
Particulate Matter Removal of Three Woody Plant Species, Ardisia crenata, Ardisia japonica, and Maesa japonica


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. PM Reduction
2.3. Plant Growth Traits
Chlorophyll b = (21.5 × A646.8 − 5.1 × A663.2)
Carotenoid = (1000 × A470 − 1.82 × chl a − 85.02 × chl b)/198
2.4. Statistical Analysis
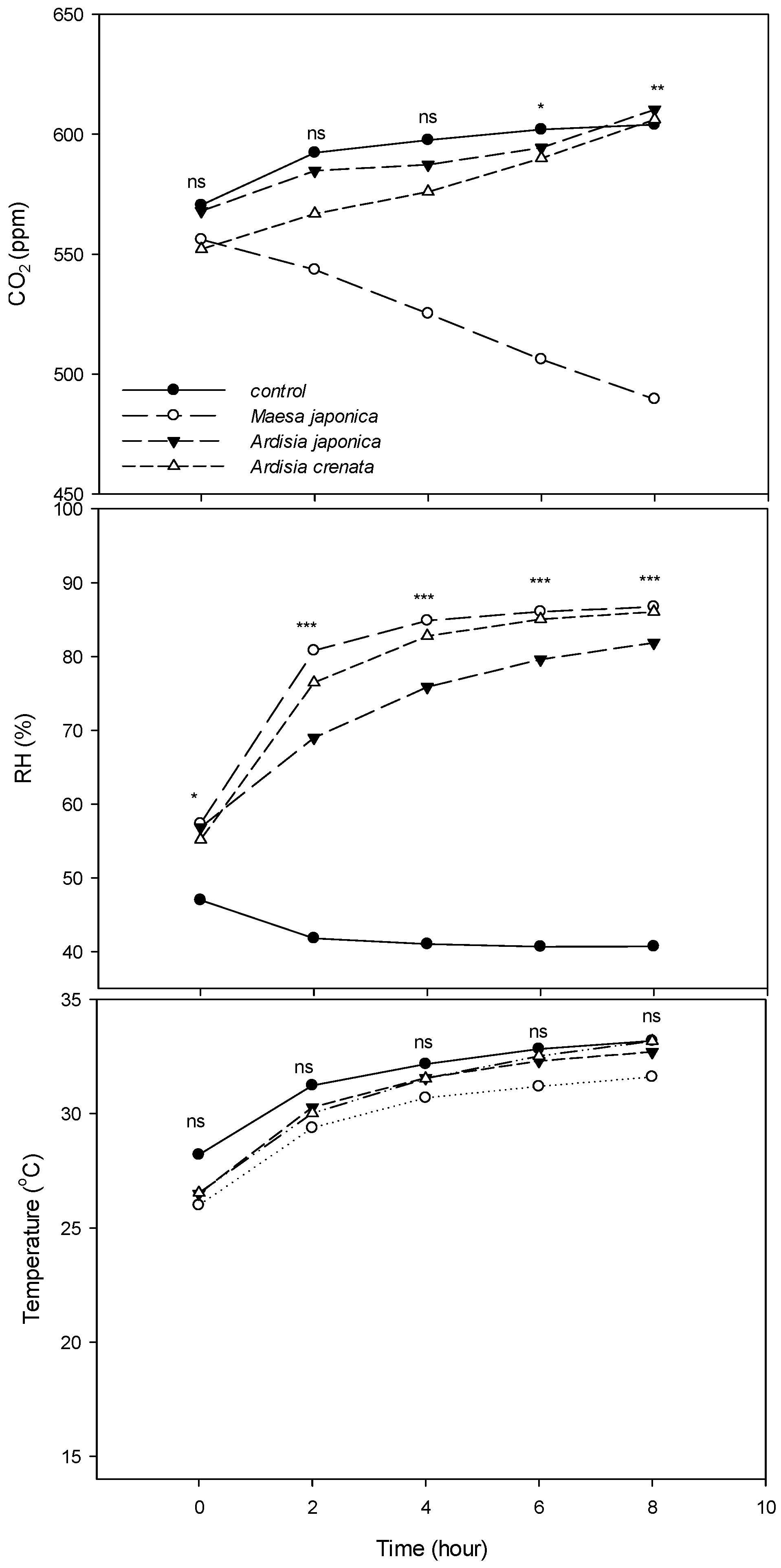
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Agency for Research on Cancer (IARC). Outdoor Air Pollution a Leading Environmental Cause of Cancer Deaths. 2013. Available online: http://www.iarc.fr/en/media-centre/pr/2013/pdfs/pr221_E.pdf (accessed on 20 September 2018).
- BéruBé, K.A.; Sexton, K.J.; Jones, T.P.; Moreno, T.; Anderson, S.; Richards, R.J. The spatial and temporal variations in PM10 mass from six UK home. Sci. Total Environ. 2004, 324, 41–53. [Google Scholar] [CrossRef]
- Kyotani, T.; Iwatsuki, M. Characterization of soluble and insoluble components in PM2.5 and PM10 fractions of airborne particulate matter in Kofu city, Japan. Atmos. Environ. 2002, 36, 639–649. [Google Scholar] [CrossRef]
- Seaton, A.; Macnee, W.; Donaldson, K.; Godden, D. Particulate air pollution and acute health effects. Lancet 1995, 345, 176–178. [Google Scholar] [CrossRef]
- Airkorea. Ambient Air Environment Standard. 2013. Available online: https://www.airkorea.or.kr/web/pmRelay?itemCode=10007&pMENUNO=108 (accessed on 23 November 2018).
- Kwon, K.J.; Urrintuya, O.; Kim, S.Y.; Yang, J.C.; Sung, J.W.; Park, B.J. Removal potential of particulate matter of 12 woody plant species for landscaping planting. J. People Plants Environ. 2020, 23, 647–654. [Google Scholar] [CrossRef]
- Kwon, K.J.; Odsure, U.; Kim, S.Y.; Yang, J.C.; Park, B.J. Comparison of the particulate matter removal capacity of 11 herbaceous landscape plants. J. People Plants Environ. 2021, 24, 267–275. [Google Scholar] [CrossRef]
- Jeong, N.R.; Kim, J.H.; Han, S.W.; Kim, J.C.; Kim, W.Y. Assessment of the particulate matter reduction potential of climbing plants on green walls for air quality management. J. People Plants Environ. 2021, 24, 377–387. [Google Scholar] [CrossRef]
- Popek, R.; Przybysz, A.; Gawrońska, H.; Klamkowski, K.; Gawroński, S.W. Impact of particulate matter accumulation on the photosynthetic apparatus of roadside woody plants growing in the urban conditions. Ecotoxicol. Environ. Saf. 2018, 163, 56–62. [Google Scholar] [CrossRef]
- Dzierżanowski, K.; Popek, R.; Gawrońska, H.; Sæbø, A.; Gawroński, S.W. Deposition of particulate matter of different size fractions on leaf surfaces and in waxes of urban forest species. Int. J. Phytoremediation 2011, 13, 1037–1046. [Google Scholar] [CrossRef]
- Leonard, R.J.; McArthur, C.; Hochuli, D.F. Particulate matter deposition on roadside plants and the importance of leaf trait combinations. Urban For. Urban Green. 2016, 20, 249–253. [Google Scholar] [CrossRef]
- Weber, F.; Kowarik, I.; Säumel, I. Herbaceous plants as filters: Immobilization of particulates along urban street corridors. Environ. Pollut. 2014, 186, 234–240. [Google Scholar] [CrossRef]
- Kaplan, R.; Kaplan, S. The Experience of Nature: A Psychological Perspective; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Lowry, C.A.; Hollis, J.H.; de Vries, A.; Pan, B.; Brunet, L.R.; Hunt, J.R.F.; Paton, J.; van Kampen, E.; Knight, D.; Evans, A.; et al. Identification of an immune-responsive mesolimbocortical serotonergic system: Potential role in regulation of emotional behavior. Neuroscience 2007, 146, 756–772. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Sun, C.; Zhou, X.; Leng, H.; Lian, Z. The effect of indoor plants on human comfort. Indoor Built Environ. 2014, 23, 709–723. [Google Scholar] [CrossRef]
- Ulrich, R.S. Aesthetic and affective response to natural environment. In Behavior and the Natural Environment, Human Behavior and Environment (Advances in Theory and Research); Altman, I., Wohlwill, J.F., Eds.; Springer: Boston, MA, USA, 1983; Volume 6. [Google Scholar] [CrossRef]
- Moya, T.A.; van den Dobbelsteen, A.; Ottelé, M.; Bluyssen, P.M. A review of green systems within the indoor environment. Indoor Built Environ. 2019, 28, 298–309. [Google Scholar] [CrossRef]
- Perez, G.; Rincon, L.; Vila, A.; Gonzalez, J.M.; Cabeza, L.F. Green vertical systems for buildings as passive systems for energy savings. Appl. Energy 2011, 88, 4854–4859. [Google Scholar] [CrossRef]
- Kim, S.J. The introduction of south-plants in indoor space. J. Korean Inst. Inter. Landsc. Archit. 2001, 3, 67–70. [Google Scholar]
- Yang, Y.P. An enumeration of Myrsinaceae of Taiwan. Bot. Bull. Acad. Sin. 1999, 40, 39–47. [Google Scholar]
- Lee, A.K.; Suh, J.K. Ecological studied on Ardisia species native to Korea and the significance as a potential indoor landscape corp. J. Korean Inst. Landsc. Archit. 1999, 1, 59–77. [Google Scholar]
- Lee, J.S.; Oh, H.W. Using trend of Korean native plants for interior landscape in Korea. Flower Res. 2002, 10, 91–96. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, Z.; Teng, M.; Wang, P.; Zhou, L. Accumulation of three different sizes of particulate matter on plant leaf surfaces: Effect on leaf traits. Arch. Biol. Sci. 2015, 67, 1257–1267. [Google Scholar] [CrossRef]
- Paek, K.Y.; Jun, Y.S. Stomatal Density, Size and Morphological Characteristics in Orchids Stomatal density, size and morphological characteristics in orchids. J. Korean Soc. Hortic. Sci. 1995, 36, 851–862. [Google Scholar]
- Gawrońska, H.; Bakera, B. Phytoremediation of particulate matter from indoor air by Chlorophytum comosum L. plants. Air Qual. Atmos. Health 2015, 8, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, N.R.; Kim, K.J.; Yoon, J.H.; Han, S.W.; You, S. Evaluation on the potential of 18 species of indoor plants to reduce particulate matter. J. People Plants Environ. 2020, 23, 637–646. [Google Scholar] [CrossRef]
- Son, D.; Kim, K.J.; Jeong, N.R.; Yun, H.G.; Han, S.W.; Kim, J.; Do, G.R.; Lee, S.H.; Shagol, C.C. The impact of the morphological characteristics of leaves on particulate matter removal efficiency of plants. J. People Plants Environ. 2019, 22, 551–561. [Google Scholar] [CrossRef]
- Lou, C.; Liu, H.; Li, Y.; Peng, Y.; Wang, J.; Dai, L. Relationships of relative humidity with PM2.5 and PM10 in the Yangtze River Delta, China. Environ. Monit. Assess. 2017, 189, 582. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Choi, B.; Chun, M.Y. Stabilizing soil moisture and indoor air quality purification in a wall-typed botanical biofiltration system controlled by humidifying cycle. Korean J. Hortic. Sci. Technol. 2015, 33, 605–617. [Google Scholar] [CrossRef] [Green Version]
- Przybysz, A.; Popek, R.; Gawrońsk, H.; Grab, K.; Łoskot, K.; Wrochna, M.; Gawronski, S.W. Efficiency of photosynthetic apparatus of plants grown in sites differing in level of particulate matter. Acta Sci. Pol. Hortorum Cultus 2014, 13, 17–30. [Google Scholar]
- Freer-Smith, P.H.; Holloway, S.; Goodman, A. The uptake of particulates by an urban woodland: Site description and particulate composition. Environ. Pollut. 1997, 95, 27–35. [Google Scholar] [CrossRef]
- Litschke, T.; Kuttler, W. On the reduction of urban particle concentration by vegetation—A review. Meteorol. Z. 2008, 17, 229–240. [Google Scholar] [CrossRef]
- Yoon, J.W.; Son, K.C.; Yang, D.S.; Kays, S.J. Removal of indoor tobacco smoke under light and dark conditions as affected by foliage plants. Korean J. Hortic. Sci. Technol. 2009, 27, 312–318. [Google Scholar]
- Hirano, T.; Kiyota, M.; Aiga, I. Physical effects of dust on leaf physiology of cucumber and kidney bean plants. Environ. Pollut. 1995, 89, 255–261. [Google Scholar] [CrossRef]
- Jeong, S.J.; Song, J.S.; Kim, W.S.; Lee, D.W.; Kim, H.D.; Kim, K.J.; Yoo, E.H.; Cho, J.G. Evaluation of selected foliage plants for improvement of indoor humidity. Hortic. Environ. Biotechnol. 2008, 49, 439–446. [Google Scholar]
- Fails, B.S.; Lewis, A.J.; Barden, J.A. Anatomy and morphology of sun-and shade-grown Ficus benjamina. J. Am. Soc. Hortic. Sci. 1982, 107, 754–757. [Google Scholar]
- Schmitz, H.; Hilgers, U.; Weidner, M. Assimilation and metabolism of formaldehyde by leaves appear unlikely to be of value for indoor air purification. New Phytol. 2000, 147, 307–315. [Google Scholar] [CrossRef]
- Son, K.; Lee, S.; Seo, S.; Song, J. Effects of foliage plants and potting soil on the absorption and adsorption of indoor air pollutants. J. Korean Soc. Hortic. Sci. 2000, 41, 305–310. [Google Scholar]
- Beckett, K.P.; Freer-Smith, P.H.; Taylor, G. Urban woodlands: Their role in reducing the effects of particulate pollution. Environ. Pollut. 1998, 99, 347–360. [Google Scholar] [CrossRef]
- Przybysz, A.; Sæbø, A.; Hanslin, H.M.; Gawroński, S.W. Accumulation of particulate matter and trace elements on vegetation as affected by pollution level, rainfall and the passage of time. Sci. Total Environ. 2014, 481, 360–369. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Plant Height (cm) | Leaf Length (cm) | Leaf Width (cm) | No. of Leaves (/Plant) | |
|---|---|---|---|---|
| Maesa japonica | 20.7 ± 2.1 a z | 9.1 ± 1.3 a | 3.9 ± 0.6 a | 52.4 ± 0.5 a |
| Ardisia japonica | 17.4 ± 1.9 b | 6.7 ± 1.1 b | 3.3 ± 0.3 b | 60.5 ± 1.6 a |
| Ardisia crenata | 15.9 ± 1.9 b | 7.0 ± 0.7 b | 2.7 ± 0.5 b | 57.1 ± 1.8 a |
| Significance | *** | *** | *** | ns |
| 0 h | 2 h | 4 h | 6 h | 8 h | ||
|---|---|---|---|---|---|---|
| PM1 | Control | 0.0121 a z | 0.0241 a | 0.0241 a | 0.0196 a | 0.0136 a |
| Maesa japonica | 0.0123 a | 0.0273 a | 0.0179 ab | 0.0097 b | 0.0050 b | |
| Ardisia japonica | 0.0117 ab | 0.0286 a | 0.0206 ab | 0.0121 b | 0.0071 b | |
| Ardisia crenata | 0.0100 b | 0.0257 a | 0.0157 b | 0.0086 b | 0.0044 b | |
| Significance | ns | ns | *** | ** | *** | |
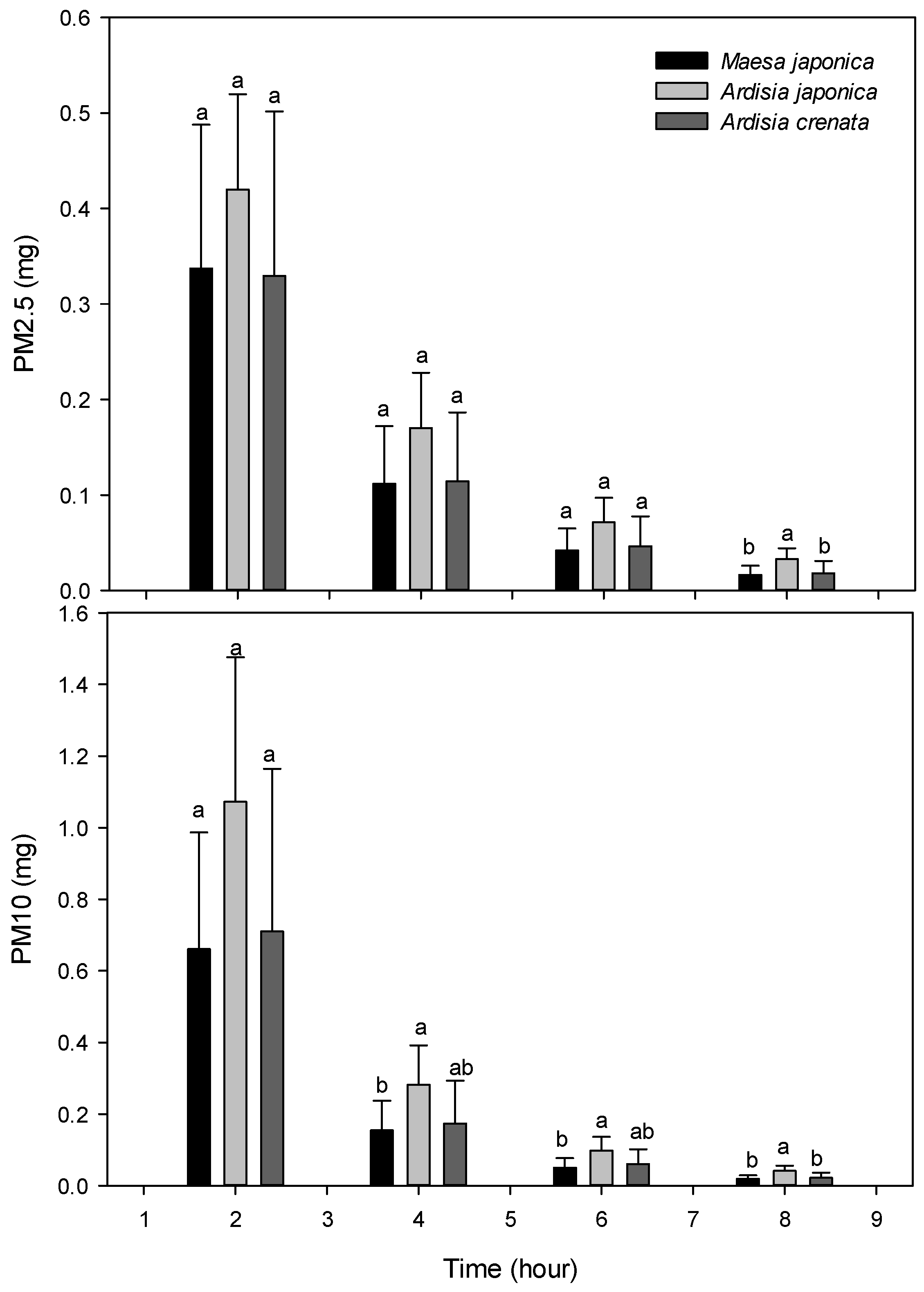
| PM2.5 | Control | 0.779 a | 0.631 a | 0.372 a | 0.199 a | 0.107 a |
| Maesa japonica | 0.827 a | 0.337 b | 0.112 b | 0.042 b | 0.017 b | |
| Ardisia japonica | 0.794 a | 0.420 b | 0.170 b | 0.071 b | 0.033 b | |
| Ardisia crenata | 0.815 a | 0.329 b | 0.114 b | 0.046 b | 0.018 b | |
| Significance | ns | ** | *** | *** | *** | |
| PM10 | Control | 9.999 a | 2.398 a | 0.889 a | 0.373 a | 0.173 a |
| Maesa japonica | 9.941 a | 0.661 b | 0.155 b | 0.050 b | 0.019 b | |
| Ardisia japonica | 9.999 a | 1.073 b | 0.282 b | 0.098 b | 0.042 b | |
| Ardisia crenata | 9.999 a | 0.710 b | 0.174 b | 0.060 b | 0.022 b | |
| Significance | ns | *** | *** | *** | *** |
| 0.2–2.5 μm | 2.5–10 μm | 10–100 μm | |
|---|---|---|---|
| Maesa japonica | 0.762 ± 0.439 a z | 1.689 ± 0.703 a | 4.867 ± 1.692 b |
| Ardisia japonica | 0.738 ± 0.338 a | 0.874 ± 0.355 b | 10.150 ± 3.065 a |
| Ardisia crenata | 0.804 ± 0.351 a | 0.907 ± 0.450 b | 6.600 ± 1.324 b |
| Significance | ns | ns | ** |
| Photosynthetic Rate (µmol m−2 s−1) | Transpiration Rate (mol m−2 s−1) | Stomatal Conductance (mol m−2 s−1) | |
|---|---|---|---|
| Maesa japonica | 5.894 ± 1.237 a z | 1.510 ± 0.361 a | 0.060 ± 0.019 a |
| Ardisia japonica | 0.862 ± 0.186 c | 0.020 ± 0.014 c | 0.000 ± 0.000 b |
| Ardisia crenata | 2.316 ± 1.128 b | 0.436 ± 0.306 b | 0.018 ± 0.013 b |
| Significance | *** | *** | *** |
| Chlorophyll Content (mg g−1 FW) | Carotenoid (mg g−1 FW) | ||
|---|---|---|---|
| Chlorophyll a | Chlorophyll b | ||
| Maesa japonica | 1.30 ± 0.36 a z | 2.55 ± 0.68 a | 2.53 ± 0.78 a |
| Ardisia japonica | 0.85 ± 0.08 b | 1.69 ± 0.16 b | 1.58 ± 0.17 b |
| Ardisia crenata | 1.21 ± 0.28 ab | 2.26 ± 0.53 ab | 2.27 ± 0.60 ab |
| Significance | ns | ns | ns |
| Fv/Fm | SLA (cm2 g−1) | Leaf pH | |
|---|---|---|---|
| Maesa japonica | 0.791 ± 0.016 a z | 155.6 ± 22.6 a | 5.39 ± 0.14 a |
| Ardisia japonica | 0.791 ± 0.010 a | 163.8 ± 24.8 a | 5.40 ± 0.19 a |
| Ardisia crenata | 0.772 ± 0.015 a | 140.1 ± 18.3 a | 5.41 ± 0.11 a |
| Significance | ns | ns | ns |
| Stomatal Number (/106 μm−2) | Stomatal Size (μm) | |
|---|---|---|
| Maesa japonica | 202.3 ± 37.2 a z | 12.3 ± 2.1 b |
| Ardisia japonica | 185.9 ± 20.9 a | 9.7 ± 1.6 b |
| Ardisia crenata | 133.0 ± 21.8 b | 21.1 ± 2.2 a |
| Significance | ** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, K.-J.; Kwon, H.-J.; Oh, Y.-A.; Kim, S.-Y.; Park, B.-J. Particulate Matter Removal of Three Woody Plant Species, Ardisia crenata, Ardisia japonica, and Maesa japonica. Sustainability 2021, 13, 11017. https://doi.org/10.3390/su131911017
Kwon K-J, Kwon H-J, Oh Y-A, Kim S-Y, Park B-J. Particulate Matter Removal of Three Woody Plant Species, Ardisia crenata, Ardisia japonica, and Maesa japonica. Sustainability. 2021; 13(19):11017. https://doi.org/10.3390/su131911017
Chicago/Turabian StyleKwon, Kei-Jung, Hyuk-Joon Kwon, Yun-Ah Oh, Soo-Young Kim, and Bong-Ju Park. 2021. "Particulate Matter Removal of Three Woody Plant Species, Ardisia crenata, Ardisia japonica, and Maesa japonica" Sustainability 13, no. 19: 11017. https://doi.org/10.3390/su131911017
APA StyleKwon, K. -J., Kwon, H. -J., Oh, Y. -A., Kim, S. -Y., & Park, B. -J. (2021). Particulate Matter Removal of Three Woody Plant Species, Ardisia crenata, Ardisia japonica, and Maesa japonica. Sustainability, 13(19), 11017. https://doi.org/10.3390/su131911017