netherlands, 1981. [Google Scholar]
- Hobbs, R.J.; Richardson, D.M.; Davis, G.W. Mediterranean-type ecosystems: Opportunities and constraints for studying the function of biodiversity. In Mediterranean Type Ecosystems: The Function of Biodiversity; Davis, G.W., Richardson, D.M., Eds.; Springer: Berlin, Germany, 1995. [Google Scholar]
- Orshan, G. Approaches to the definition of Mediterranean growth forms. In Mediterranean-Type Ecosystems: The Role of Nutrients; Kruger, F.J., Mitchell, D.T., Jarvis, J.U.M., Eds.; Springer: Berlin, Germany, 1983. [Google Scholar]
- Aschmann, H. Distribution and peculiarity of Mediterranean ecosystems. In Mediterranean Type Ecosystems; Di Castri, F., Mooney, H., Eds.; Chapman & Hall: London, UK, 1973; pp. 11–19. [Google Scholar]
- Polade, S.D.; Gershunov, A.; Cayan, D.R.; Dettinger, M.D.; Pierce, D.W. Precipitation in a Warming World: Assessing Projected Hydro-Climate Changes in California and Other Mediterranean Climate Regions. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Vamvaka, N. Mediterranean Type Ecosystems. Bachelor’s Thesis, Department of Environment, University of Aegean, Mytilene, Greece, 2004. [Google Scholar]
- Kazakou, E. The Effect of Ecological Factors Key to Development of Vegetation Mediterranean Ecosystems. Bachelor’s Thesis, Environment Section, University of the Aegean, Mytilene, Greece, 2001. [Google Scholar]
- Raposo, M.A.; Gomes, C.J.P.; Nunes, L.J. Selective Shrub Management to Preserve Mediterranean Forests and Reduce the Risk of Fire: The Case of Mainland Portugal. Fire 2020, 3, 65. [Google Scholar] [CrossRef]
- Blondel, J.; Aronson, J. Biodiversity and ecosystem function in the Mediterranean basin: Human and non-human determinants. In Mediterranean-Type Ecosystems: The Function of Biodiversity; Davis, G.W., Richardson, D.M., Eds.; Springer: Berlin, Germany, 1995. [Google Scholar]
- Mittermeier, R.A.; Gil, P.R.; Hoffmann, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.G.; Lamoreux, J.; Da Fonseca, G.A.B. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; University of Chicago Press: Chicago, IL, USA, 2004. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Kark, S.; Levin, N.; Grantham, H.; Possingham, H. Between-country collaboration and consideration of costs increase conservation planning efficiency in the Mediterranean Basin. Proc. Nat. Acad. Sci. USA 2009, 106, 15360–15365. [Google Scholar] [CrossRef] [Green Version]
- Naveh, Z.; Dan, J. The human degradation of Mediterranean landscape in Israel. In Mediterranean Type Ecosystems, Origin and Structures; di Castri, F., Mooney, H.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1973; pp. 373–390. [Google Scholar]
- Turner, M.G. Landscape ecology: The effect of pattern on process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Levin, S.A. The problem of pattern and scale in ecology. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Peterson, G.D. Contagious disturbance, ecological memory, and the emergence of landscape pattern. Ecosystems 2002, 5, 239–338. [Google Scholar] [CrossRef]
- Lieberman, D.; Lieberman, M.; Hartshorn, G.S.; Peralta, R. Growth rates and age-size relationships of tropical wet forest trees in Costa Rica. J. Trop. Ecol. 1985, 1, 97–109. [Google Scholar] [CrossRef]
- Aquino, F.G.; Oliveira, M.C.; Schiavini, I.; Ribeiro, J.F. Dinâmica de populações de Anadenanthera macrocarpa e Acacia glomerosa em mata seca semidecídua na Estação Ecológica do Panga (Uberlândia-MG). Boletim Herbário Ezechias Paulo Heringer 1999, 4, 90–102. [Google Scholar]
- Naveh, Z. The evolutionary significance of fire in the Mediterranean region. Vegetatio 1975, 29, 199–208. [Google Scholar] [CrossRef]
- Oakley, K.P. Fire as a Palaeolithic tool and weapon. Proc. Prehist. Soc. 1955, 21, 36–47. [Google Scholar] [CrossRef]
- Perlés, C. Préhistoire du Feu; Masson: París, France, 1977. [Google Scholar]
- Trabaud, L. Fire and survival traits of plants. In The Role of Fire in Ecological Systems; Trabaud, L., Ed.; SPB Acedemic Publishing: The Hague, The Netherlands, 1987; pp. 91–95. [Google Scholar]
- Stapert, D.; Johansen, L. Making fire in the Stone Age: Flint and pyrite. Geol. Mijnb. 1999, 78, 147–164. [Google Scholar] [CrossRef]
- Farina, A. Principles and Methods in Landscape Ecology; Chapman & Hall: London, UK, 1998. [Google Scholar]
- Moreira, A.; Rego, F.C.; Ferreira, P.G. Temporal (1958–1995) pattern of change in a cultural landscape of northwestern Portugal: Implications for fire occurrence. Landscape Ecol. 2001, 16, 557–567. [Google Scholar] [CrossRef]
- Pausas, J.G.; Ramos, J.I. Landscape pattern, fire regime and vegetation dynamics—A modelling approach. In Ecology, Conservation and Management of Mediterranean Climate Ecosystems of the World, Proceedings of the MEDECOS 10th International Conference, Rhodes, Greece; Arianoutsou, M., Papanastasis, V.P., Eds.; Millpress: Rotterdam, The Netherlands, 2004. [Google Scholar]
- Sarlis, G.P. Improvement and Management of Natural Pastures Part A; Stamoulis Press: Athens, Greece, 1998. [Google Scholar]
- Tsibrakou, K. The Production of Viscose Material of Meadow “Kostilata” Theodorian Artas; Department of Agriculture and Food Technology: Arta, Greece, 2017. [Google Scholar]
- Arianoutsou, M. Biological Activity after Fire in a Phryganic Ecosystem. Ph.D. Thesis, University of Thessaloniki, Thessaloniki, Greece, 1979. (In Greek with an English Summary). [Google Scholar]
- Johnson, E.A. Fire and Vegetation Dynamics: Studies from the North American Boreal Forest; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Zedler, P.H.; Gautier, C.R.; McMaster, G.S. Vegetation change in response to extreme events: The effect of a short interval between fires in California chaparral and coastal scrub. Ecology 1983, 64, 809–818. [Google Scholar] [CrossRef]
- D¢Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle and global change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Lloret, F.; Pausas, J.G.; Vila, M. Response of Mediterranean plant species to different fire regimes in Garraf Natural Park (Catalonia, Spain): Field observations and modelling predictions. Plant Ecol. 2003, 167, 223–235. [Google Scholar] [CrossRef]
- Arianoutsou, M. Annual Report to the General Secretariat for Research and Technology Research Pened. In Program Entitled: Assessment of Mediterranean Ecosystems Desertification Trends as a Result of the Action of Fire; National and Kapodistrian University of Athens: Athens, Greece, 1998; p. 114. [Google Scholar]
- Trabaud, L. Post-fire community dynamics in the Mediterranean Basin. In The Role of Fire in Mediterranean-Type Ecosystems; Moreno, J.M., Oechel, W.C., Eds.; Ecological Studies 107; Springer: New York, NY, USA, 1994; pp. 1–15. [Google Scholar]
- Kazanis, D.; Arianoutsou, Ì. Vegetation composition in a post-fire successional gradient of Pinus halepensis forests of Attica, Greece. Int. J. Wildland Fire 1996, 6, 83–91. [Google Scholar] [CrossRef]
- Arianoutsou, M. Aspects of demography in post-fire plant communities of Greece. In Landscape Degradation and Biodiversity in Mediterranean Type Ecosystems; Rundel, P.W., Montenegro, G., Jaksic, F., Eds.; Ecological Studies 136; Springer: Berlin/Heidelberg, Germany, 1998; pp. 273–295. [Google Scholar]
- Arianoutsou, M.; Neéman, G. Post-fire regeneration of natural Pinus halepensis forests in the East Mediterranean Basin. In Ecology, Biogeography and Management of Mediterranean Pine Forests; Neéman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 269–290. [Google Scholar]
- Papanastasis, V.P. Fire ecology and management of phrygana communities in Greece: H.A. Mooney and C.E. Conrad (techn. coord.). In Proceedings of the Symposium on the Environmental Consequences of Fire and Fuel Management in Mediterranean Ecosystems, Palo Alto, CA, USA, 1–5 August 1977; pp. 476–482. [Google Scholar]
- Arianoutsou, M. Post-fire successional recovery of a phryganic (east Mediterranean) ecosystem. Acta Oecologica Oecologia Plant. 1984, 5, 387–394. [Google Scholar]
- Thanos, C.; Marcou, S.; Christodoulakis, D.; Yannitsaros, A. Early post-fire regeneration in Pinus brutia forest ecosystem of Samos island (Greece). Acta Oecologica Oecologia Plant. 1989, 10, 79–94. [Google Scholar]
- Hanes, T.L. Succession after fire in the chaparral of southern California. Ecol. Monogr. 1971, 41, 27–52. [Google Scholar] [CrossRef]
- Kruger, F.J. Plant community diversity and dynamics in relation to fire. In Mediterranean Type Ecosystems; Kruger, F.J., Mitchell, D.T., Jarvis, J.U., Eds.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 446–472. [Google Scholar]
- Papanastasis, V.; Vrachnakis, M.; Iovi, K.; Platis, P.; Karalazos, A.; Hatziminaoglou, I. Dynamics of herbage production in a grazed of mountain grassland in Greece. Multifunction grasslands: Quality forage, animal products and landcapes. In Proceedings of the 19th General Meeting of the European Grassland Federation, La Rochelle, France, 27–30 May 2002; Volume 7, pp. 348–349. [Google Scholar]
- Solomou, A.D.; Karetsos, G.; Skoufogianni, E.; Martinos, K.; Sfougaris, A.; Tsagari, K. Assessment of Greek forests protection and management. In Sustainable Development in Mountain Regions; Springer: Berlin/Heidelberg, Germany, 2016; pp. 141–166. [Google Scholar]
- Noy-Meir, I. Effects of grazing on Mediterranean grasslands: The community level. In Proceeding of the International Workshop of European Grassland Federation, Ecological Basis of Livestock Grazing in Mediterranean Ecosystems, Thessaloniki, Greece, 23–25 October 1997; pp. 27–39. [Google Scholar]
- Thirwood, J.V. Man and the Mediterranean Forest; Academic Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Tsoumis, G. The Depletion of Forests in the Mediterranean Region a Historical Review from the Ancient Times to the Present; Scientific Annals of the Department of Forestry and Natural Environment: Thessaloniki, Greece, 1985; p. 281300. [Google Scholar]
- Vokou, D.; Diamantopoulos, J.; Mardiris, T.A.; Margaris, N.S. Desertification in northern Greece the case of Kella. In Desertification in Europe; Fantechi, R., Margaris, N.S., Eds.; D. Reidel Publishing Co.: Dordrecht, The Netherlands, 1986; p. 155160. [Google Scholar]
- Pantis, J.; Margaris, N.S. Can system dominated by asphodels be considered as semideserts? Int. J. Biometeorol. 1988, 32, 8791. [Google Scholar] [CrossRef]
- Papanastasis, V.; Kyriakakis, S.; Ispikoudis, J. Forestry and grazing practices in Crete. In Stability and Change in the Cretan Landscape; Grove, A.T., Moody, J., Rackham, O., Eds.; Petromaroula 1; Corpus Christi College: Cambridge, UK, 1990; p. 446. [Google Scholar]
- Anderson, V.J.; Briske, D.D. Herbivore-induced species replacement in grasslands: Is it driven by herbivory tolerance or avoidance? Ecol. Appl. 1995, 5, 1014–1024. [Google Scholar] [CrossRef]
- Caldwell, M.M.; Richards, J.H.; Manwaring, J.H.; Eissenstat, D.M. Rapid shifts in phosphate acquisition show direct competition between neighboring plants. Nature 1987, 327, 615–616. [Google Scholar] [CrossRef]
- Gomez Sal, A.; De Miguel, J.M.; Casado, M.A.; Pineda, F.D. Successional changes in the morphology and ecological responses of a grazed pasture ecosystem in Central Spain. Vegetatio 1986, 67, 33–44. [Google Scholar]
- Fernandez Ales, R.; Laffarga, J.M.; Ortega, F. Strategies in Mediterranean grassland annuals in relation to stress and disturbance. J. Veg. Sci. 1993, 4, 313–322. [Google Scholar] [CrossRef]
- Dale, V.H.; Beyeler, S.C. Challenges in the development and use of ecological indicators. Ecol. Indic. 2001, 1, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Dale, V.H.; Beyeler, S.C.; Jackson, B. Understory vegetation indicators of anthropogenic disturbance in longleaf pine forest at Fort Benning, Georgia, USA. Ecol. Indic. 2002, 1, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- White, P.S. Pattern, process, and natural disturbance in vegetation. Bot. Rev. 1979, 45, 229–299. [Google Scholar] [CrossRef]
- Huston, M.A. Biological Diversity: The Coexistence of Species on Changing; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Ferris–Kaan, R.; Peace, A.J.; Humphrey, J.W. Assessing structural diversity in managed forest. In Assessment of Biodiversity for Improved Forest Planning; Bachmann, P., Ed.; European Forest Institute Proceedings 18; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 331–342. [Google Scholar]
- Kneeshaw, D.D.; Leduc, A.; Drapeau, P.; Gauthier, S.; Pare, D.; Carignan, R.; Doucet, R.; Bouthillier, L.; Messier, C. Development of integrated ecological standards of sustainable forest management at an operational scale. For. Chron. 2000, 76, 481–493. [Google Scholar] [CrossRef] [Green Version]
- Zumeta, D.C.; Ellefson, P.V. Conserving the biological diversity of forests: Program and organizational experiences of state governments in the United States. Environ. Manag. 2000, 26, 393–402. [Google Scholar] [CrossRef]
- Larsson, S.; Danell, K. Science and the management of boreal forest biodiversity. Scand. J. For. Res. 2001, 3, 5–9. [Google Scholar] [CrossRef]
- Beatty, S.W. Influence of microtopography and canopy species on spatial patterns of forest understory plants. Ecology 1984, 65, 1406–1419. [Google Scholar] [CrossRef]
- Collins, B.S.; Dunne, K.P.; Pickett, S.T.A. Responses of forest herbs to canopy gaps. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 217–234. [Google Scholar]
- Siccama, T.G.; Bormann, F.H.; Likens, G.E. The Hubbard Brook Ecosystem Study: Productivity, nutrients, and phytosociology of the herbaceous layer. Ecol. Monogr. 1970, 40, 389–402. [Google Scholar] [CrossRef]
- Hutchinson, T.F.; Boerner, R.E.J.; Iverson, L.R.; Sutherland, S.; Kenedy-Sutherland, E. Landscape patterns of understory composition and richness across a moisture and nitrogen mineralization gradient in Ohio (U.S.A.) Quercus forests. Plant. Ecol. 1999, 144, 177–189. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Barnes, B.V. The use of ground flora to indicate edaphic factors in upland ecosystems of the McCormick Experimental Forest, Upper Michigan. Can. J. For. Res. 1982, 12, 661–672. [Google Scholar] [CrossRef]
- Meilleur, A.; Bouchard, A.; Bergeron, Y. The use of understory species as indicators of landform ecosystem type in heavily disturbed forest: An evaluation in the Haut-SaintLaurent, Quebec. Vegetatio 1992, 102, 13–32. [Google Scholar] [CrossRef]
- Laface, V.L.A.; Musarella, C.M.; Cano Ortiz, A.; Quinto Canas, R.; Cannavò, S.; Spampinato, G. Three New Alien Taxa for Europe and a Chorological Update on the Alien Vascular Flora of Calabria (Southern Italy). Plants 2020, 9, 1181. [Google Scholar] [CrossRef] [PubMed]
- Levizou, E.; Kyparissis, A. Ecophysiology of Mediterranean Plants; University of Ioannina, Department of Biological Applications and Technology: Ioannina, Greece, 2005; pp. 69–70. [Google Scholar]
- Rackham, O.; Moody, J. The Making of the Cretan Landscape; Manchester University Press: Manchester, UK, 1996. [Google Scholar]
- Klironomos, J. The actions of fungi in ecosystems. BioScience 2007, 57, 981–982. [Google Scholar] [CrossRef] [Green Version]
- Rossman, A.; Tulloss, R.; O’Dell, T.; Thorn, G. Protocols for an All Taxa Biodiversity Inventory of Fungi in a Costa Rican Conservation Area; Parkway Publishers: Blowing Rock, NC, USA, 1998. [Google Scholar]
- Dighton, J. Fungi in Ecosystem Processes; Dighton, J., Ed.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Heilmann-clausen, J.; Barron, E.S.; Boddy, L.; Dahlberg, A.; Griffith, G.W.; Nord, J.; Ovaskainen, O.; Perini, C.; Senn-irlet, B.; Halme, P. A fungal perspective on conservation biology. Conserv. Biol. 2014, 29, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoff, J.A.; Klopfenstein, N.B.; Tonn, J.R.; Mcdonald, G.I.; Zambino, P.J.; Rogers, J.D.; Peever, T.L.; Carris, L.M. Roles of Woody Root-Associated Fungi in Forest Ecosystem Processes: Recent Advances in Fungal Identification; General Technical Report; RMRS-RP-47; USDA Forest Service: Washington, DC, USA, 2004.
- Dahlberg, A.; Genney, D.R.; Heilmann-Clausen, J. Developing a comprehensive strategy for fungal conservation in Europe: Current status and future needs. Fungal Ecol. 2010, 3, 50–64. [Google Scholar] [CrossRef]
- Laganà, A.; Salerni, E.; Barluzzi, C.; De Dominics, V.; Perini, C. Fungi (macromycetes) in various types of Mediterranean forest ecosystems (Tuscany, Italy). Pol. Bot. J. 2002, 47, 143–165. [Google Scholar]
- Cockle, K.L.; Martin, K.; Robledo, G. Linking fungi, trees, and hole-using birds in a Neotropical tree-cavity network: Pathways of cavity production and implications for conservation. For. Ecol. Manag. 2012, 264, 210–219. [Google Scholar] [CrossRef]
- Zervakis, G.I.; Venturella, G. Adverse effects of human activities on the diversity of macrofungi in forest ecosystems. Bocconea 2007, 21, 77–84. [Google Scholar]
- Richard, F.; Moreau, P.-A.; Selosse, M.-A.; Gardes, M. Diversity and fruiting patterns of ectomycorrhizal and saprobic fungi in an old growth Mediterranean forest dominated by Quercus ilex. Can. J. Bot. 2004, 82, 1711–1729. [Google Scholar] [CrossRef] [Green Version]
- Zotti, M.; Pautasso, M. Macrofungi in Mediterranean Quercus ilex woodlands: Relations to vegetation structure, ecological gradients and higher-taxon approach. Czech. Mycol. 2013, 65, 193–218. [Google Scholar] [CrossRef]
- Rodríguez, M.H.; Oria-De-Rueda, J.A.; Martín-Pinto, P. Post-fire fungal succession in a Mediterranean ecosystem dominated by Cistus ladanifer L. Forest Ecol. Manag. 2013, 289, 48–57. [Google Scholar] [CrossRef]
- Richard, F.; Millot, S.; Gardes, M.; Selosse, M.-A. Diversity and specificity of ectomycorrhizal fungi retrieved from an old-growth Mediterranean forest dominated by Quercus ilex. New Phytol. 2005, 166, 1011–1023. [Google Scholar] [CrossRef]
- Lancellotti, E.; Schiaffino, A.; Franceschini, A.; Marras, F. Study of the ectomycorrhizal community in a cork oak stand of Sardinia. Micol. Ital. 2007, 36, 89–98. [Google Scholar]
- Lonsdale, D.; Pautasso, M.; Holdenrieder, O. Wood-decaying fungi in the forest: Conservation needs and management options. Eur. J. For. Res. 2008, 127, 1–22. [Google Scholar] [CrossRef]
- De Dominicis, V.; Barluzzi, C. Coenological research on macrofungi in evergreen oak woods in the hills near Siena (Italy). Vegetatio 1983, 54, 177–187. [Google Scholar] [CrossRef]
- Barluzzi, C.; De Dominicis, V.; Perini, C. Primi risultati di indagini micologiche sulle leccete delle colline del litorale grossetano. Micol. Ital. 1983, 12, 29–35. [Google Scholar]
- Perini, C.; Barluzzi, V.; De Dominicis, V. Mycocoenological research on evergreen Oak woods in the hills of the Maremma coastline (NW of Grosseto, Italy). Phytocoenologia 1989, 17, 289–306. [Google Scholar] [CrossRef]
- Signorello, P. Indagini micocenologiche sulle cenosi a Quercus ilex L. dell’Etna. Micol. Ital. 1996, 25, 74–80. [Google Scholar]
- Laganà, A.; Loppi, S.; De Dominicis, V. Relationship between environmental factors and the proportions of fungal trophic groups in forest ecosystems of the central Mediterranean area. For. Ecol. Manag. 1999, 124, 145–151. [Google Scholar] [CrossRef]
- Salerni, E.; Laganà, A.; De Dominicis, V. Mycocenological studies in deciduous Oak woods of central-southern Tuscany (Italy). Cryptogam. Mycol. 2001, 22, 35–55. [Google Scholar] [CrossRef]
- Santi, E.; Maccheroni, S.; Rocchini, D.; Bonini, I.; Brunialti, G.; Favilli, L.; Perini, C.; Pezzo, F.; Piazzino, S.; Rota, E.; et al. Simple to sample: Vascular plants as surrogate group in a nature reserve. J. Nat. Conserv. 2010, 18, 2–11. [Google Scholar] [CrossRef]
- Zotti, M.; Vizzini, A.; Traverso, M.; Boccardo, F.; Pavarino, M.; Mariotti, M.G. The macrofungi checklist of Liguria (Italy): The current status of surveys. Mycotaxon 2008, 105, 167–170. [Google Scholar]
- Ortega, A.; Lorite, J. Macrofungi diversity in cork-oak and holm-oak forests in Andalusia (southern Spain); an efficient parameter for establishing priorities for its evaluation and conservation. Cent. Eur. J. Biol. 2007, 2, 276–296. [Google Scholar] [CrossRef]
- Halme, P.; Heilmann-Clausen, J.; Rama, T.; Kosonen, T.; Kunttu, P. Monitoring fungal biodiversity—Towards an integrated approach. Fungal Ecol. 2012, 5, 750–758. [Google Scholar] [CrossRef]
- Rosa García, R.; Fraser, M.D.; Celaya, R.; Ferreira, L.M.M.; García, U.; Osoro, K. Grazing land management and biodiversity in the Atlantic European heathlands: A review. Agroforest. Syst. 2013, 87, 19–43. [Google Scholar] [CrossRef]
- Nauta, M.M.; Jalink, L.M. Grassland in the coastal dunes: The effect of nature management on the mycota. In Fungal Conservation: Issues and Solutions; Moore, D., Nauta, M.M., Evans, S.E., Rotheroe, M., Eds.; British Mycological Society, Cambridge University Press: Cambridge, UK, 2001; pp. 136–143. [Google Scholar]
- Raatikainen, K.M.; Heikkinen, R.K.; Pykala, J. Impacts of local and regional factors on vegetation of boreal semi-natural grasslands. Plant Ecol. 2007, 189, 155–173. [Google Scholar] [CrossRef]
- Pykala, J. Maintaining Plant Species Richness by Cattle Grazing: Mesic Semi-Natural Grasslands as Focal Habitats; Publications in Botany from the University of Helsinki: Helsinki, Finland, 2007; pp. 36–37. [Google Scholar]
- Jakobsson, S. Perinteiset kulttuuribiotoopit. In Suomen Helttasienten ja Tattien Ekologia, Levinneisyys ja Uhanalaisuus; Salo, P., Niemelä, T., Nummela-Salo, U., Ohenoja, E., Eds.; Suomen Ympäristökeskus: Helsinki, Finland, 2005. [Google Scholar]
- Olff, H.; Ritchie, M.E. Effects of herbivores on grassland plant diversity. Trends Ecol. Evol. 1998, 13, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Mwendera, E.J.; Saleem, M.A.M.; Woldu, Z. Vegetation response to cattle grazing in the Ethiopian highlands. Agric. Ecosyst. Environ. 1997, 64, 43–51. [Google Scholar] [CrossRef]
- Pykala, J. Effects of restoration with cattle grazing on plant species composition and richness of semi-natural grasslands. Biodivers. Conserv. 2003, 12, 2211–2226. [Google Scholar] [CrossRef]
- Lindborg, R.; Eriksson, O. Historical landscape connectivity affects present plant species diversity. Ecology 2004, 85, 1840–1845. [Google Scholar] [CrossRef]
- Poyry, J.; Luoto, M.; Paukkunen, J.; Pykala, J.; Raatikainen, K.; Kuussaari, M. Different responses of plants and herbivore insects to a gradient of vegetation height: An indicator of the vertebrate grazing intensity and successional age. Oikos 2006, 115, 401–412. [Google Scholar] [CrossRef]
- Chiarucci, A.; D’Auria, F.; De Dominicis, V.; Laganá, A.; Perini, C.; Salerni, E. Using vascular plants as a surrogate taxon to maximize fungal species richness in reserve design. Conserv. Biol. 2005, 19, 1644–1652. [Google Scholar] [CrossRef]
- Brown, N.; Bhagwat, S.; Watkinson, S. Macrofungal diversity in fragmented and disturbed forests of the Western Ghats of India. J. Appl. Ecol. 2006, 43, 11–17. [Google Scholar] [CrossRef]
- McMullan-Fisher, S.J.M.; Kirkpatrick, J.B.; May, T.W.; Pharo, E.J. Surrogates for macrofungi and mosses in reservation planning. Conserv. Biol. 2010, 24, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, K.; Morschhauser, T.; Pál-Fám, F.; Botta-Dukát, Z. Exploring the relationship between macrofungi diversity, abundance, and vascular plant diversity in semi-natural and managed forests in north-east Hungary. Ecol. Res. 2013, 28, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Schmit, J.P.; Mueller, G.M.; Leacock, P.R.; Mata, J.L.; Wu, Q.; Huang, Y. Assessment of tree species richness as a surrogate for macrofungal species richness. Biol. Conserv. 2005, 121, 99–110. [Google Scholar] [CrossRef]
- Arnolds, E. Decline of ectomycorrhizal fungi in Europe. Agric. Ecosyst. Environ. 1991, 35, 209–244. [Google Scholar] [CrossRef]
- Gabel, A.C.; Gabel, M.L. Comparison of diversity of macrofungi and vascular plants at seven sites in the Black Hills of South Dakota. Am. Midl. Nat. 2007, 157, 258–296. [Google Scholar] [CrossRef]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, Present, and Future Perspectives of Environmental DNA (EDNA) Metabarcoding: A Systematic Review in Methods, Monitoring, and Applications of Global EDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Bohmann, K.; Evans, A.; Gilbert, M.T.P.; Carvalho, G.R.; Creer, S.; Knapp, M.; Douglas, W.Y.; De Bruyn, M. Environmental DNA for Wildlife Biology and Biodiversity Monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; De Vere, N. Environmental DNA Metabarcoding: Transforming How We Survey Animal and Plant Communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Van der Heyde, M.; Bunce, M.; Wardell-Johnson, G.; Fernandes, K.; White, N.E.; Nevill, P. Testing Multiple Substrates for Terrestrial Biodiversity Monitoring Using Environmental DNA Metabarcoding. Mol. Ecol. Resour. 2020, 20, 732–745. [Google Scholar] [CrossRef]
- Ekschmitt, K.; Bakonyi, G.; Bongers, M.; Bongers, T.; Boström, S.; Dogan, H.; Harrison, A.; Nagy, P.; O’Donnell, A.G.; Papatheodorou, E.M. Nematode Community Structure as Indicator of Soil Functioning in European Grassland Soils. Eur. J. Soil Biol. 2001, 37, 263–268. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Young, I.M.; Caul, S. Nematode and Protozoan Population Dynamics on Decomposing Barley Leaves Incubated at Different Soil Matric Potentials. Pedobiologia 1995, 39, 454–461. [Google Scholar]
- Gupta, V.V.; Yeates, G.W. Soil microfauna as bioindicators of soil health. Ιn Biological Indicators of Soil Health; Pankhurst, C., Doube, B.M., Gupta, V.V.S.R., Eds.; CAB International: New York, NY, USA, 1997; pp. 201–233. [Google Scholar]
- Elliot, E.T. Rationale for Developing Bioindicators of Soil Health. Biological indicators of soil health. Biol. Indic. Soil Health 1997, 49–78. Available online: https://agris.fao.org/agris-search/search.do;jsessionid=29564E0FFAD720E56AE51D5AE85C62E1?request_locale=zh_CN&recordID=US1997059136&query=&sourceQuery=&sortField=&sortOrder=&agrovocString=&advQuery=¢erString=&enableField (accessed on 8 August 2021).
- Henkin, Z.; Landau, S.Y.; Ungar, E.D.; Perevolotsky, A.; Yehuda, Y.; Sternberg, M. Effect of Timing and Intensity of Grazing on the Herbage Quality of a Mediterranean Rangeland. J. Anim. Feed Sci. 2007, 16, 318–322. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.H.; Mcsorley, R.; Bohlen, P.; Gathumbi, S.M. Cattle Grazing Increases Microbial Biomass and Alters Soil Nematode Communities in Subtropical Pastures. Soil Biol. Biochem. 2006, 38, 1956–1965. [Google Scholar] [CrossRef]
- Li, Q.; Mayzlish, E.; Shamir, I.; Pen-Mouratov, S.; Sternberg, M.; Steinberger, Y. Impact of Grazing on Soil Biota in a Mediterranean Grassland. Land Degrad. Dev. 2005, 16, 581–592. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Leemans, D.K.; Cook, R.; Hobbs, P.J. Seasonality of the Soil Biota of Grazed and Ungrazed Hill Grasslands. Soil Biol. Biochem. 1997, 29, 1285–1294. [Google Scholar] [CrossRef]
- Hu, J.; Wu, J.; Ma, M.; Nielsen, U.N.; Wang, J.; Du, G. Nematode Communities Response to Long-Term Grazing Disturbance on Tibetan Plateau. Eur. J. Soil Biol. 2015, 69, 24–32. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W.K. A Generalized Model of the Effects of Grazing by Large Herbivores on Grassland Community Structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Talamucci, P.; Chaulet, C. Contraintes et evolution des ressources fourrageres dans le basin mediterranees. In Proceedings of the XVI International Grassland Congress, Nice, France, 4–11 October 1989; pp. 1731–1740. [Google Scholar]
- Le Houerou, H.N. Grazing lands of the Mediterranean basin. In Natural Grassland; Coupland, R.T., Ed.; Ecosystems of the World; Elsevier: Amsterdam, The Netherlands, 1993; Volume 8, pp. 171–196. [Google Scholar]
- Papanastasis, V.P.; Mansat, P. Grasslands and related forage resources in Mediterranean areas. In Grasslands and Land Use Systems; Parente, G., Frame, J., Orsi, S., Eds.; Grassland Science in Europe, ERSA: Gorizia, Italy, 1996; Volume 1, pp. 47–57. [Google Scholar]
- Bartolomé, J.; Miró, J.; Panadès, X.; Broncano, M.J.; Plaixats, J.; Rigau, T.; Milán, M.J.; Baraza, E. Preference by Donkeys and Goats among Five Mediterranean Forest Species: Implications for Reducing Fire Hazard. Animals 2020, 10, 1302. [Google Scholar] [CrossRef]
- Papanastasis, V.P.; Yiakoulaki, M.D.; Decandia, M.; Dini-Papanastasi, O. Integrating Woody Species into Livestock Feeding in the Mediterranean Areas of Europe. Anim. Feed Sci. Technol. 2008, 140, 1–17. [Google Scholar] [CrossRef]
- Wilson, A.D. A review of browse in the nutrution of grazing animals. J. Range Manag. 1969, 22, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Holechek, J.L. Comparative contribution of grasses, forbs and shrubs to the nutrition of range ungulates. Rangelands 1984, 6, 261–263. [Google Scholar]
- Ramirez, R.G.; Loyo, A.; Mora, R.; Sanchez, E.M.; Chaire, A. Forage intake and nutrition of range goats in a shrubland in northeastern Mexico. J. Anim. Sci. 1991, 69, 879–885. [Google Scholar] [CrossRef]
- Nastis, A.S. Nutritive value of fodder shrubs. In Fodder Trees and Shrubs in the Mediterranean Production Systems: Objectives and Expected Results of the EC Research Contract; Papanastasis, V.P., Ed.; Agriculture, Agrimed Research Programme, Commission of the European Communities Report EUR 14459 EN; Commission of the European Communities: Luxembourg, 1993; pp. 75–81. [Google Scholar]
- Papachristou, T.G. Intake, digestibility and nutrient utilization of oriental hornbeam and manna ash browse by goats and sheep. Small Rumin. Res. 1997, 23, 91–98. [Google Scholar] [CrossRef]
- Papachristou, T.G. Foraging behaviour of goats and sheep on Mediterranean kermes oak shrublands. Small Rumin. Res. 1997, 24, 85–93. [Google Scholar] [CrossRef]
- Papachristou, T.G.; Platis, P.D.; Papanastasis, V.P.; Tsiouvaras, C.N. Use of deciduous woody species as a diet supplement for goats grazing Mediterranean shrublands during the dry season. Anim. Feed Sci. Technol. 1999, 80, 267–279. [Google Scholar] [CrossRef]
- Papachristou, T.G.; Nastis, A.S. Diets of goats grazing oak shrublands of varying cover in northern Greece. J. Range Manag. 1993, 6, 220–226. [Google Scholar] [CrossRef]
- Dumont, B.; Meuret, M.; Prudhon, M. Direct observation of biting for studying grazing behavior of goats and llamas on garrigue rangelands. Small Rumin. Res. 1995, 16, 27–35. [Google Scholar] [CrossRef]
- Espejo-Diaz, M. Evaluation and improvement of efficacy of animal production systems using natural resources by grazing ruminants in Mediterranean areas. In Optimal Exploitation of Marginal Mediterranean Areas by Extensive Ruminant Production Systems, Proceedings of the International Symposium 18–20 June 1994; Zervas, N.P., Hatziminaoglou, J., Eds.; EAAP Publication: Thessaloniki, Greece, 1996; pp. 105–112. [Google Scholar]
- Meuret, M. Food harvesting by small ruminants foraging on rangeland and woodland undergrowth (in French with English abstract). INRA Prod. Anim. 1997, 10, 391–401. [Google Scholar] [CrossRef]
- Dini, O. Genetic potential of Robinia pseudoacacia L. In Fodder Trees and Shrubs in the Mediterranean Production Systems: Objectives and Expected Results of the EC Research Contract; Papanastasis, V.P., Ed.; Agriculture, Agrimed Research Programme, Commission of the European Communities EUR 11459 EN; Commission of the European Communities: Luxembourg, 1993; pp. 153–159. [Google Scholar]
- Platis, P.D.; Papanastasis, V.P. Productivity of deciduous fodder trees and shrubs in relation to the year of cutting. In Management of Mediterranean Shrublands and Related Forage Resources; REUR Technical Series 28; FAO: Rome, Italy, 1993; pp. 134–136. [Google Scholar]
- Papachristou, T.G.; Papanastasis, V.P. Forage value of Mediterranean deciduous woody fodder species and its implication to management of silvo–pastoral systems for goats. Agrofor. Syst. 1994, 27, 269–282. [Google Scholar] [CrossRef]
- Ainalis, A.B.; Tsiouvaras, C.N.; Noitsakis, B.; Papanastasis, V.P. Growth dynamics of some woody fodder plants in relation to spacing and grazing. In Ecological Basis of Livestock Grazing in Mediterranean Ecosystems; Papanastasis, V.P., Ed.; International Workshop, EAAP Publication: Thessaloniki, Greece, 1998; pp. 80–85. [Google Scholar]
- Getabalew, M.; Alemneh, T. Factors Affecting the Productivity of Rangelands. J. Plant Sci. Agric. Res. 2019, 3, 19. [Google Scholar]
- Papanastasis, V.P. Review of papers on woody forage plants. Herba 1993, 6, 28–33. [Google Scholar]
- Kebaili, A.; Papanastasis, V.P. Comparison of seasonally available biomass in several woody species of mountainous Mediterranean rangelands. In Animal Production and Natural Resources Utilisation in the Mediterranean Mountain Areas; Georgoudis, A., Rosati, A., Mosconi, C., Eds.; EAAP Scientific Series No. 115; Wageningen Academic Publishers: Wageningen, The Netherlands, 2005; pp. 124–128. [Google Scholar]
- Papachristou, T.G.; Dziba, L.E.; Provenza, F.D. Foraging ecology of goats and sheep on wooded rangelands. Small Rumin. Res. 2005, 59, 141–156. [Google Scholar] [CrossRef]
- Provenza, F.D. Acquired aversions as the basis for varied diets of ruminants foraging on rangelands. J. Anim. Sci. 1996, 74, 2010–2020. [Google Scholar] [CrossRef]
- Sales-Baptista, E.; Ferraz-de-Oliveira, M.I. Grazing in Silvopastoral Systems: Multiple Solutions for Diversified Benefits. Agrofor. Syst. 2021, 95, 1–6. [Google Scholar] [CrossRef]
- Biró, M.; Molnár, Z.; Öllerer, K.; Lengyel, A.; Ulicsni, V.; Szabados, K.; Kiš, A.; Perić, R.; Demeter, L.; Babai, D. Conservation and Herding Co-Benefit from Traditional Extensive Wetland Grazing. Agric. Ecosyst. Environ. 2020, 300, 106983. [Google Scholar] [CrossRef]
- Öllerer, K.; Varga, A.; Kirby, K.; Demeter, L.; Biró, M.; Bölöni, J.; Molnár, Z. Beyond the obvious impact of domestic livestock grazing on temperate forest vegetation—A global review. Biol. Conserv. 2019, 237, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.; Coppa, M.; Verdier-Metz, I.; Montel, M.C.; Joy, M.; Casasús, I.; Blanco, M. The contribution of mountain pastures to the link to terroir in dairy and meat products. In Mountain Pastures and Livestock Farming Facing Uncertainty: Environmental, Technical and Socio-Economic Challenges; Casasús, I., Lombardi, G., Eds.; CIHEAM: Zaragoza, Spain, 2016; pp. 105–115. [Google Scholar]
- Santini, F.; Guri, F.; Gomez, Y.; Paloma, S. Labelling of Agricultural and Food Products of Mountain Farming; Joint Research Center Scientific and Policy Reports; Citeseer: Luxembourg, 2013; Volume 154, Available online: http://ec.europa.eu/agriculture/external-studies/2013/mountain-farming/fulltext_en.pdf (accessed on 20 July 2021).
- Thénard, V.; Choisis, J.P.; Pagès, Y. Towards sustainable dairy sheep farms based on self-sufficiency: Patterns and environmental issues. Mountain pastures and livestock farming. Options Mediterr. 2016, 116, 81–85. [Google Scholar]
- Joy, M.; Sanz, A.; Ripoll, G.; Panea, B.; Ripoll-Bosch, R.; Blasco, I.; Alvarez-Rodriguez, J. Does forage type (grazing vs. hay) fed to ewes before and after lambing affect suckling lambs performance, meat quality and consumer purchase intention? Small Rumin. Res. 2012, 104, 1–9. [Google Scholar] [CrossRef]
- Beck, M.R.; Gregorini, P. Animal Design Through Functional Dietary Diversity for Future Productive Landscapes. Front. Sustain. Food Syst. 2021, 5, 138. [Google Scholar] [CrossRef]
- Renna, M.; Ravetto Enri, S.; Probo, M.; Lussiana, C.; Malfatto, V.; Battaglini, L.M.; Lonati, M.; Lombardi, G. Alpine grasslands: Relations among botanical and chemical variables affecting animal product quality. In Proceedings of the 19th Meeting of the FAO CIHEAM, Mountain Pastures Network–Mountain Pastures and Livestock Farming Facing Uncertainty: Environmental, Technical and Socio-Economic Challenges; Casasús, I., Lombardi, G., Eds.; CIHEAM: Zaragoza, Spain, 2016; pp. 14–16. [Google Scholar]
- Prache, S.; Theriez, M. Traceability of lamb production systems: Carotenoids in plasma and adipose tissue. Anim. Sci. 1999, 69, 29–36. [Google Scholar] [CrossRef]
- Joy, M.; Ripoll, G.; Molino, F.; Dervishi, E.; Alvarez-Rodriguez, J. Influence of the type of forage supplied to ewes in pre- and post-partum periods on the meat fatty acids of suckling lambs. Meat Sci. 2012, 90, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Rodríguez, J.; Sanz, A.; Delfa, R.; Revilla, R.; Joy, M. Performance and grazing behavior of Churra Tensina sheep stocked under different management systems during lactation on Spanish mountain pastures. Livest. Sci. 2007, 107, 152–161. [Google Scholar] [CrossRef]
- Prache, S.; Cornu, A.; Berdague, J.L.; Priolo, A. Traceability of animal feeding diet in the meat and milk of small ruminants. Small Rumin. Res. 2005, 59, 157–168. [Google Scholar]
- Priolo, A.; Micol, D.; Agabriel, J.; Prache, S.; Dransfield, E. Effect of grass or concentrate feeding systems on lamb carcass and meat quality. Meat Sci. 2002, 62, 179–185. [Google Scholar] [CrossRef]
- Valvo, M.A.; Lanza, M.; Bella, M.; Fasone, V.; Scerra, M.; Biondi, L.; Priolo, A. Effect of ewe feeding system (grass v. concentrate) on intramuscular fatty acids of lambs raised exclusively on maternal milk. Anim. Sci. 2005, 81, 431–436. [Google Scholar] [CrossRef]
- Atti, N.; Rouissi, H.; Othmane, M.H. Milk production, milk fatty acid composition and conjugated linoleic acid (CLA) content in dairy ewes raised on feddlot or grazing pasture. Livest. Sci. 2006, 104, 121–127. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.J.; Lee, M.R.F.; Scollan, N.D. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems, Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Mariaca, R.G.; Berger, T.F.H.; Gauch, R.; Imhof, M.I.; Jeangros Band Bosset, J.O. Occurrence of volatile mono- and sesquiterpenoids in highland and lowland plant species as possible precursors for flavor compounds in milk and dairy products. J. Agric. Food Chem 1997, 45, 4423–4434. [Google Scholar] [CrossRef]
- Buchin, S.; Martin, B.; Dupont, D.; Bornard, A.; Achilleos, C. Influence of the composition of Alpine highland pasture on the chemical, rheological and sensory properties of cheese. J. Dairy Res. 1999, 66, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Bendall, J.G. Aroma compounds of fresh milk from New Zealand cows fed different diets. J. Agric. Food Chem. 2001, 49, 4825–4832. [Google Scholar] [CrossRef]
- Viallon, C.; Martin, B.; Verdier-Metz, I.; Pradel, P.; Garel, J.P.; Coulon, J.B.; Berdagué, J.L. Transfer of monoterpenes and sesquiterpenes from forages into milk fat. Lait 2000, 80, 12–16. [Google Scholar]
- De Noni, I.; Battelli, G. Terpenes and fatty acid profiles of milk fat and Bitto cheese as affected by transhumance of cows on different mountain pastures. Food Chem. 2008, 109, 299–309. [Google Scholar] [CrossRef]
- Tornambé, G.; Cornu, A.; Verdier-Metz, I.; Pradel, P.; Kondojoyan, N.; Figueredo, G.; Hulin, S.; Martin, B. Addition of pasture plant essential oil in milk: Influence on chemical and sensory properties of milk and cheese. J. Dairy Sci. 2008, 91, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Besle, J.M.; Viala, D.; Martin, B.; Pradel, P.; Meunier, B.; Berdagué, J.L.; Fraisse, D.; Lamaison, J.L.; Coulon, J.B. Ultraviolet-absorbing compounds in milk are related to the forage polyphenols. J. Dairy Sci. 2010, 93, 2846–2856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, B.; Verdier-Metz, I.; Buchin, S.; Hurtaud, C.; Coulon, J.B. How does the nature of forages and pastures diversity influence the sensory quality of dairy livestock products? Anim. Sci. 2005, 81, 205–212. [Google Scholar] [CrossRef]
- Coppa, M.; Ferlay, A.; Monsallier, F.; Verdier-Metz, I.; Pradel, P.; Didienne, R.; Farruggia, A.; Montel, M.C.; Martin, B. Milk fatty acid composition and cheese texture and appearance from cows fed hay or different grazing systems on upland pastures. J. Dairy Sci. 2011, 94, 1132–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, B.; Lherm, M.; Béranger, C. Evolutions et perspectives de l’élevage des ruminants dans les montagnes françaises. INRA Prod. Anim. 2014, 27, 5–16. [Google Scholar] [CrossRef]
- Coppa, M.; Martin, B.; Pradel, P.; Leotta, B.; Priolo, A.; Vasta, V. Effect of a hay-based diet or different upland grazing systems on milk volatile compounds. J. Agric. Food Chem. 2011, 59, 4947–4954. [Google Scholar] [CrossRef]
- Mollard, A.; Pecqueur, B. De l’hypothèse au modèle du panier de biens et de services. Histoire succincte d’une recherché. Économie Rurale 2007, 300, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Dervillé, M.; Allaire, G. Quelles adaptations possibles à la suppression des quotas pour les filières laitières de montagne? Une approche en termes de régime de concurrence. INRA Prod. Anim. 2014, 27, 17–30. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Topalidou, E.; Solomou, A.D.; Santos, S.S.; Krystallidou, E.; Kakara, S.; Mantzanas, K. Dynamic Role and Importance of Multi-Kingdom Communities in Mediterranean Wood-Pastures. Sustainability 2021, 13, 10179. https://doi.org/10.3390/su131810179
Topalidou E, Solomou AD, Santos SS, Krystallidou E, Kakara S, Mantzanas K. Dynamic Role and Importance of Multi-Kingdom Communities in Mediterranean Wood-Pastures. Sustainability. 2021; 13(18):10179. https://doi.org/10.3390/su131810179
Chicago/Turabian StyleTopalidou, Eleni, Alexandra D. Solomou, Susana S. Santos, Evdokia Krystallidou, Styliani Kakara, and Konstantinos Mantzanas. 2021. "Dynamic Role and Importance of Multi-Kingdom Communities in Mediterranean Wood-Pastures" Sustainability 13, no. 18: 10179. https://doi.org/10.3390/su131810179
APA StyleTopalidou, E., Solomou, A. D., Santos, S. S., Krystallidou, E., Kakara, S., & Mantzanas, K. (2021). Dynamic Role and Importance of Multi-Kingdom Communities in Mediterranean Wood-Pastures. Sustainability, 13(18), 10179. https://doi.org/10.3390/su131810179