
Enhancing Symbiotic Nitrogen Fixation and Soybean Growth through Co-Inoculation with Bradyrhizobium and Pseudomonas Isolates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Bacteria from Root Nodules
2.2. Characterization of nifH and nodD Genes in Rhizobial Isolates
2.3. Nodulation Test of Rhizobial Isolates
2.4. Determination of Plant Growth-Promoting and Plant Constituent-Hydrolyzing Traits of Isolate Bra6 and Non-Rhizobial Isolates
2.5. Co-Inoculation Test in Sterile Water Agar through Flask Experiment
2.6. Co-Inoculation Test in Non-Sterile Strongly Acidic Soils through Pot Experiment
3. Results
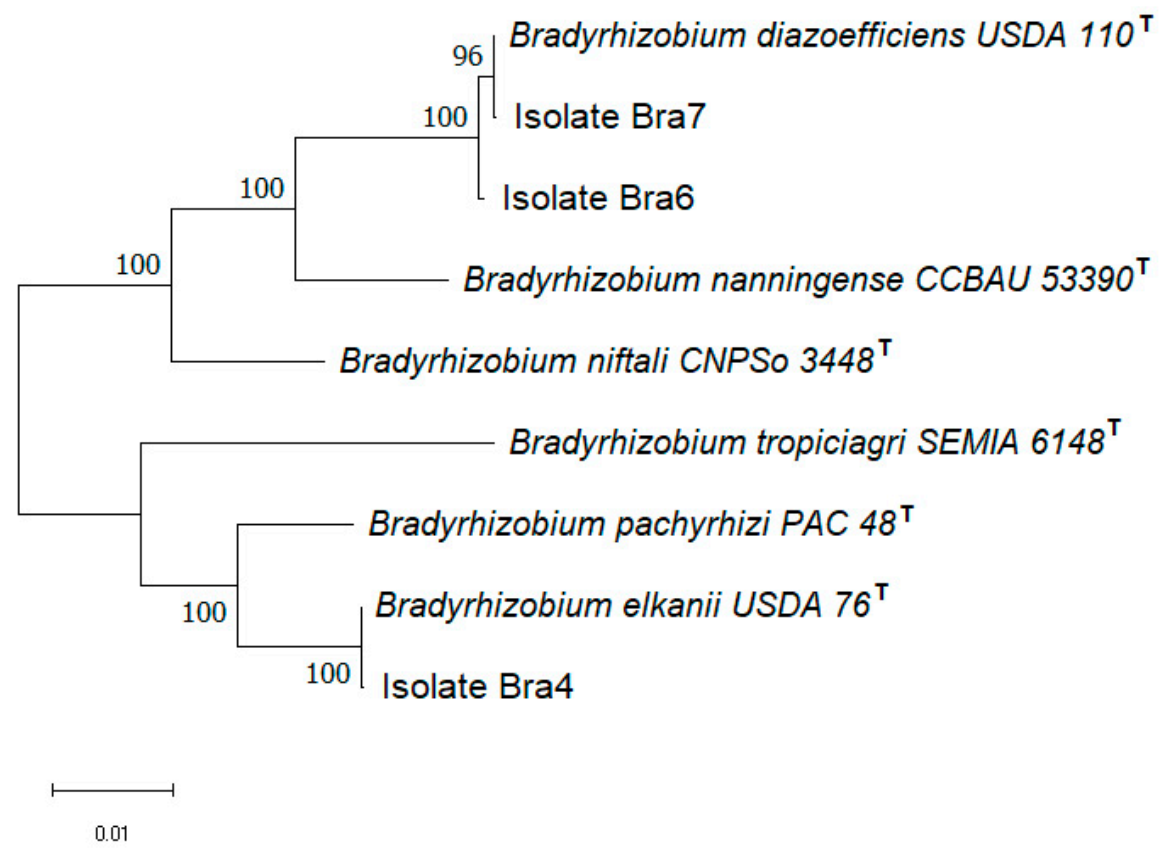
3.1. Screening and Identification of Root Nodules-Associated Isolates
3.2. Characterization of nifH and nodD Genes in Rhizobial Isolates
3.3. Screening of Effective Rhizobial Isolates Inducing Root Nodulation
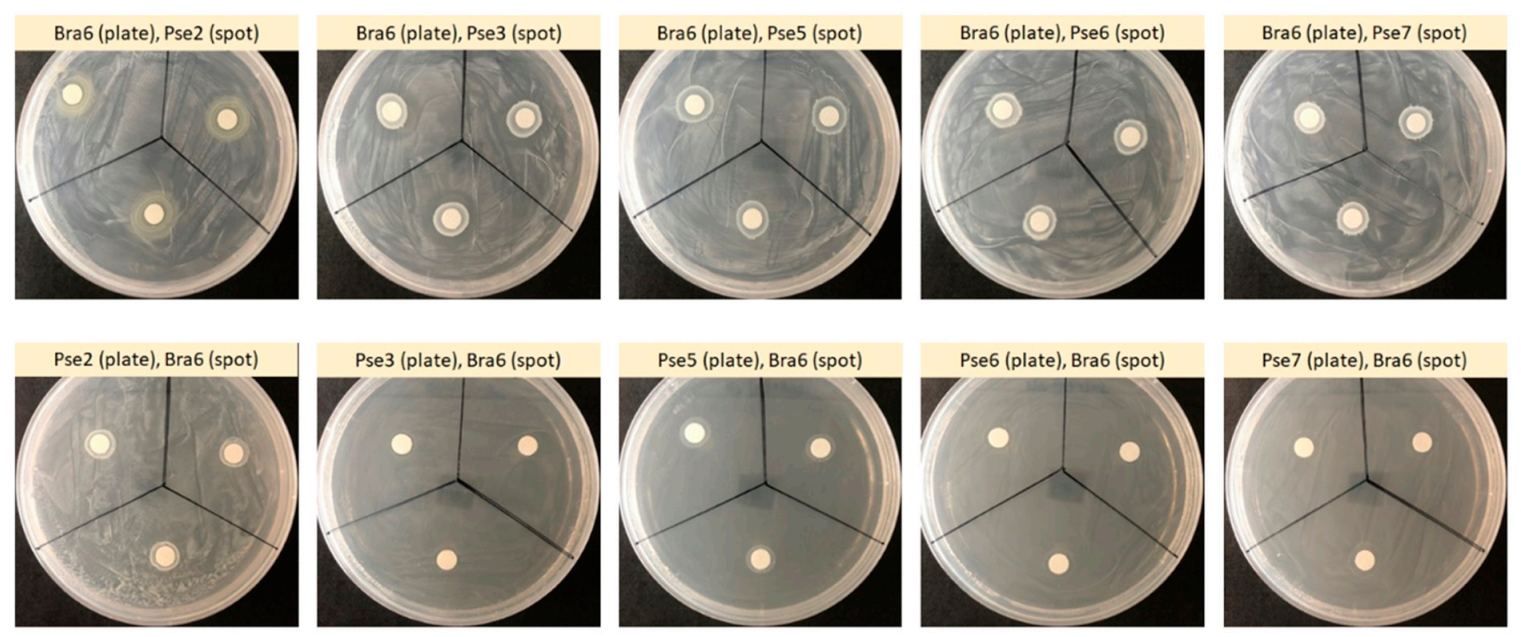
3.4. Plant Growth-Promoting and Plant Constituent-Hydrolyzing Traits of Isolate Bra6 and Non-Rhizobial Isolates
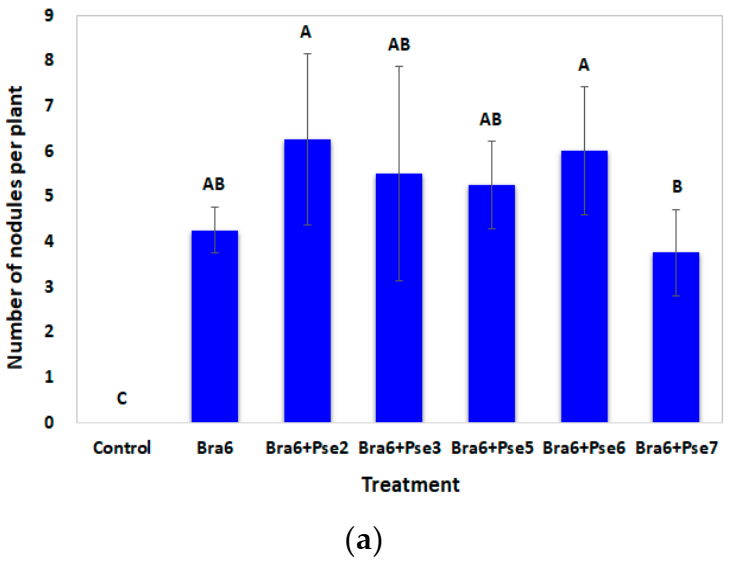
3.5. Influence of Co-Inoculation on Nodulation and Soybean Growth in Sterile Water Agar Evaluated through Flask Experiment
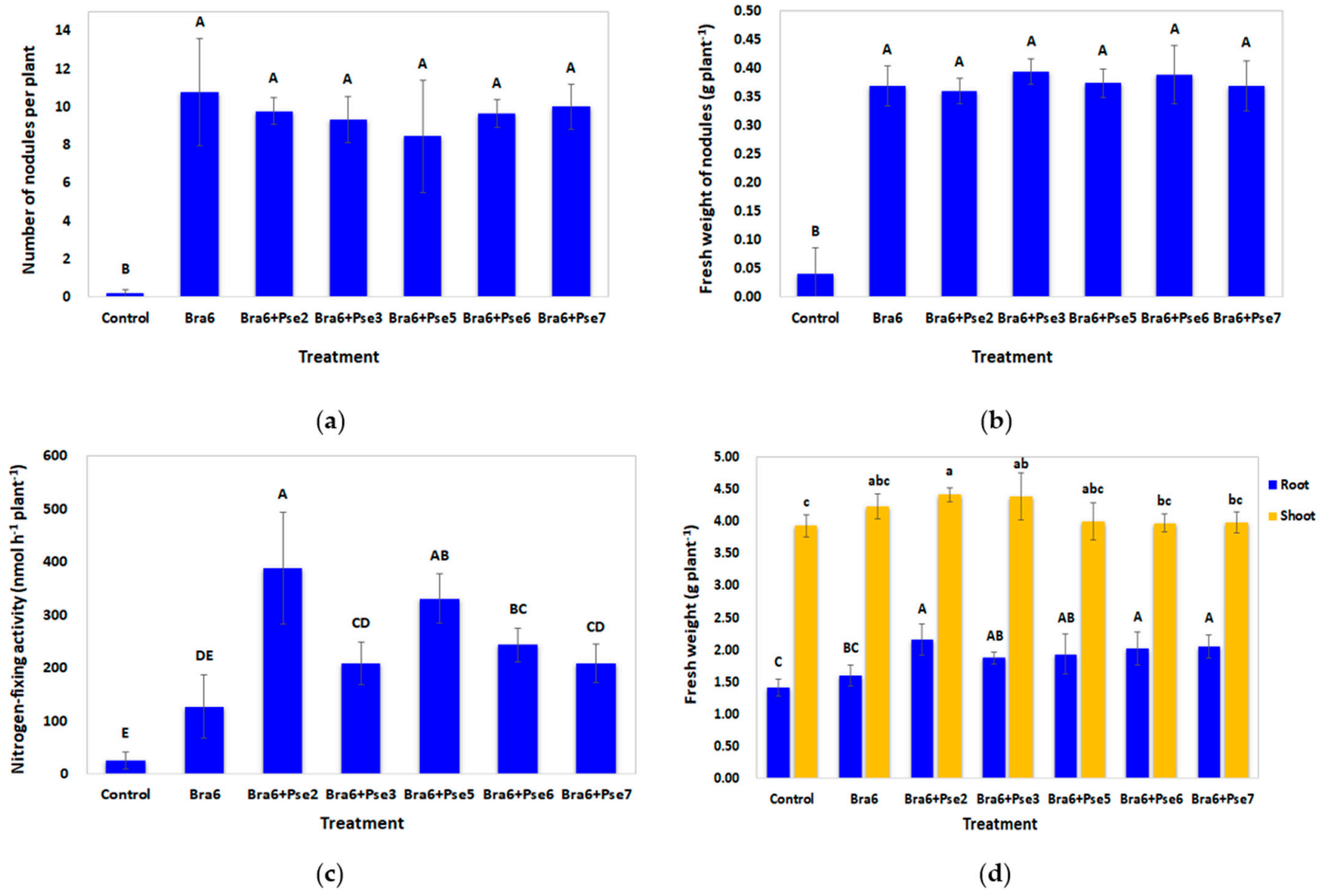
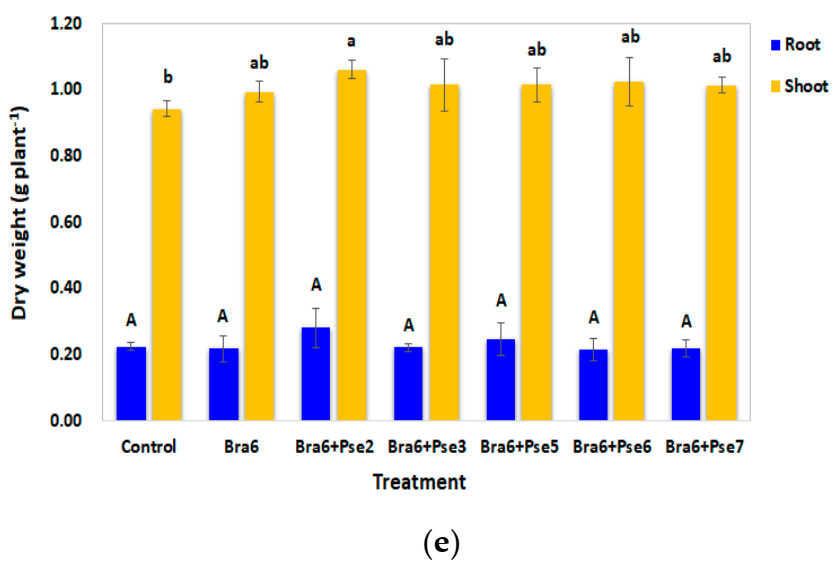
3.6. Influence of Co-Inoculation on Nodulation, Symbiotic Nitrogen Fixation, and Soybean Growth in Non-Sterile Strongly Acidic Soils Evaluated through Pot Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef] [PubMed]
- Spaink, H.P.; Kondorosi, A.; Hooykaas, P.J.J. The Rhizobiaceae; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998. [Google Scholar]
- Chen, W.F.; Wang, E.T.; Ji, Z.J.; Zhang, J.J. Recent development and new insight of diversification and symbiosis specificity of legume rhizobia: Mechanism and application. J. Appl. Microbiol. 2021, 131, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Suzaki, T.; Kawaguchi, M. Root nodulation: A developmental program involving cell fate conversion triggered by symbiotic bacterial infection. Curr. Opin. Plant. Biol. 2014, 21, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Clúa, J.; Roda, C.; Zanetti, M.E.; Blanco, F.A. Compatibility between legumes and rhizobia for the establishment of a successful nitrogen-fixing symbiosis. Genes 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemneh, A.A.; Zhou, Y.; Ryder, M.H.; Denton, M.D. Mechanisms in plant growth-promoting rhizobacteria that enhance legume-rhizobial symbioses. J. Appl. Microbiol. 2020, 129, 1133–1156. [Google Scholar] [CrossRef]
- Shin, C.-S.; Sharif, M.; Lee, H.-Y. Evaluating the effect of bacterial inoculation and fertilization on the soil nutrient status of coal mine soil by growing soybean (Glycine max) and shrub lespedeza (Lespedeza bicolor). Sustainability 2018, 10, 4793. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.-H.; Arafat, Y.; Letuma, P.; Ali, L.; Tayyab, M.; Waqas, M.; Li, Y.-C.; Lin, W.-W.; Lin, S.; Lin, W.-X. Restoration of long-term monoculture degraded tea orchard by green and goat manures application system. Sustainability 2019, 11, 1011. [Google Scholar] [CrossRef] [Green Version]
- Shen, F.T.; Lin, S.H. Priming effects of cover cropping on bacterial community in a tea plantation. Sustainability 2021, 13, 4345. [Google Scholar] [CrossRef]
- Zhang, B.; Du, N.; Li, Y.; Shi, P.; Wei, G. Distinct biogeographic patterns of rhizobia and non-rhizobial endophytes associated with soybean nodules across China. Sci. Total Environ. 2018, 643, 569–578. [Google Scholar] [CrossRef]
- Sharaf, H.; Rodrigues, R.R.; Moon, J.; Zhang, B.; Mills, K.; Williams, M.A. Unprecedented bacterial community richness in soybean nodules vary with cultivar and water status. Microbiome 2019, 7, 63. [Google Scholar] [CrossRef]
- Mayhood, P.; Mirza, B.S. Soybean root nodule and rhizosphere microbiome: Distribution of rhizobial and nonrhizobial endophytes. Appl. Environ. Microbiol. 2021, 87, e02884-20. [Google Scholar] [CrossRef]
- Sohn, S.-I.; Ahn, J.-H.; Pandian, S.; Oh, Y.-J.; Shin, E.-K.; Kang, H.-J.; Cho, W.-S.; Cho, Y.-S.; Shin, K.-S. Dynamics of bacterial community structure in the rhizosphere and root nodule of soybean: Impacts of growth stages and varieties. Int. J. Mol. Sci. 2021, 22, 5577. [Google Scholar] [CrossRef]
- Zeffa, D.M.; Fantin, L.H.; Koltun, A.; de Oliveira, A.L.M.; Nunes, M.P.B.A.; Canteri, M.G.; Gonçalves, L.S.A. Effects of plant growth-promoting rhizobacteria on co-inoculation with Bradyrhizobium in soybean crop: A meta-analysis of studies from 1987 to 2018. PeerJ 2020, 8, e7905. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.M. A Manual for the Practical Study of the Root-Nodule Bacteria; Blackwell Scientific: Oxford, UK, 1970. [Google Scholar]
- Huang, Y.S.; Shen, F.T. Bioprospecting of facultatively oligotrophic bacteria from non-rhizospheric soils. Appl. Soil Ecol. 2016, 108, 315–324. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Nzoué, A.; Miché, L.; Klonowska, A.; Laguerre, G.; de Lajudie, P.; Moulin, L. Multilocus sequence analysis of bradyrhizobia isolated from Aeschynomene species in Senegal. Syst. Appl. Microbiol. 2009, 32, 400–412. [Google Scholar] [CrossRef]
- Sterner, J.P.; Parker, M.A. Diversity and relationships of bradyrhizobia from Amphicarpaea bracteata based on partial nod and ribosomal sequences. Syst. Appl. Microbiol. 1999, 2, 387–392. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hung, M.H.; Arun, A.B.; Shen, F.T.; Rekha, P.D.; Young, C.C. Indigenous rhizobia associated with native shrubby legumes in Taiwan. Pedobiologia 2005, 49, 577–584. [Google Scholar] [CrossRef]
- Hardy, R.W.F.; Burns, R.C.; Holsten, R.D. Application of the acetylene reduction assay for measurement of nitrogen fixation. Soil Biol. Biochem. 1973, 5, 47–81. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Huang, C.M.; Chen, W.C.; Lin, S.H.; Wang, Y.N.; Shen, F.T. Exploration of root-associated bacteria from the medicinal plant Platycodon grandiflorum. Microbes Environ. 2019, 34, 413–420. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Kjeldahl, J. A new method for the determination of nitrogen in organic matter. Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Bremner, J.M. Nitrogen total. In Methods of Soil Analysis Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America Inc.: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Miller, R.O. Nitric-perchloric acid wet digestion in an open vessel. In Handbook of Reference Methods for Plant Analysis; Kalra, Y.P., Ed.; CRC Press: Washington, DC, USA, 1998; pp. 57–61. [Google Scholar]
- Zhang, Y.M.; Li, Y.; Chen, W.F.; Wang, E.T.; Tian, C.F.; Li, Q.Q.; Zhang, Y.Z.; Sui, X.H.; Chen, W.X. Biodiversity and biogeography of rhizobia associated with soybean plants grown in the North China Plain. Appl. Environ. Microbiol. 2011, 77, 6331–6342. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.F.; Zhou, Y.J.; Zhang, Y.M.; Li, Q.Q.; Zhang, Y.Z.; Li, D.F.; Wang, S.; Wang, J.; Gilbert, L.B.; Li, Y.R.; et al. Comparative genomics of rhizobia nodulating soybean suggests extensive recruitment of lineage-specific genes in adaptations. Proc. Natl. Acad. Sci. USA 2012, 109, 8629–8634. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Ma, Q.; Chen, Y.; Tian, B.; Xu, L.; Bai, Y.; Chen, W.; Li, X. Variation in rhizosphere microbial communities and its association with the symbiotic efficiency of rhizobia in soybean. ISME J. 2020, 14, 1915–1928. [Google Scholar] [CrossRef] [Green Version]
- Bakari, R.; Mungai, N.; Thuita, M.; Masso, C. Impact of soil acidity and lime on soybean (Glycine max) nodulation and nitrogen fixation in Kenyan soils. Acta Agric. Scand. Sect. B Soil Plant. Sci. 2020, 70, 667–678. [Google Scholar]
- Delamuta, J.R.M.; Ribeiro, R.A.; Ormeño-Orillo, E.; Melo, I.S.; Martínez-Romero, E.; Hungria, M. Polyphasic evidence supporting the reclassification of Bradyrhizobium japonicum group Ia strains as Bradyrhizobium diazoefficiens sp. nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 3342–3351. [Google Scholar] [CrossRef]
- Liu, L.; Chen, X.; Hu, S.; Zhan, Q.; Peng, W. Genetic diversity and distribution of rhizobia associated with soybean in red soil in Hunan Province. Arch. Microbiol. 2021, 203, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Tokgöz, S.; Lakshman, D.K.; Ghozlan, M.H.; Pinar, H.; Roberts, D.P.; Mitra, A. Soybean nodule-associated non-rhizobial bacteria inhibit plant pathogens and induce growth promotion in tomato. Plants 2020, 9, 1494. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, K.C.; Sharma, P.; Sirari, A.; Singh, I.; Gill, B.S.; Singh, U.; Saharan, K. Synergism of Pseudomonas aeruginosa (LSE-2) nodule endophyte with Bradyrhizobium sp. (LSBR-3) for improving plant growth, nutrient acquisition and soil health in soybean. World J. Microbiol. Biotechnol. 2019, 35, 47. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhou, X.; Smith, D.L. Enhanced soybean plant growth resulting from coinoculation of Bacillus strains with Bradyrhizobium japonicum. Crop. Sci. 2003, 43, 1774–1781. [Google Scholar] [CrossRef]
- Mishra, P.K.; Mishra, S.; Selvakumar, G.; Kundu, S.; Gupta, H.S. Enhanced soybean (Glycine max L.) plant growth and nodulation by Bradyrhizobium japonicum-SB1 in presence of Bacillus thuringiensis-KR1. Acta Agric. Scand. Sect. B Soil Plant. Sci. 2009, 59, 189–196. [Google Scholar]
- Jia, J.; Wang, X.; Deng, P.; Ma, L.; Baird, S.M.; Li, X.; Lu, S.E. Pseudomonas glycinae sp. nov. isolated from the soybean rhizosphere. Microbiologyopen 2020, 9, e1101. [Google Scholar] [CrossRef]
- Masciarelli, O.; Llanes, A.; Luna, V. A new PGPR co-inoculated with Bradyrhizobium japonicum enhances soybean nodulation. Microbiol. Res. 2014, 169, 609–615. [Google Scholar] [CrossRef]
- Subramanian, P.; Kim, K.; Krishnamoorthy, R.; Sundaram, S.; Sa, T. Endophytic bacteria improve nodule function and plant nitrogen in soybean on co-inoculation with Bradyrhizobium japonicum MN110. Plant. Growth Regul. 2015, 76, 327–332. [Google Scholar] [CrossRef]
- Sibponkrung, S.; Kondo, T.; Tanaka, K.; Tittabutr, P.; Boonkerd, N.; Yoshida, K.-I.; Teaumroong, N. Co-inoculation of Bacillus velezensis strain S141 and Bradyrhizobium strains promotes nodule growth and nitrogen fixation. Microorganisms 2020, 8, 678. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Romano, S.; Pocsfalvi, G.; Fiume, I.; Esposito, R.; Angelini, C.; Bianco, C. Bacterial IAA-delivery into Medicago root nodules triggers a balanced stimulation of C and N metabolism leading to a biomass increase. Microorganisms 2019, 7, 403. [Google Scholar] [CrossRef] [Green Version]
- Chebotar, V.K.; Asis, C.A.; Akao, S. Production of growth-promoting substances and high colonization ability of rhizobacteria enhance the nitrogen fixation of soybean when inoculated with Bradyrhizobium japonicum. Biol. Fertil. Soils 2001, 34, 427–432. [Google Scholar] [CrossRef]
- dos Santos Lima Fagotti, D.; Abrantes, J.L.F.; Cerezini, P.; Fukami, J.; Nogueira, M.A.; del Cerro, P.; Valderrama-Fernández, R.; Ollero, F.J.; Megías, M.; Hungria, M. Quorum sensing communication: Bradyrhizobium-Azospirillum interaction via N-acyl-homoserine lactones in the promotion of soybean symbiosis. J. Basic Microbiol. 2019, 59, 38–53. [Google Scholar] [CrossRef] [Green Version]
- Torres, D.; Benavidez, I.; Donadio, F.; Mongiardini, E.; Rosas, S.; Spaepen, S.; Vanderleyden, J.; Pěnčík, A.; Novák, O.; Strnad, M.; et al. New insights into auxin metabolism in Bradyrhizobium japonicum. Res. Microbiol. 2018, 169, 313–323. [Google Scholar] [CrossRef]
- Lucas-García, J.A.; Probanza, A.; Ramos, B.; Colón-Flores, J.J.; Gutiérrez-Mañero, F.J. Effects of plant growth promoting rhizobacteria (PGPRs) on the biological nitrogen fixation, nodulation and growth of Lupinus albus I. cv. Multolupa. Eng. Life Sci. 2004, 4, 71–77. [Google Scholar] [CrossRef]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, W.F.; Kloepper, J.W. Bacterial endophytes in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.N.; Yadav, N.; Dhaliwal, H.S.; Saxena, A.K. Endophytic microbes: Biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie Van Leeuwenhoek 2020, 113, 1075–1107. [Google Scholar] [CrossRef]
- Collmer, A.; Keen, N.T. The role of pectic enzymes in plant pathogenesis. Annu. Rev. Phytopathol. 1986, 24, 383–409. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Hit Taxon Name | Similarity | ||
|---|---|---|---|---|
| 16S rDNA | nifH | nodD | ||
| Bra4 | Bradyrhizobium elkanii USDA 76T | 1399/1399 (100%) | 734/737 (99.6%) | 328/329 (99.7%) |
| Bradyrhizobium pachyrhizi PAC 48T | 1399/1399 (100%) | 703/737 (95.4%) | 311/329 (94.5%) | |
| Bradyrhizobium tropiciagri SEMIA 6148T | 1399/1399 (100%) | 662/734 (90.2%) | 284/327 (86.9%) | |
| Bradyrhizobium brasilense UFLA 03-321T | 1399/1399 (100%) | 660/734 (89.9%) | NA | |
| Bradyrhizobium ripae WR4T | 1399/1399 (100%) | NA † | NA | |
| Bra6 | Bradyrhizobium diazoefficiens USDA 110T | 1394/1399 (99.6%) | 737/737 (100%) | 327/328 (99.7%) |
| Bradyrhizobium nanningense CCBAU 53390T | 1399/1399 (100%) | 687/737 (93.2%) | 310/328 (94.5%) | |
| Bradyrhizobium centrosematis A9T | 1399/1399 (100%) | 628/719 (87.3%) | NA | |
| Bradyrhizobium guangxiense CCBAU 53363T | 1262/1262 (100%) | 626/737 (84.9%) | NA | |
| Bra7 | Bradyrhizobium diazoefficiens USDA 110T | 1384/1384 (100%) | 737/737 (100%) | 327/328 (99.7%) |
| Bradyrhizobium niftali CNPSo 3448T | 1384/1384 (100%) | 669/737 (90.8%) | 303/328 (92.4%) | |
| Isolate | Hit Taxon Name | Similarity |
|---|---|---|
| Pse2 | Pseudomonas punonensis CECT 8089T | 1430/1436 (99.6%) |
| Pseudomonas straminea JCM 2783T | 1428/1435 (99.5%) | |
| Pseudomonas argentinensis CH01T | 1428/1435 (99.5%) | |
| Pse3 | Pseudomonas mucoides P154aT | 1221/1232 (99.1%) |
| Pseudomonas bijieensis L22-9T | 1420/1435 (99.0%) | |
| Pseudomonas gessardii DSM 17152T | 1422/1439 (98.8%) | |
| Pseudomonas mediterranea CFBP 5447T | 1422/1439 (98.8%) | |
| Pse5 | Pseudomonas atacamensis M7D1T | 1448/1450 (99.9%) |
| Pseudomonas koreensis Ps 9-14T | 1447/1450 (99.8%) | |
| Pse6 | Pseudomonas glycinae MS586T | 1450/1450 (100%) |
| Pseudomonas kribbensis 46-2T | 1448/1450 (99.9%) | |
| Pse7 | Pseudomonas glycinae MS586T | 1450/1450 (100%) |
| Pseudomonas kribbensis 46-2T | 1448/1450 (99.9%) |
| Treatment | Root Length (cm) | Shoot Length (cm) | Number of Leaves | Number of Nodules | Nitrogen-Fixing Activity (nmol Ethylene h−1 Nodule−1) |
|---|---|---|---|---|---|
| Control † | 12.25 ± 3.66 a ‡ | 14.00 ± 3.34 a | 17 ± 3 a | 1 ± 1 c | 0.10 |
| Bra4 | 13.25 ± 1.60 a | 12.77 ± 2.98 a | 11 ± 1 b | 5 ± 2 a | 15.47 |
| Bra6 | 12.25 ± 2.50 a | 13.00 ± 4.10 a | 13 ± 2 b | 3 ± 2 bc | 29.38 |
| Bra7 | 13.83 ± 1.81 a | 13.33 ± 1.03 a | 11 ± 1 b | 4 ± 1 ab | 8.47 |
| Isolate | Nitrogen-Fixing Activity (nmol Ethylene Tube−1 h−1) | Phosphate-Solubilizing Activity (µg mL−1) | Production of Siderophore (Halo Diameter/Colony Diameter) | Production of IAA in Tryptophan-Containing YEM (µg mL−1) | |
|---|---|---|---|---|---|
| pH 5 | pH 7 | ||||
| Bra6 | 0.337 | ND † | ND | ND | ND |
| Pse2 | 0.091 | 311.33 | 1.11 | 3.30 | 32.91 |
| Pse3 | 0.040 | 303.52 | 1.26 | 3.22 | 35.11 |
| Pse5 | 0.081 | 1391.43 | 1.24 | 8.70 | 73.76 |
| Pse6 | 0.053 | 1212.86 | 1.26 | 8.61 | 78.29 |
| Pse7 | 0.027 | 1230.71 | 1.29 | 11.22 | 80.24 |
| Treatment | N | P | K | Ca | Mg | S | Fe | Mn | Cu | Zn | B | Na | Al |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mg plant−1 | |||||||||||||
| Control † | 4.8 ± 0.7 a ‡ | 0.9 ± 0.1 b | 1.9 ± 0.3 c | 0.5 ± 0.1 c | 0.5 ± 0.1 b | 0.5 ± 0.1 c | 0.027 ± 0.004 bc | 0.003 ± 0.000 c | 0.002 ± 0.000 bc | 0.006 ± 0.001 b | 0.008 ± 0.001 c | 0.4 ± 0.1 a | 0.014 ± 0.002 b |
| Bra6 | 4.8 ± 0.1 a | 0.9 ± 0.0 b | 2.3 ± 0.0 b | 0.6 ± 0.0 bc | 0.6 ± 0.0 b | 0.6 ± 0.0 b | 0.029 ± 0.001 ab | 0.004 ± 0.000 bc | 0.002 ± 0.000 c | 0.006 ± 0.000 b | 0.008 ± 0.000 bc | 0.3 ± 0.0 b | 0.016 ± 0.000 a |
| Bra6 + Pse2 | 4.5 ± 0.1 ab | 1.2 ± 0.0 a | 2.4 ± 0.1 ab | 0.7 ± 0.0 a | 0.7 ± 0.0 a | 0.7 ± 0.0 a | 0.031 ± 0.001 a | 0.004 ± 0.000 ab | 0.002 ± 0.000 ab | 0.007 ± 0.000 a | 0.011 ± 0.000 a | 0.4 ± 0.0 a | 0.017 ± 0.001 a |
| Bra6 + Pse3 | 4.8 ± 0.2 a | 1.2 ± 0.1 a | 2.7 ± 0.1 a | 0.7 ± 0.0 a | 0.7 ± 0.0 a | 0.6 ± 0.0 ab | 0.024 ± 0.001 c | 0.004 ± 0.000 ab | 0.002 ± 0.000 a | 0.007 ± 0.000 a | 0.011 ± 0.001 a | 0.4 ± 0.0 a | 0.013 ± 0.001 b |
| Bra6 + Pse5 | 4.8 ± 0.7 a | 1.0 ± 0.2 b | 2.5 ± 0.4 ab | 0.7 ± 0.1 a | 0.7 ± 0.1 a | 0.6 ± 0.1 bc | 0.019 ± 0.003 d | 0.004 ± 0.001 a | 0.001 ± 0.000 c | 0.008 ± 0.001 a | 0.009 ± 0.001 b | 0.3 ± 0.0 b | 0.011 ± 0.002 c |
| Bra6 + Pse6 | 4.0 ± 0.4 bc | 1.0 ± 0.1 ab | 2.4 ± 0.2 ab | 0.7 ± 0.1 ab | 0.7 ± 0.1 a | 0.6 ± 0.1 b | 0.017 ± 0.002 d | 0.003 ± 0.000 c | 0.002 ± 0.000 c | 0.006 ± 0.001 b | 0.009 ± 0.001 b | 0.4 ± 0.0 a | 0.010 ± 0.001 c |
| Bra6 + Pse7 | 3.6 ± 0.4 c | 0.6 ± 0.6 c | 1.4 ± 0.1 d | 0.4 ± 0.0 d | 0.4 ± 0.0 c | 0.3 ± 0.0 d | 0.012 ± 0.001 e | 0.002 ± 0.000 d | 0.001 ± 0.000 d | 0.003 ± 0.000 c | 0.005 ± 0.001 d | 0.2 ± 0.0 c | 0.008 ± 0.001 d |
| Treatment | N | P | K | Ca | Mg | S | Fe | Mn | Cu | Zn | B | Na | Al |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mg plant−1 | |||||||||||||
| Control † | 8.3 ± 1.3 c ‡ | 2.0 ± 0.1 ab | 9.6 ± 0.3 e | 6.0 ± 0.2 c | 2.7 ± 0.1 c | 2.7 ± 0.1 ab | 0.161 ± 0.004 a | 0.426 ± 0.011 d | 0.011 ± 0.000 bc | 0.055 ± 0.001 bc | 0.032 ± 0.001 de | 0.2 ± 0.0 d | 0.194 ± 0.005 a |
| Bra6 | 19.0 ± 0.3 b | 2.1 ± 0.1 a | 12.4 ± 0.4 bc | 7.3 ± 0.2 b | 3.0 ± 0.1 bc | 2.6 ± 0.1 b | 0.159 ± 0.005 a | 0.559 ± 0.017 b | 0.012 ± 0.000 a | 0.059 ± 0.002 b | 0.041 ± 0.001 a | 0.2 ± 0.0 cd | 0.158 ± 0.005 bc |
| Bra6 + Pse2 | 24.1 ± 0.8 a | 2.1 ± 0.1 a | 13.9 ± 0.3 a | 8.1 ± 0.2 a | 3.3 ± 0.1 a | 2.8 ± 0.7 a | 0.137 ± 0.003 b | 0.605 ± 0.015 a | 0.012 ± 0.000 a | 0.067 ± 0.002 a | 0.036 ± 0.001 bc | 0.2 ± 0.0 b | 0.169 ± 0.004 b |
| Bra6 + Pse3 | 18.9 ± 3.8 b | 2.0 ± 0.2 ab | 12.8 ± 1.0 b | 7.1 ± 0.6 b | 3.1 ± 0.2 ab | 2.6 ± 0.2 ab | 0.099 ± 0.008 e | 0.498 ± 0.039 c | 0.012 ± 0.001 ab | 0.058 ± 0.004 b | 0.034 ± 0.003 bcd | 0.2 ± 0.0 bc | 0.123 ± 0.009 f |
| Bra6 + Pse5 | 20.5 ± 1.2 ab | 1.7 ± 0.1 c | 10.8 ± 0.5 d | 6.9 ± 0.3 b | 2.9 ± 0.1 bc | 2.1 ± 0.1 c | 0.111 ± 0.005 d | 0.514 ± 0.025 bc | 0.009 ± 0.000 d | 0.055 ± 0.003 bc | 0.033 ± 0.002 cd | 0.2 ± 0.0 cd | 0.139 ± 0.007 de |
| Bra6 + Pse6 | 20.7 ± 2.7 ab | 1.8 ± 0.1 c | 11.5 ± 0.8 cd | 7.0 ± 0.5 b | 2.9 ± 0.2 bc | 2.3 ± 0.2 c | 0.111 ± 0.008 d | 0.508 ± 0.036 c | 0.012 ± 0.001 a | 0.052 ± 0.004 c | 0.029 ± 0.002 e | 0.3 ± 0.0 a | 0.131 ± 0.009 ef |
| Bra6 + Pse7 | 19.6 ± 2.2 b | 1.9 ± 0.0 bc | 11.5 ± 0.3 cd | 6.8 ± 0.2 b | 2.9 ± 0.1 bc | 2.3 ± 0.1 c | 0.122 ± 0.003 c | 0.538 ± 0.013 bc | 0.010 ± 0.000 c | 0.058 ± 0.001 b | 0.037 ± 0.001 b | 0.2 ± 0.0 d | 0.150 ± 0.004 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, T.-C.; Lin, S.-H.; Shen, F.-T. Enhancing Symbiotic Nitrogen Fixation and Soybean Growth through Co-Inoculation with Bradyrhizobium and Pseudomonas Isolates. Sustainability 2021, 13, 11539. https://doi.org/10.3390/su132011539
Tu T-C, Lin S-H, Shen F-T. Enhancing Symbiotic Nitrogen Fixation and Soybean Growth through Co-Inoculation with Bradyrhizobium and Pseudomonas Isolates. Sustainability. 2021; 13(20):11539. https://doi.org/10.3390/su132011539
Chicago/Turabian StyleTu, Ting-Chen, Shih-Han Lin, and Fo-Ting Shen. 2021. "Enhancing Symbiotic Nitrogen Fixation and Soybean Growth through Co-Inoculation with Bradyrhizobium and Pseudomonas Isolates" Sustainability 13, no. 20: 11539. https://doi.org/10.3390/su132011539
APA StyleTu, T. -C., Lin, S. -H., & Shen, F. -T. (2021). Enhancing Symbiotic Nitrogen Fixation and Soybean Growth through Co-Inoculation with Bradyrhizobium and Pseudomonas Isolates. Sustainability, 13(20), 11539. https://doi.org/10.3390/su132011539