
Vegetative Propagation and ISSR-Based Genetic Identification of Genotypes of Ilex aquifolium ‘Agrifoglio Commune’

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Rooting Procedure
2.2. Effect of Season and Auxin Treatment on Rooting of Cuttings
2.3. Effect of Type of Cutting and Auxin Treatment on Rooting of Cuttings
2.4. Data Recording and Statistical Analysis
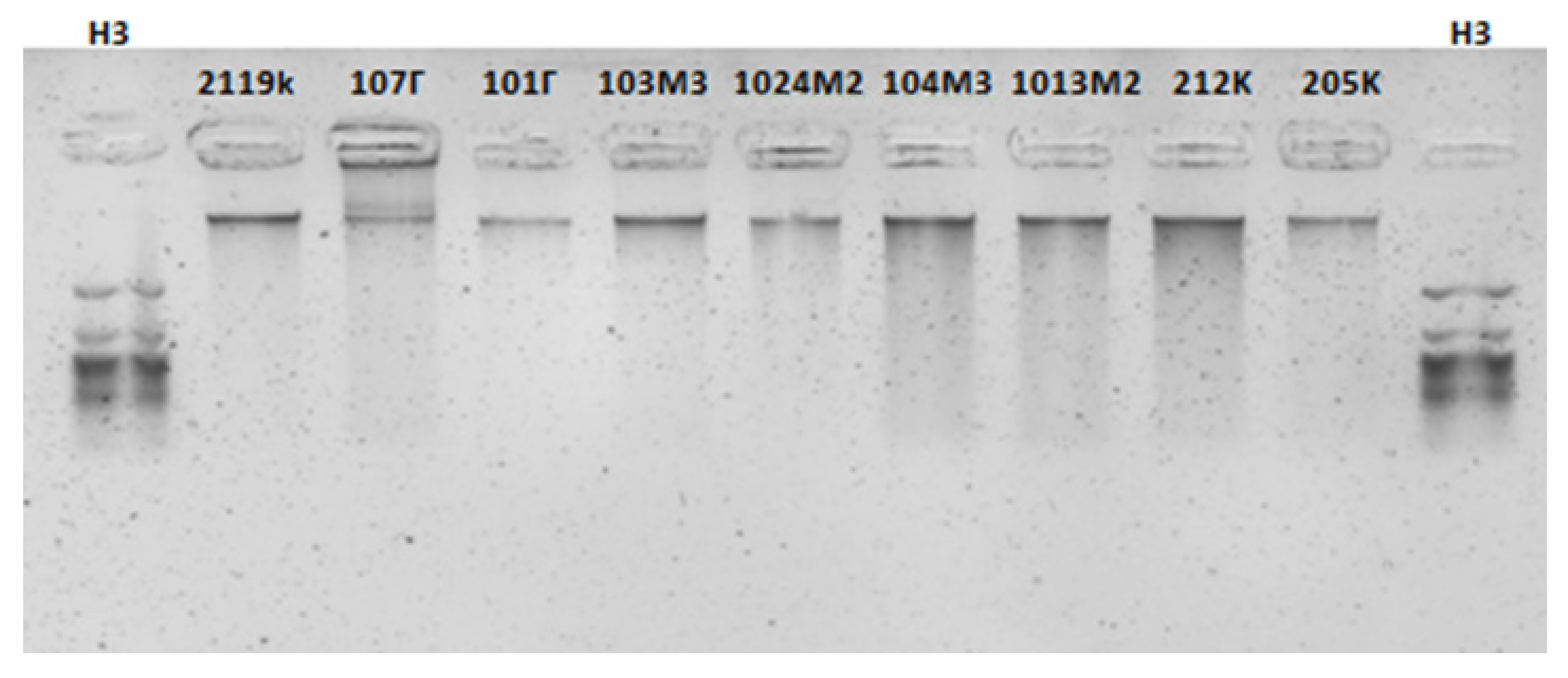
2.5. Genomic DNA Isolation
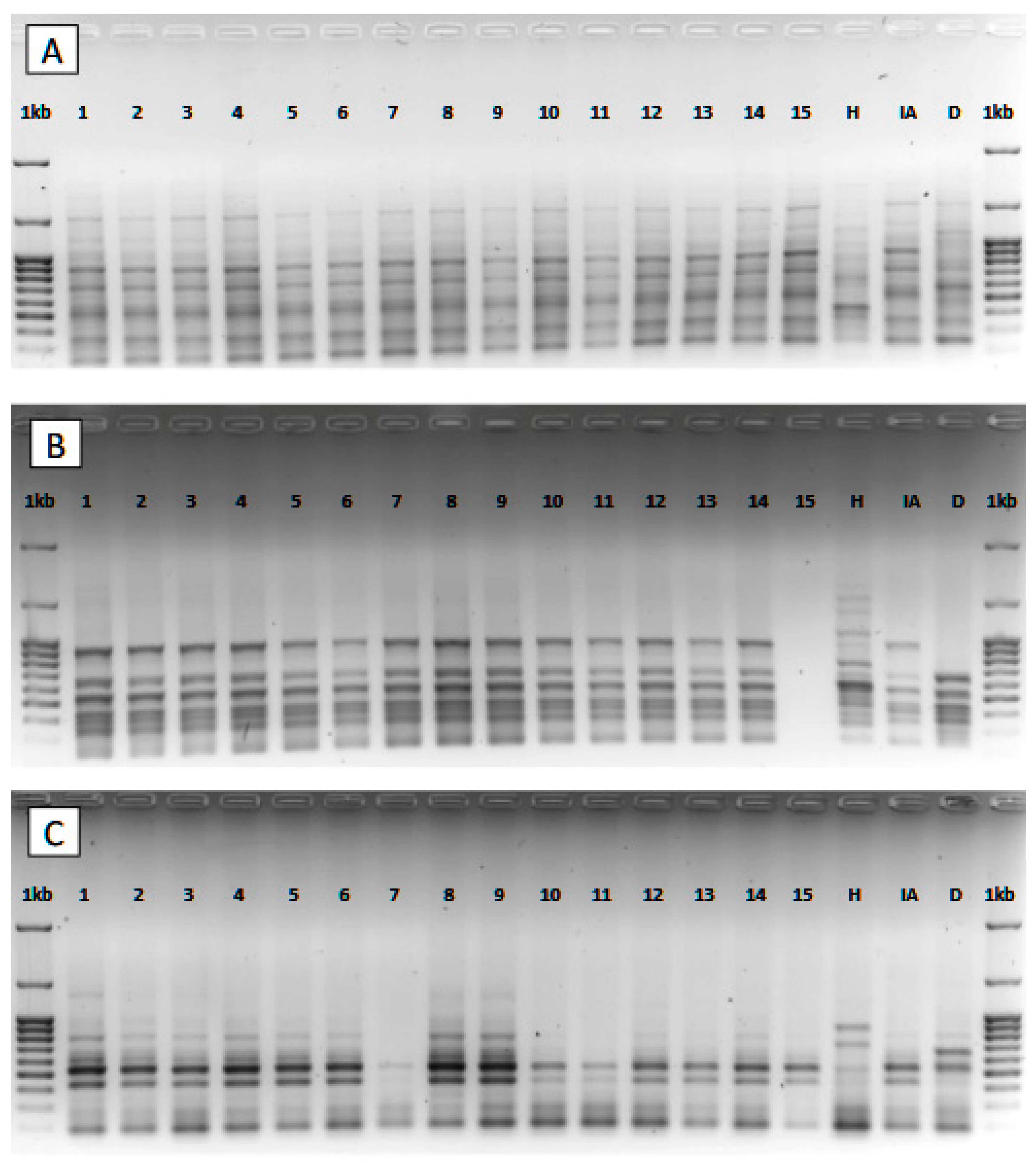
2.6. PCR Amplification
- I.
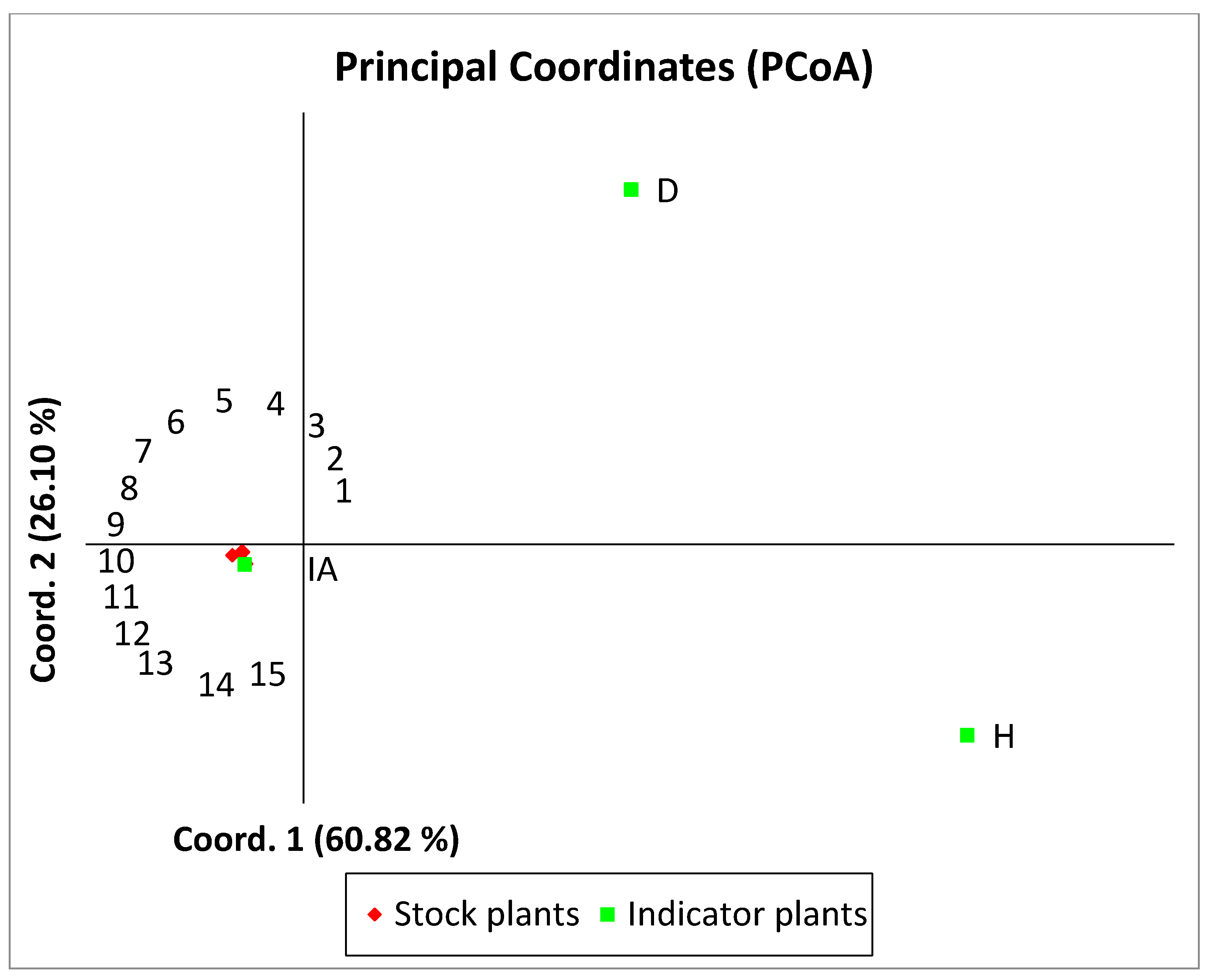
- The principal coordinates analysis (PCoA) with GenAlEx 6.501 program [26]. The PCoA input data type was tri distance matrix, and the PCoA method was covariance-standardized.
- II.
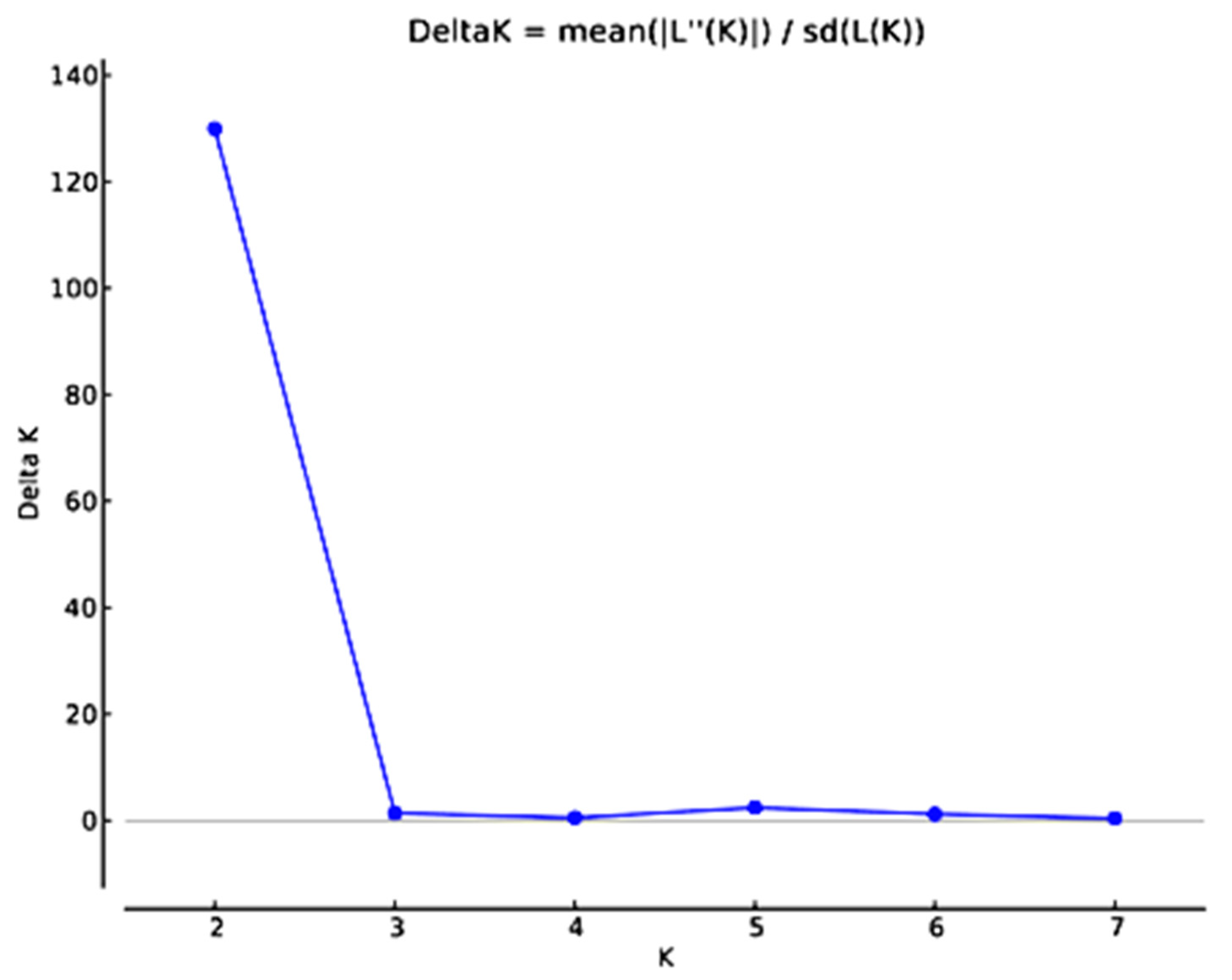
- A STRUCTURE analysis, where similarity estimates were analyzed using the Structure 2.3.4 software package [27]. This software places cultivars in K clusters that have distinct marker frequencies, where K is chosen a priori and can be varied across different runs (K = 1–8). Samples can have memberships in several clusters, with membership coefficients equaling 1 across clusters. A nonadmixture ancestry model was used and binary frequencies were correlated, with a burn-in length of 30,000 followed by 100,000 runs at each K [28,29]. K was determined using the ad hoc statistic ΔK as in Evanno et al. [30] with STRUCTURE HARVESTER program [31].
- III.
- An unweighted paired group method of cluster analysis (UPGMA) dendrogram with MEGA 4 v.4.1 software [32].
3. Results and Discussion
3.1. Effect of Season and Auxin Treatment on Rooting of Cuttings
3.2. Effect of Type of Cutting and Auxin Treatment on Rooting of Cuttings
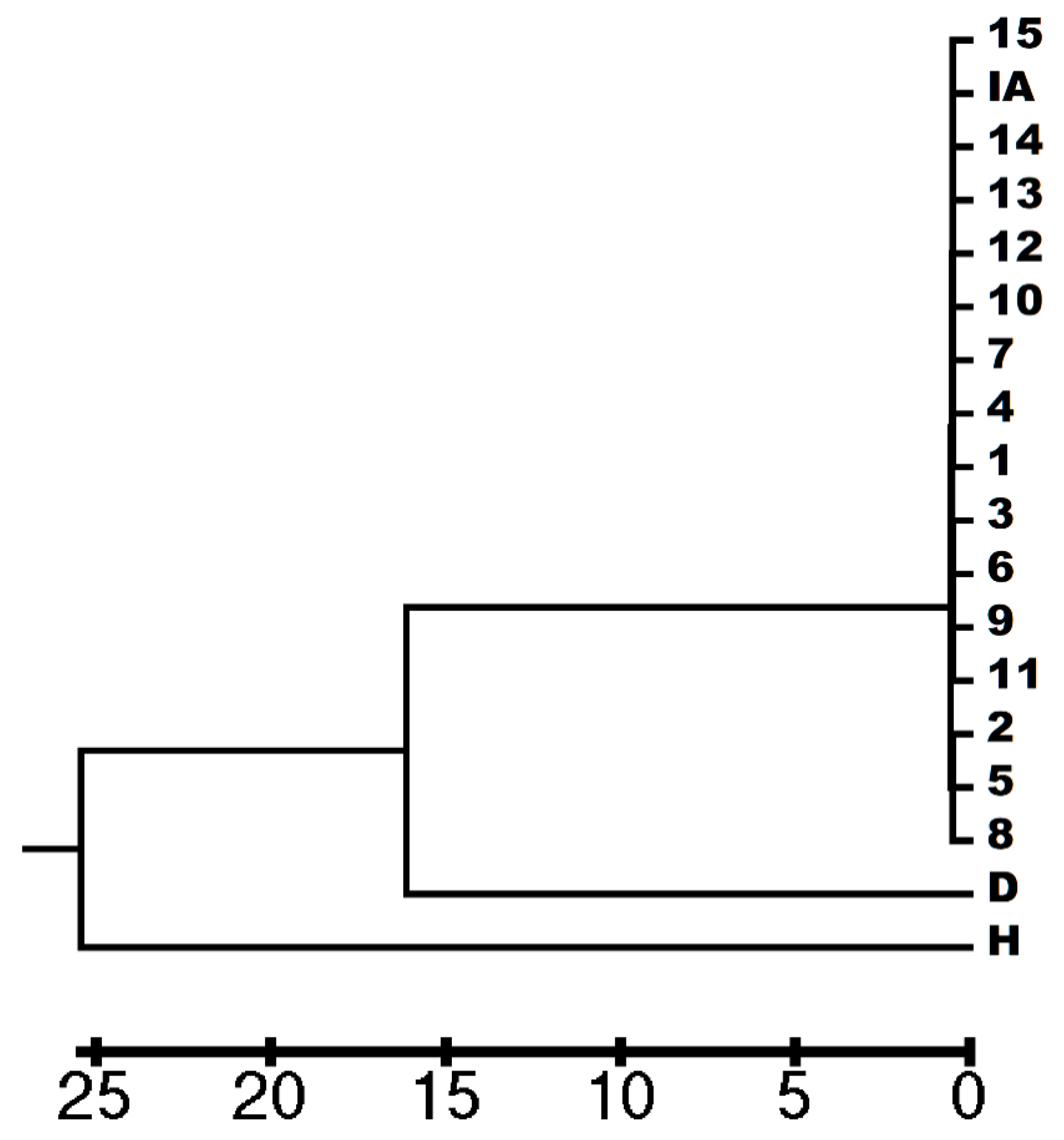
3.3. Genetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerrero-Hue, N.; Caudullo, G.; de Rigo, D. Ilex aquifolium in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office EU: Luxembourg, 2016; p. 102. [Google Scholar]
- Niemiera, A.X. Evergreen Hollies (Ilex spp.); Virginia Cooperative Extension; Virginia Tech and Virginia State University: Blacksburg, VA, USA, 2018; pp. 1482–1487. [Google Scholar]
- Peterken, G.F.; Lloyd, P.S. Ilex aquifolium L. J. Ecology 1967, 55, 841–858. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T., Jr.; Geneve, R.L. Hartmann and Kester’s Plant Propagation: Principles and Practices, 8th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2011; p. 915. [Google Scholar]
- Dirr, M.A.; Heuser, C.W., Jr. The Reference Manual of Woody Plant Propagation: From Seed to Tissue Culture; Varsity Press: Athens, GA, USA, 1987; p. 239. [Google Scholar]
- French, C.L.; Liu, W.C. Seasonal variation in the effects of CO2, mist and supplementary lightening from high pressure sodium lamps on rooting of English holly cuttings. HortScience 1984, 19, 519–521. [Google Scholar]
- Rifaki, N.; Economou, A.; Scaltsoyiannes, A. Factors affecting the rooting of Ilex aquifolium L. cuttings. Prop. Ornam. Plants 2001, 1, 31–35. [Google Scholar]
- Tsaktsira, M.; Alevropoulos, A.; Tsoulpha, P.; Scaltsoyiannes, V.; Scaltsoyiannes, A.; Iliev, I. Inter- and intra-genetic variation on rooting ability of Ilex aquifolium L. varieties and cultivars. Prop. Ornam. Plants 2018, 18, 131–138. [Google Scholar]
- Blythe, E.K.; Sibley, J.L.; Tilt, K.M.; Ruter, J.M. Auxin application to stem cuttings of selected woody landscape plants by incorporation into a stabilized organic rooting substrate. J. Environ. Hort. 2004, 22, 63–70. [Google Scholar] [CrossRef]
- Blythe, E.; Sibley, J. Sodium cellulose glycolate as a thickening agent for liquid auxin formulations can enhance rooting of stem cuttings. J. Environ. Hort. 2007, 25, 126–130. [Google Scholar] [CrossRef]
- Lambert, C.R.; Blazich, F.A.; LeBude, A.V. Propagation of Ilex vomitoria ‘Dare County’ by stem cuttings. J. Environ. Hort. 2012, 30, 55–57. [Google Scholar] [CrossRef]
- Blythe, E.; Sibley, J. Winter stem cutting propagation of ‘Dwarf Burford’ holly without use of a conventional auxin treatment. HortTechnology 2009, 19, 130–132. [Google Scholar] [CrossRef] [Green Version]
- Blythe, E.K.; Sibley, J.L. Winter stem cutting propagation of Heller’s Japanese holly with and without the use of a conventional auxin treatment. HortTechnology 2012, 22, 771–773. [Google Scholar] [CrossRef] [Green Version]
- Maynard, B.K. Evaluating the role of pH in the rooting of cuttings. Comb. Proc. Int. Plant Prop. Soc. 2000, 50, 268–273. [Google Scholar]
- Jabbarzadeh, Z.; Khosh-Khui, M.; Salehi, H.; Saberivand, A. Inter simple sequence repeat (ISSR) markers as reproducible and specific tools for genetic diversity analysis of rose species. Afr. J. Biotechol. 2010, 9, 6091–6095. [Google Scholar]
- Ng, W.L.; Tan, S.G. Inter-simple sequence repeat (ISSR) markers: Are we doing it right. ASM Sci. J. 2015, 9, 30–39. [Google Scholar]
- Pakseresht, F.; Talebi, R.; Karami, E. Comparative assessment of ISSR, DAMD and SCoT markers for evaluation of genetic diversity and conservation of landrace chickpea (Cicer arietinum L.) genotypes collected from north-west of Iran. Physiol. Mol. Biol. Plants 2013, 19, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Cuesta, C.; Ordas, R.J.; Rodriguez, A.; Fernandez, B. PCR-based molecular markers for assessment of somaclonal variation in Pinus picea clones micro-propagated in vitro. Biol. Plant. 2010, 54, 435–442. [Google Scholar] [CrossRef]
- Alatar, A.; Faisal, M. Encapsulation of Rauvolfia tetraphylla microshoots as artificial seeds and evaluation of genetic fidelity using RADP and ISSR markers. J. Medic. Plants Res. 2012, 6, 1367–1374. [Google Scholar]
- Faisal, M.; Alatar, A.A.; Ahmad, N.; Anis, M.; Hegazy, A.K. Assessment of genetic fidelity in Rauvolfia serpentina plantlets grown from synthetic (encapsulated) seeds following in vitro storage at 4 °C. Molecules 2012, 17, 5050–5061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrotra, S.; Khwaja, O.; Kukreja, A.K.; Rahman, L. ISSR and RAPD based evaluation of genetic stability of encapsulated micro shoots of Glycyrrhiza glabra following 6 months of storage. Mol. Biotechnol. 2012, 52, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Hatzilazarou, S.; Kostas, S.; Nendou, T.; Economou, A. Conservation, regeneration and genetic stability of regenerants from alginate-encapsulated shoot explants of Gardenia jasminoides Ellis. Polymers 2021, 13, 1666. [Google Scholar] [CrossRef]
- Graf, W. Key for the main assortment of Ilex aquifolium cultivars in Central Europe based on leaf morphology. Acta Hortic. 2008, 885, 123–130. [Google Scholar] [CrossRef]
- Klockars, A.J.; Sax, G. Post hoc comparisons: The Scheffe test. In Multiple Comparisons; Klockars, A.J., Sax, G., Eds.; Sage Publications: Beverly Hills, CA, USA, 1986; Volume 61, pp. 58–61. [Google Scholar]
- Nei, M. Genetic distance between populations. Amer. Naturalist 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Wen, X.; Falush, D. Documentation for Structure Software: Version 2.3; University of Chicago: Chicago, IL, USA, 2010; p. 38. [Google Scholar]
- Baldoni, L.; Tosti, N.; Ricciolini, C.; Belaj, A.; Arcioni, S.; Pannelli, G.; Germana, M.A.; Mulas, M.; Porceddu, A. Genetic structure of wild and cultivated olives in the Central Mediterranean basin. Ann. Bot. 2006, 98, 935–942. [Google Scholar] [CrossRef]
- Ganopoulos, I.V.; Kazantzis, K.; Chatzicharisis, I.; Karayiannis, I.; Tsaftaris, A.S. Genetic diversity, structure and fruit trait associations in Greek sweet cherry cultivars using microsatellite based (SSR/ISSR) and morpho-physiological markers. Euphytica 2011, 181, 237–251. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA 4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Vlad, M.; Vlad, I.; Vlad, I.A.; Cheregi, G. Research on the improvement of propagation technology in Ilex aquifolium. Ann. Univ. Oradea 2015, 25, 141–144. [Google Scholar]
- Mbangcolo, M.M.; Reinten, E.Y.; Agenbag, G.A. Effect of species, cutting position and exogenous rooting substances on rooting of honeybush (Cyclopia spp.) cuttings. S. Afr. J. Plant Soil 2013, 30, 53–55. [Google Scholar] [CrossRef]
- Malan, D.G. Propagation of Proteaceae. Acta Hortic. 1992, 316, 27–34. [Google Scholar] [CrossRef]
- Brock, J.A.; Griffin, J.J. Rooting response of stem cuttings of Shantung maple (Acer truncatum) to time of year, cutting position, and auxin concentration, formulation, and solvent. J. Environ. Hort. 2014, 32, 163–166. [Google Scholar] [CrossRef]
- Tilahun, A.; Manahlie, B.; Abebe, G.; Negash, G. Effect of cutting position and indole butyric acid (auxin) concentration on rooting response of Araucaria heterophylla. Afr. J. Biotechnol. 2019, 18, 86–91. [Google Scholar]
- Stephen, O.U.; Chikordi, A.I. Age of stem cuttings and its effect on the growth of Manihot spp. World J. Agric. Sci. 2015, 3, 1–3. [Google Scholar]
- Mashudi, M.; Adinugraha, H.A. The growth success of Alstonia scholaris (L.) R. Br. shoot cuttings from several shoots position and the cut type of cuttings. J. Penel. Kehut. Wallacea 2015, 4, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Solikin, S. Effect of nodes position on the growth and yield of stem cutting of sambiloto (Andrographis paniculata). Nusantara Biosci. 2018, 10, 226–231. [Google Scholar] [CrossRef]
- Beemnet, M.K.; Aster, K.; Solomon, A.M. Effect of node number, part used and season of propagation on nursery establishment of lemon verbena (Aloysia triphylla L.). In Improving Quality Production of Horticultural Crops for Sustainable Development Proceedings, Jimma, Ethiopia, 4–5 February 2011; Jimma University College of Agriculture and Veterinary Medicine: Jimma, Ethiopia, 2012; pp. 121–133. [Google Scholar]
- Beemnet, M.K.; Solomon, A.M. Effect of cutting position and rooting hormone on propagation ability of stevia (Stevia rebaudiana Bertoni). Afr. J. Plant. Sci. Biotechnol. 2012, 6, 5–8. [Google Scholar]
- Zewdinesh, D.Z.; Beemnet, M.K.; Bazaznew, D. Effect of cutting size and position on propagation ability of lavender (Lavandula angustifolia L.). In Proceedings of the 4th Biennial Conference of Ethiopian Horticultural Science Society, Ambo, Ethiopia, 12–13 April 2013; pp. 129–137. [Google Scholar]
- Zewdinesh, D.Z.; Beemnet, M.K. Effect of cutting size and position on propagation ability of sage (Salvia officinalis L.). Int. J. Adv. Biol. Biomed. Res. 2016, 4, 68–76. [Google Scholar]
- Kostas, S.; Hatzilazarou, S.; Pipinis, E.; Vasileiadis, A.; Magklaras, P.; Smyrnioudis, I.; Vasilakis, T.; Chazakis, M.; Anastasiadi, V.; Ziogou, F.-T.; et al. Propagation of Pistacia lentiscus var. Chia genotypes and determination of their ornamental traits combined with a genetic analysis using ISSR markers. Agronomy 2021, 11, 205. [Google Scholar] [CrossRef]
- Huang, W.J.; Ning, G.G.; Liu, G.F.; Bao, M.Z. Determination of genetic stability of long-term micropropagated plantlets of Platanus acerifolia using ISSR markers. Biol. Plant 2009, 53, 159–163. [Google Scholar] [CrossRef]
- Martins, M.; Sarmento, D.; Oliveira, M.M. Genetic stability of micropropagated almond plantlets, as assessed by RAPD and ISSR markers. Plant Cell Rep. 2004, 23, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Modi, A.R.; Singh, A.S.; Gajera, B.B.; Patel, A.R.; Patel, M.P.; Subhash, N. Assessment of genetic fidelity of micropropagated date palm (Phoenix dactylifera L.) plants by RAPD and ISSR markers assay. Physiol. Mol. Biol. Plants 2010, 16, 207–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solano, M.C.P.; Ruiz, J.S.; Arnao, M.T.G.; Castro, O.C.; Tovar, M.E.G.; Bello, J.J.B. Evaluation of in vitro shoot multiplication and ISSR marker based assessment of somaclonal variants at different subcultures of vanilla (Vanilla planifolia Jacks). Physiol. Mol. Biol. Plants 2019, 25, 561–567. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; Kostas, S.; Joachim, M.; Economou, A. Regeneration of Viburnum dentatum L. from alginate-encapsulated shoot explants after short-term cold storage and assessment of genetic stability using ISSR analysis. Agronomy 2020, 10, 1660. [Google Scholar] [CrossRef]
- Saeed, T.; Shahzad, A.; Sharma, S. Studies on single and double layered biocompatible encapsulation of somatic embryos in Albizia lebbeck and genetic homogeneity appraisal among synseed derived lines through ISSR markers. Plant Cell Tiss. Org. Cult. 2020, 140, 431–445. [Google Scholar] [CrossRef]
- Zografou, P.; Linos, A.; Hagidimitriou, M. Genetic diversity among different genotypes of Pistacia lentiscus var. Chia (mastic tree). Options Mediterr. 2010, 94, 159–163. [Google Scholar]
- Abuduli, A.; Aydin, Y.; Sakiroglu, M.; Onay, A.; Ercisli, S.; Uncuoglu, A.A. Molecular evaluation of genetic diversity in wild-type mastic tree (Pistacia lentiscus L.). Biochem. Genet. 2016, 54, 619–635. [Google Scholar] [CrossRef] [PubMed]
- Araujo, F.S.; Pacheco, M.V.; Vieira, F.A.; Ferrari, C.S.; Felix, F.C.; Chagas, K.P.T. ISSR molecular markers for the study of the genetic diversity of Mimosa caesalpiniaefolia Benth. Idesia 2016, 34, 47–52. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | Tm (°C) | Number of Amplified Bands | Number of Polymorphic Bands | Polymorphism (%) | |
|---|---|---|---|---|---|---|
| 1 | UBC-809 | AGA GAG AGA GAG AGA GG | 54 | 14 | 13 | 92.86 |
| 2 | UBC-810 | GAG AGA GAG AGA GAG AT | 52 | 9 | 8 | 88.89 |
| 3 | UBC-815 | CTC TCT CTC TCT CTC TG | 50 | 13 | 10 | 76.92 |
| 4 | UBC-816 | CAC ACA CAC ACA CAC AT | 54 | 12 | 10 | 83.33 |
| 5 | UBC-818 | CAC ACA CAC ACA CAC AG | 56 | 13 | 11 | 84.62 |
| 6 | UBC-834 | AGA GAG AGA GAG AGA GYT | 56 | 8 | 5 | 62.50 |
| 7 | UBC-841 | GAG AGA GAG AGA GAG AYC | 54 | 11 | 10 | 90.91 |
| 8 | UBC-845 | CTC TCT CTC TCT CTC TRG | 50 | 7 | 1 | 14.29 |
| Rooting (%) | Number of Roots | Length of Roots | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Factors | df | F | Sign. | df | F | Sign. | df | F | Sign. |
| Season | 1 | 0.673 | NS | 1 | 0.116 | NS | 1 | 19.870 | *** |
| Auxin | 1 | 215.606 | *** | 1 | 156.775 | *** | 1 | 41.295 | *** |
| Season x Auxin | 1 | 12.587 | ** | 1 | 16.869 | *** | 1 | 12.535 | ** |
| Season | Rooting (%) | Number of Roots | Length of Roots (cm) | |||
|---|---|---|---|---|---|---|
| Control | 0.2% ΝAA | Control | 0.2% ΝAA | Control | 0.2% ΝAA | |
| Spring | 37.5 ± 10.4 B 1 a 2 | 93.7 ± 6.3 A a | 8.4 ± 5.1 B a | 20.3 ± 2.2 A b | 3.0 ± 1.1 A a | 3.8 ± 1.2 A a |
| Autumn | 13.7 ± 7.5 B b | 100 ± 0.0 A a | 2.1 ± 1.2 B b | 25.7 ± 2.9 A a | 1.2 ± 0.6 B b | 3.4 ± 0.9 A a |
| Rooting (%) | Number of Roots | Length of Roots (cm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Factors | df | F | Sign. | df | F | Sign. | df | F | Sign. |
| Type of cutting | 1 | 29.471 | *** | 1 | 1.451 | NS | 1 | 0.516 | NS |
| Auxin | 1 | 639.500 | *** | 1 | 115.781 | *** | 1 | 104.982 | *** |
| Type of cutting x Auxin | 1 | 25.130 | *** | 1 | 0.747 | NS | 1 | 0.002 | NS |
| Type of Cutting | Rooting (%) | Number of Roots | Length of Roots (cm) | |||
|---|---|---|---|---|---|---|
| Control | 0.2% ΝAA | Control | 0.2% ΝAA | Control | 0.2% ΝAA | |
| Terminal | 13.7 ± 7.5 B 1 b 2 | 100 ± 0.0 A a | 2.1 ± 1.2 B a | 25.7 ± 2.9 A a | 1.2 ± 0.6 B a | 3.4 ± 0.9 A a |
| Subterminal | 12.5 ± 6.4 B b | 83.7 ± 2.5 A b | 1.4 ± 0.7 B a | 21.5 ± 1.2 A b | 1.3 ± 0.8 B a | 3.6 ± 1.0 A a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsaktsira, M.; Chavale, E.; Kostas, S.; Pipinis, E.; Tsoulpha, P.; Hatzilazarou, S.; Ziogou, F.-T.; Nianiou-Obeidat, I.; Iliev, I.; Economou, A.; et al. Vegetative Propagation and ISSR-Based Genetic Identification of Genotypes of Ilex aquifolium ‘Agrifoglio Commune’. Sustainability 2021, 13, 10345. https://doi.org/10.3390/su131810345
Tsaktsira M, Chavale E, Kostas S, Pipinis E, Tsoulpha P, Hatzilazarou S, Ziogou F-T, Nianiou-Obeidat I, Iliev I, Economou A, et al. Vegetative Propagation and ISSR-Based Genetic Identification of Genotypes of Ilex aquifolium ‘Agrifoglio Commune’. Sustainability. 2021; 13(18):10345. https://doi.org/10.3390/su131810345
Chicago/Turabian StyleTsaktsira, Maria, Eleana Chavale, Stefanos Kostas, Elias Pipinis, Parthena Tsoulpha, Stefanos Hatzilazarou, Fotios-Theocharis Ziogou, Irini Nianiou-Obeidat, Ivan Iliev, Athanasios Economou, and et al. 2021. "Vegetative Propagation and ISSR-Based Genetic Identification of Genotypes of Ilex aquifolium ‘Agrifoglio Commune’" Sustainability 13, no. 18: 10345. https://doi.org/10.3390/su131810345
APA StyleTsaktsira, M., Chavale, E., Kostas, S., Pipinis, E., Tsoulpha, P., Hatzilazarou, S., Ziogou, F. -T., Nianiou-Obeidat, I., Iliev, I., Economou, A., & Scaltsoyiannes, A. (2021). Vegetative Propagation and ISSR-Based Genetic Identification of Genotypes of Ilex aquifolium ‘Agrifoglio Commune’. Sustainability, 13(18), 10345. https://doi.org/10.3390/su131810345