1. Introduction
Due to global warming, high rearing temperature is becoming one of the most common issues in poultry production [
1]. Heat stress (HS) occurs when the ambient temperature exceeds the optimal range recommended for growth. Due to the high selection for fast growth performance, broilers in the last phase of rearing are especially sensitive to HS [
2]. Indeed, HS can impair the performance, metabolism, and health of broilers. The effects of HS can include reduced feed intake (FI), body weight gain (BW), and increased water consumption [
3]. Excessively high ambient temperature is strongly correlated with high mortality rates [
4]. In addition, previous reports have shown that HS induces oxidative damage by increasing the production of reactive oxygen species in cells [
5,
6].
Several strategies have been adopted to mitigate the deleterious effects of HS in poultry. The most common approaches include improvements in ventilation, the selection of resistant strains, and nutritional management [
7]. The latter consists of the dietary supplementation of various types of substances such as γ-aminobutyric acid (GABA). GABA is a four-carbon non-protein amino acid involved in inhibitory synaptic transmission [
8]. In addition, GABA has been implicated in several regulatory functions such as memory, blood pressure, and respiration [
9]. In broilers, GABA is used as a feed supplement to reduce the adverse effect of high environmental temperature [
10]. Previous research demonstrated its effectiveness for increasing overall growth performance, improving nutrient absorption, and reducing oxidative stress in chickens under high temperatures [
11,
12].
Recently, in ovo feeding has been considered a cost-effective way to alleviate the drawbacks of post-hatch and incubational HS in broilers. Originally, in ovo feeding was developed as a tool for providing a continuous supply of critical nutrients during the first few days of the chick’s life [
13]. The methodology consists of injecting nutrients into the amnion of eggs so that by consuming the content of the amnion, the embryo can access nutrients before it starts to hatch [
14]. The most recent advances involve testing the heat resistance of birds after the in ovo feeding of prebiotics [
15] or amino acids [
16]. Methionine–cysteine in ovo injection improved embryonic development, antioxidant status, and jejunum histomorphometry of broilers chicks exposed to incubational HS [
17]. Furthermore, the damages (high corticosterone and HSP70 mRNA levels) occasioned by incubational HS were alleviated after the in ovo feeding of sulfur-containing amino acids [
18]. Interestingly, evidence suggests that broilers’ resistance toward post-hatch HS can also be improved by in ovo feeding. For example, the in ovo feeding of L-leucine was proven effective in affording thermotolerance in broilers under acute heat stress, primarily by changing amino acid metabolism up to market age [
19]. In addition, in ovo injection of galactooligosaccharides reduced the harmful effects of hyperthermia on feed efficiency during the finisher feeding phase [
20]. When injected in ovo, we found that GABA could improve the early heat resistance of broilers by enhancing antioxidant functions and regulating fatty-acid-metabolism-related genes in 10-day-old chicks [
21].
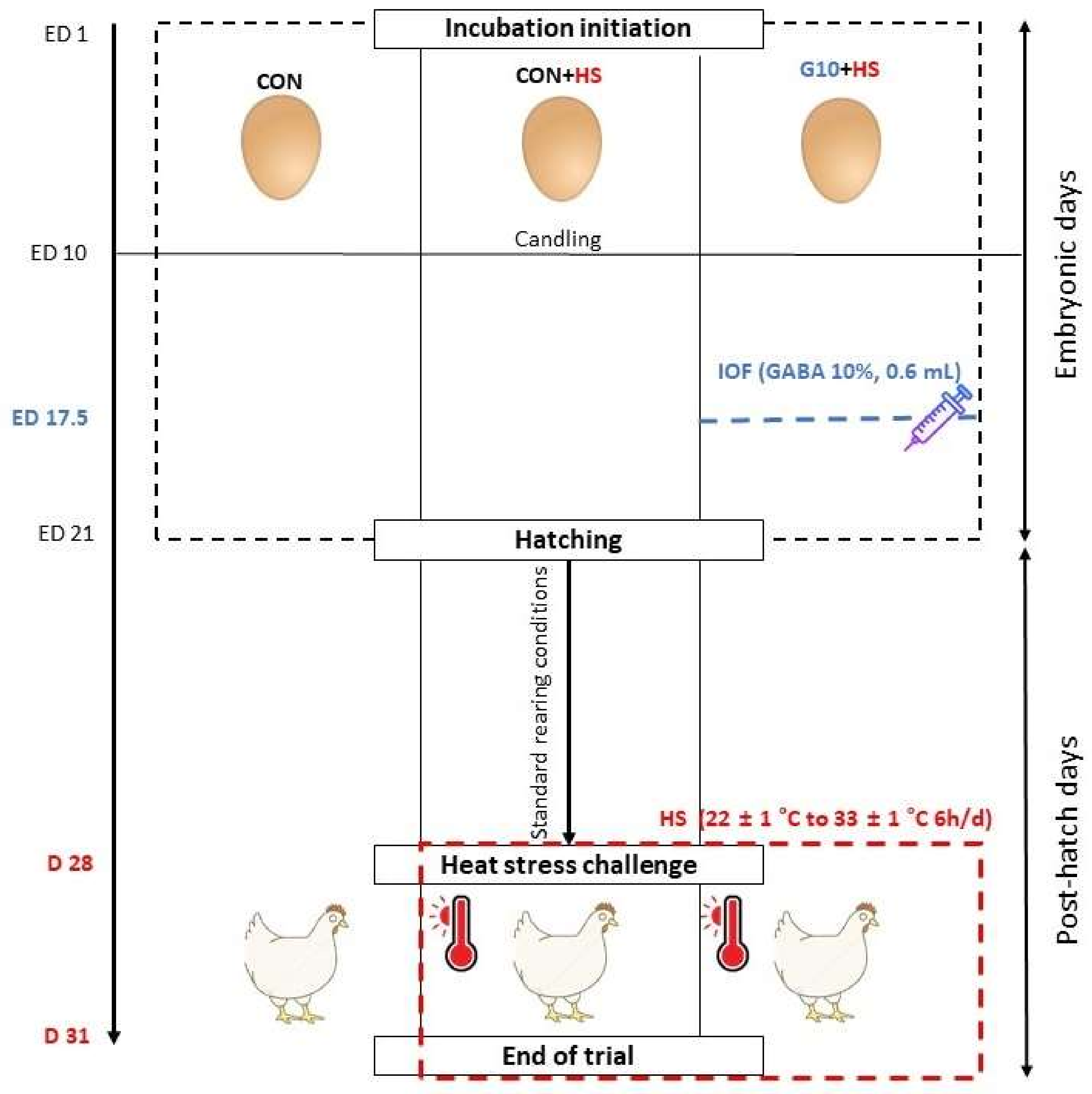
There have been no studies addressing the potential long-term effects of in ovo feeding of GABA in broilers. Therefore, the objective of this study was to investigate the effects of in ovo feeding of GABA on the growth performances, organ indexes, blood biochemical parameters, antioxidant status, and gene expression of stress-related genes in broilers subjected to cyclic HS near market age.
4. Discussion
In poultry, HS is known for its adverse impacts on health, physiology, and efficiency [
29,
30]. Chronic HS exposure induces a reduction in broilers’ feed intake and body weight while concomitantly increasing FCR [
31]. Similarly, in our study, 4 days of cyclic HS led to a significant reduction in ADFI of the birds reared under HS compared to those under normal housing temperature. For regulating their body temperature under HS, broilers reduce their feed consumption to promote heat loss and decrease their metabolic heat production [
32]. Therefore, even though reducing FI helps cope with high ambient temperature, it is naturally followed by severe growth impairment [
33]. These findings were confirmed in the results of our study. The ADG of heat-stressed birds was statistically lower (by 47.4% for CON+HS vs. 29.1% for G10+HS) compared to the control birds.
Interestingly, the in ovo feeding of GABA could mitigate the weight loss (by 35%) occasioned by cyclic HS. This might be explained by the fact that birds hatched from eggs injected in ovo recorded a higher ADFI (15.8%) than control birds under the same high environmental conditions. Additionally, our results showed that the in ovo feeding of GABA numerically reduced the FCR (15.9%) compared to the control birds under HS. Previous results have already documented the orexigenic effect of dietary GABA supplementation [
11,
34]. These results might suggest that the mechanism behind in ovo feeding is closely linked to how dietary supplementation works. Indeed, in ovo feeding given during the late embryonic stages targeted the amnion [
35]. The embryo, by consuming the content of the amnion, could access the injected nutrients before it started to hatch [
14]. GABA’s ability to improve the feed intake of HS-exposed birds was explained by the fact that it can lower the expression of anorexigenic neuropeptides and upregulate orexigenic neuropeptides [
11].
The organ index is an important parameter reflecting the development status of organs. A recent study [
36] reported an increase in the liver index when broilers were reared under chronic HS. There was a weakly significant (
p = 0.099) tendency to increase the liver index of HS birds in the present study. In poultry, the liver is known as the main organ of fat synthesis. Consequently, higher liver indexes might be correlated with higher fat synthesis during HS [
37]. Gastrointestinal length can be an indicator of digestive health. HS significantly decreased the absolute organ length of the duodenum, but its relative length was not affected. Therefore, no conclusive effect of either HS or the in ovo feeding of GABA on duodenum length could be ascertained. Rectal temperature is generally utilized as a marker for HS resistance in broilers [
38]. Previous reports have recognized the HS-stimulating effect of rectal temperature in broilers [
11,
34]. As expected, birds belonging to both groups under cyclic HS had a higher rectal temperature in the current study.
DPPH-RSA has been widely used as a method to evaluate plasma total antioxidant capacity (TAC) [
39]. In the present study, the in ovo feeding of GABA resulted in improved TAC in broilers subjected to cyclic HS. Researchers [
40] previously reported that GABA supplementation increased TAC in chickens under HS. These findings indicate that similarly to dietary GABA, in ovo GABA injection could instigate the retrieval of antioxidant functions after cyclic HS exposure. In response to cyclic HS, broilers exhibited higher plasma levels of corticosterone. These results of the present study show consistency with those reported by earlier studies [
41,
42], mentioning that heat-challenged birds tend to have increased plasma corticosterone compared to those at thermoneutral temperature. This might be explained by the fact that heat stress activates the hypothalamic–pituitary–adrenal (HPA) axis, leading to increased corticosterone secretion [
43]. Therefore, enhanced corticosterone release under HS may have reduced overall feed intake and body weight gain [
44,
45], as observed in the current study.
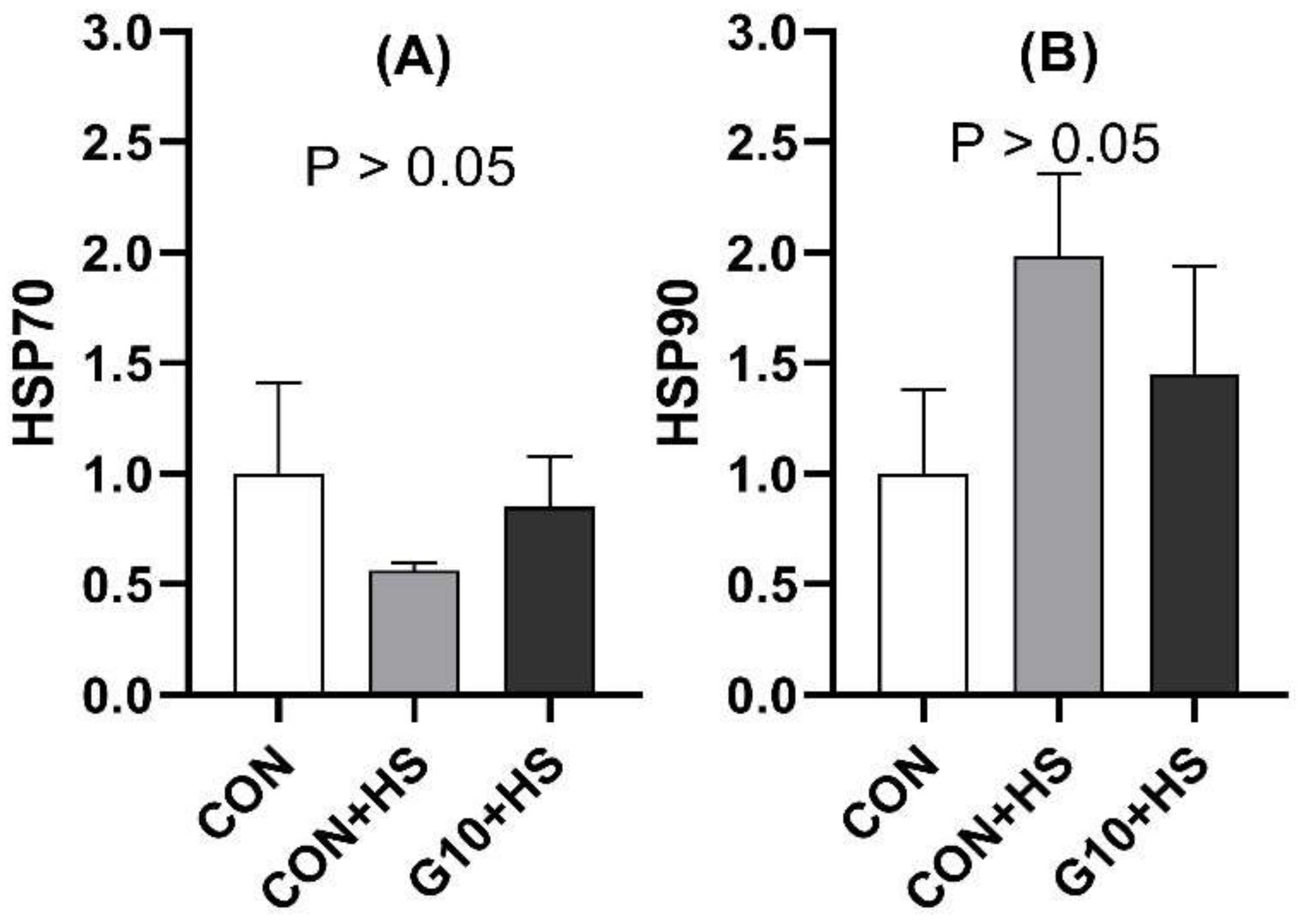
Interestingly, the in ovo feeding of GABA could reduce the plasma corticosterone levels of the birds exposed to HS. Earlier studies have revealed that dietary and GABA-enriched barley bran could lower adrenocorticotropic hormone (ACTH) and corticosterone levels in pigs and rats [
46,
47]. Indeed, under stressful conditions, the hypothalamus releases the corticotropin-releasing hormone, which stimulates the secretion of ACTH from the anterior pituitary gland [
48]. Subsequently, ACTH provokes the synthesis of glucocorticoids such as corticosterone to combat stress [
49]. Furthermore, GABA was shown to inhibit ACTH secretion by activating α1- and α2-adrenergic receptors [
50]. Thus, we could hypothesize that the potential effect of GABA on corticosterone is linked to its inhibitory effect on ACTH.
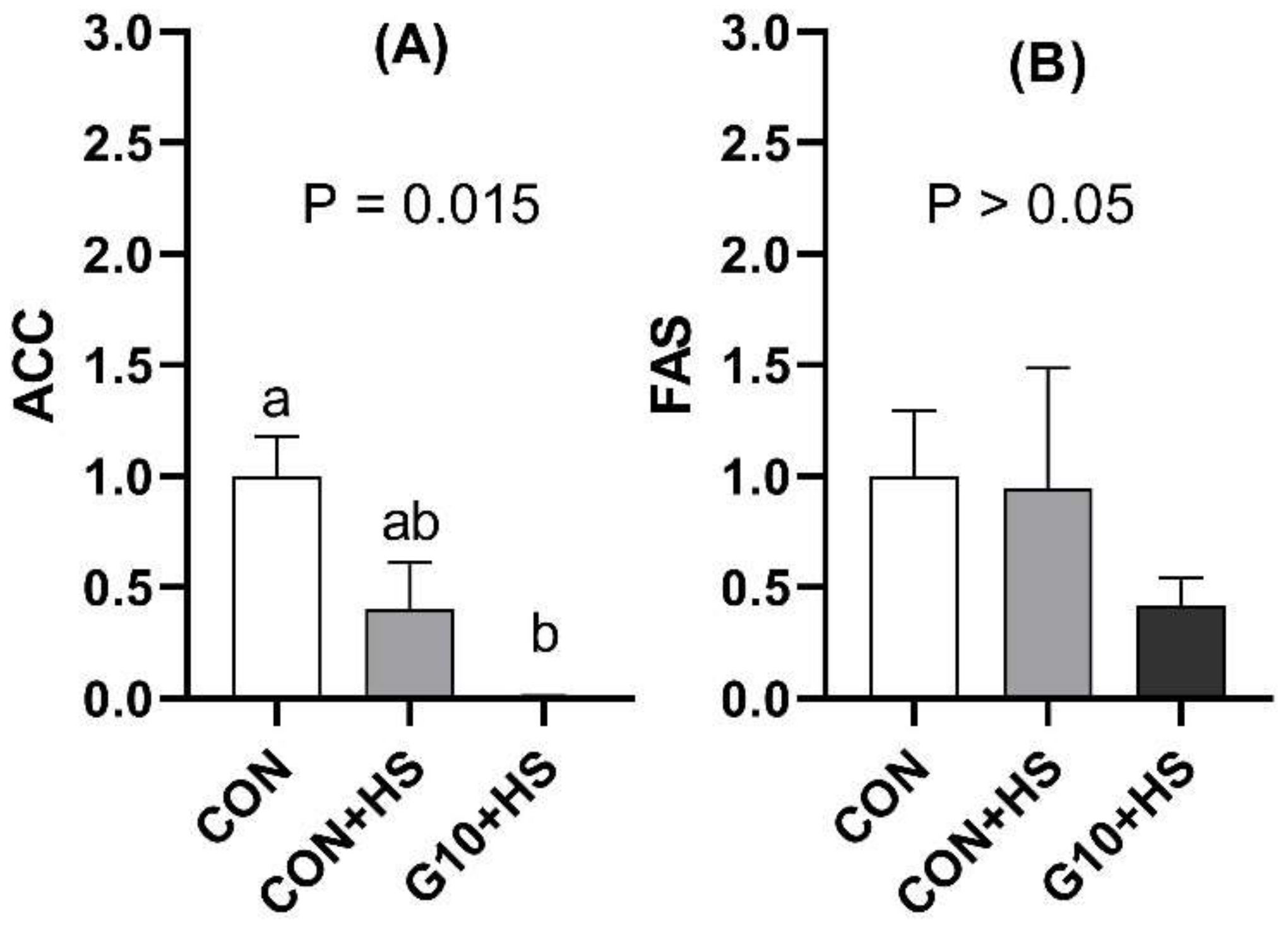
In poultry, acetyl-CoA carboxylase (ACC) and fatty acid synthase (FAS) are the key enzymes responsible for fat synthesis. ACC is involved in converting acetyl-CoA into malonyl-CoA and then into palmitate, whereas FAS determines the maximum capacity for fatty acid synthesis in tissues [
51]. Our results showed that the hepatic mRNA levels of ACC were downregulated in heat-stressed broilers after the in ovo injection of GABA. In addition, a numerically lower value of relative mRNA levels of FAS was observed in the birds fed GABA in ovo. As previously mentioned, the liver is known to be the main organ of fat synthesis in poultry, and one of the deleterious effects of HS in broilers is lipid accumulation, engendered by the upregulation of FAS and ACC genes [
36,
52]. Therefore, lower mRNA hepatic levels of FAS and ACC indicate less fatty acid synthesis during HS in birds that were fed GABA in ovo.
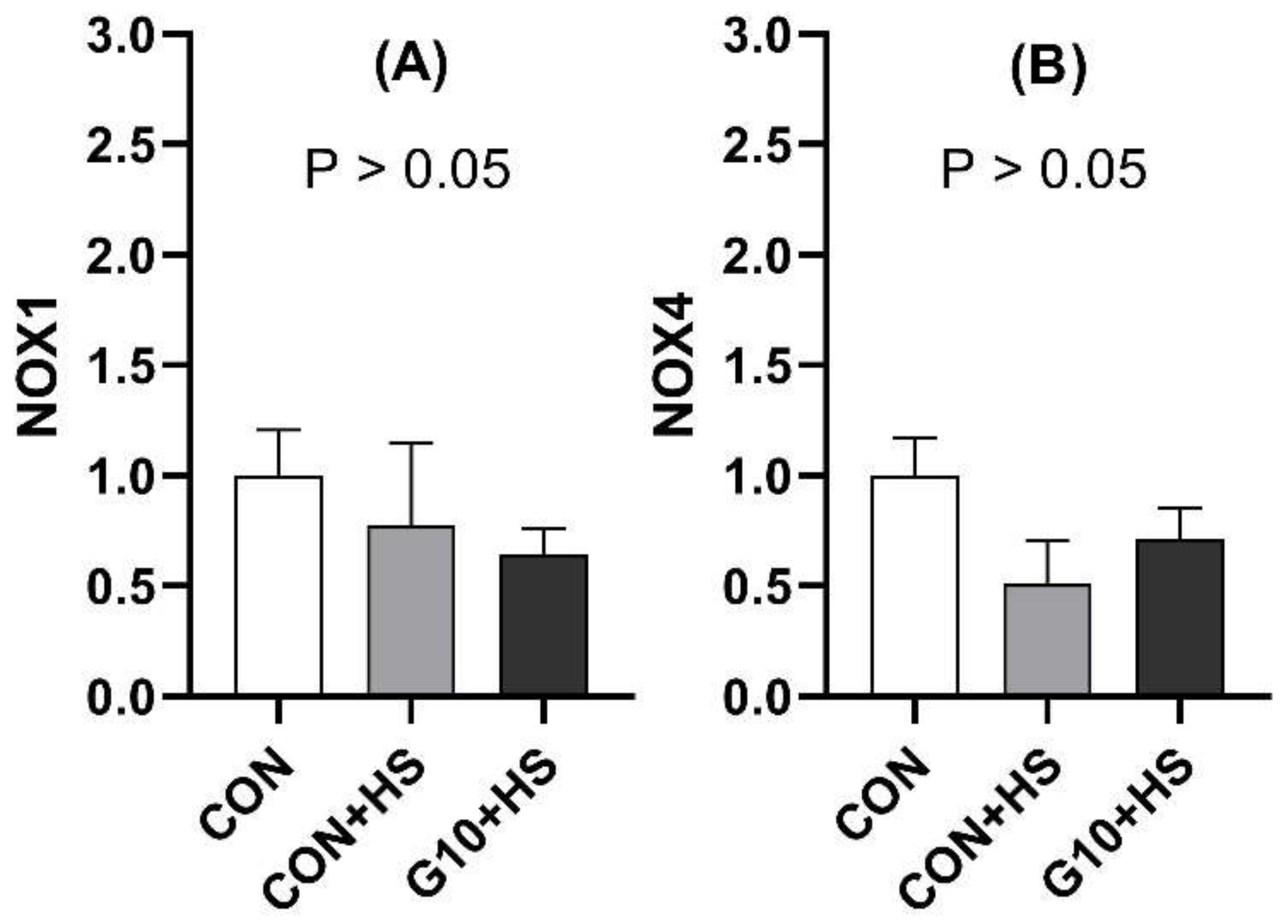
Interestingly, we observed a positive correlation between FAS and NOX1 mRNA levels. The NOX gene family, responsible for ROS generation, was affected by heat stress [
27], suggesting that the in ovo feeding of GABA may decrease ROS generation during cyclic HS. The mechanism by which GABA potentially reduces ROS generation could be related to its effect on glutathione peroxidase synthesis [
53,
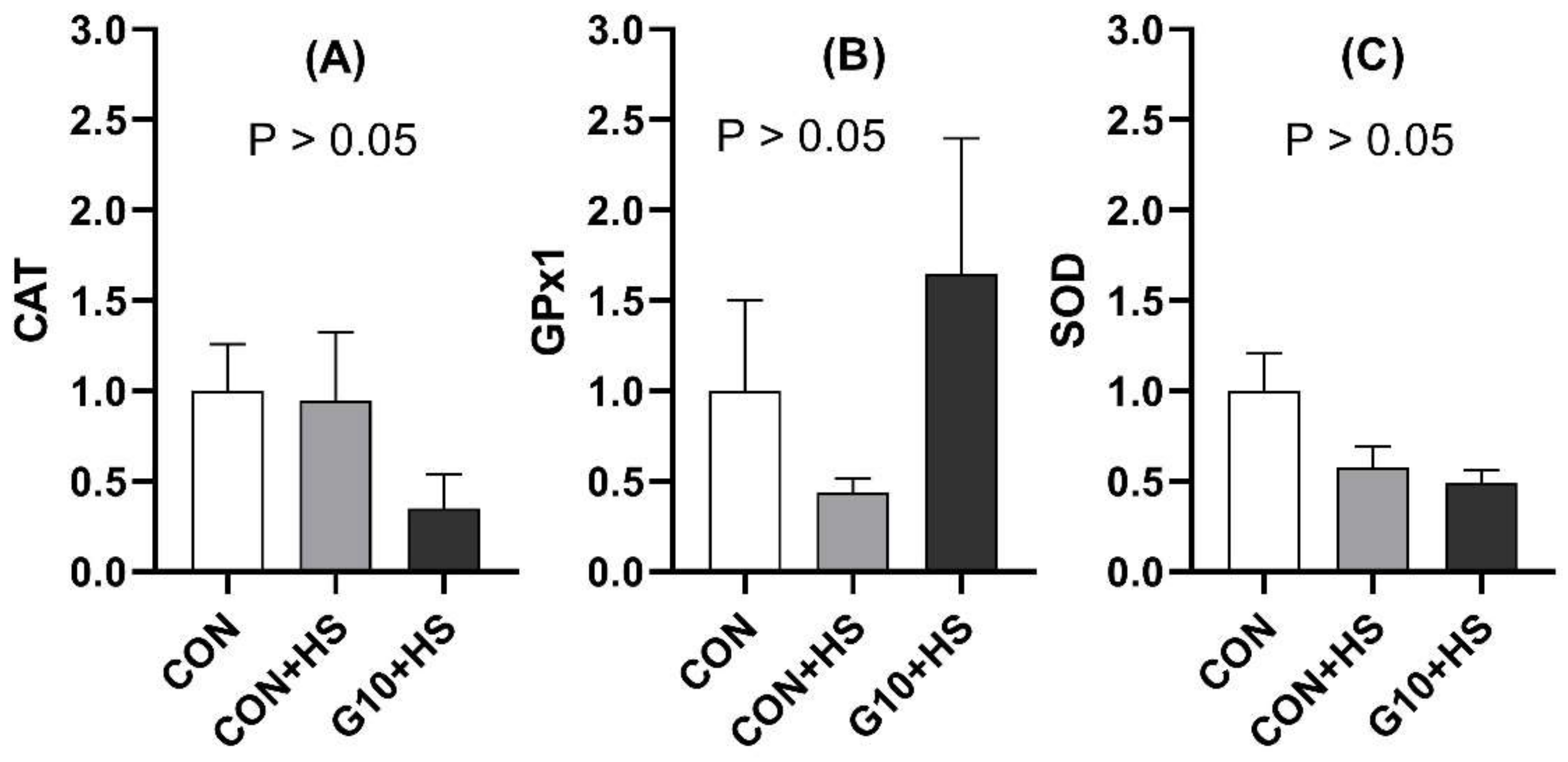
54]. Significant positive correlations were detected between CAT and SOD mRNA levels. SOD and CAT are among the major antioxidant enzymes [
55]. SOD is responsible for the dismutation of superoxide radicals to molecular oxygen and hydrogen peroxide (H
2O
2), whereas CAT is responsible for transforming H
2O
2 into water and oxygen [
56]. The actions of both enzymes are synchronized, which may justify the observed positive correlation.
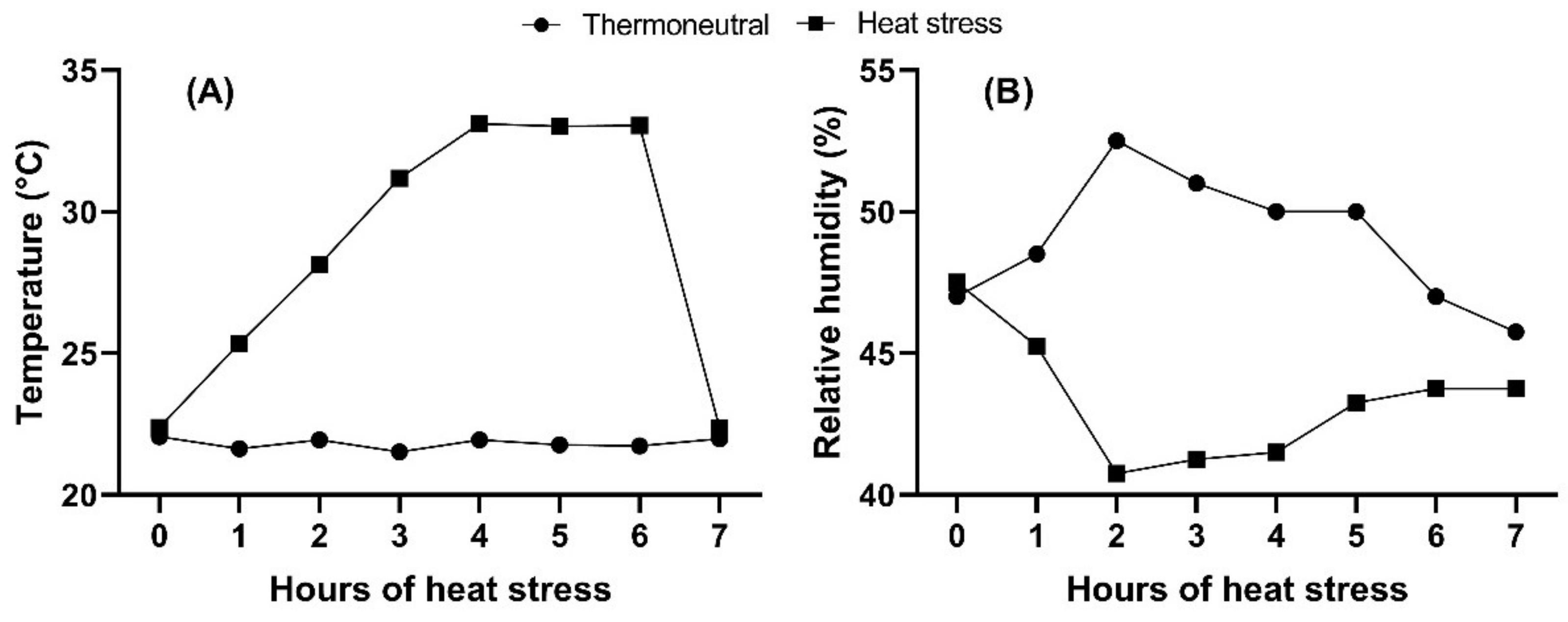
As the Materials and Methods section mentioned, a general blackout for 6 h led to a decrease in the temperature (from 36.8 to 20 °C) during the incubation. However, a potential effect of the incubation temperature drop can be neglected because all the treatment groups were kept in the same incubator. Even though a cold temperature during embryogenesis has been shown to enhance growth performance, such temperature manipulations were lower than 20 °C and repetitive to affect the thermoregulatory mechanism of chicks [
57,
58]. Furthermore, all the relevant data reported were cross-checked with the literature and showed no discrepancies in the current study. For example, the recorded growth performances of Arbor Acres in this study were concordant to recently published articles [
59,
60]. In addition, the average BWs obtained after hatch (45.1 g), 7 days (165.7 g), 21 days (981.1 g), and 28 days (1624.1 g) were in line with the Arbor Acres broiler performances (as-hatched performance) objective, which recommends 43 g, 204 g, 978 g, and 1567 g for the same periods, respectively [
61]. Similarly, plasma parameters [
62] were within the same range of previously reported results.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}