
Bio-Flocculation Property Analyses of Oleaginous Microalgae Auxenochlorella protothecoides UTEX 2341



Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Growth Conditions
2.2. The Details of Freeze-Dried Algae Samples for Bio-Flocculation Analysis
2.3. Evaluation of Microalgae Flocculation Ability
2.3.1. The Self-Flocculation Analysis of Algae
2.3.2. Bio-Flocculation Analysis of Freeze-Dried Algae Powder
2.4. The Composition Analysis of Flocculant
2.5. The Analysis of Biomass and Lipid Content of UTEX 2341
2.6. Statistical Analysis
3. Results and Discussion
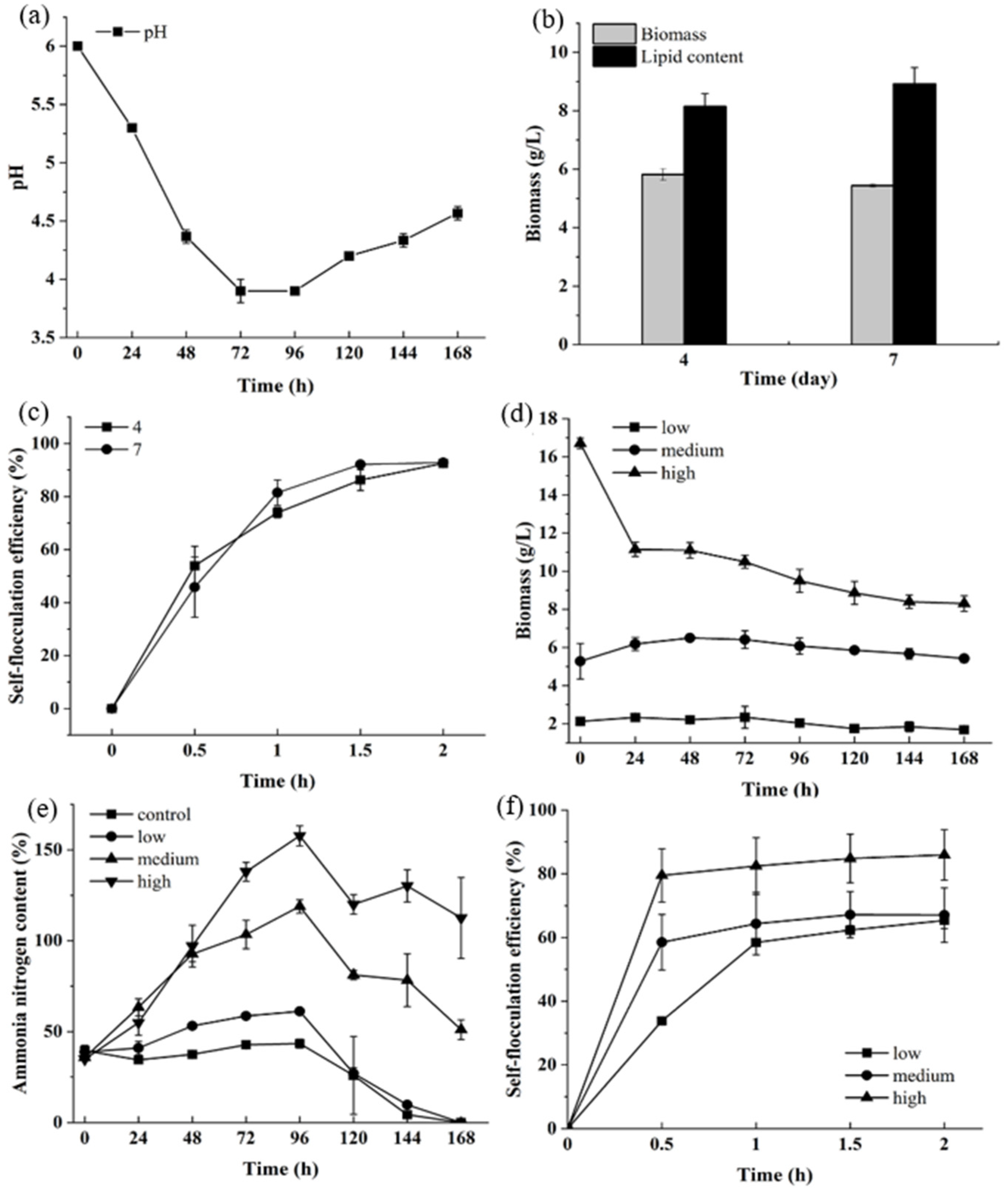
3.1. The Growth and Self-Flocculation Analysis of UTEX 2341 Cultured in Heterotrophic Medium and Municipal Wastewater
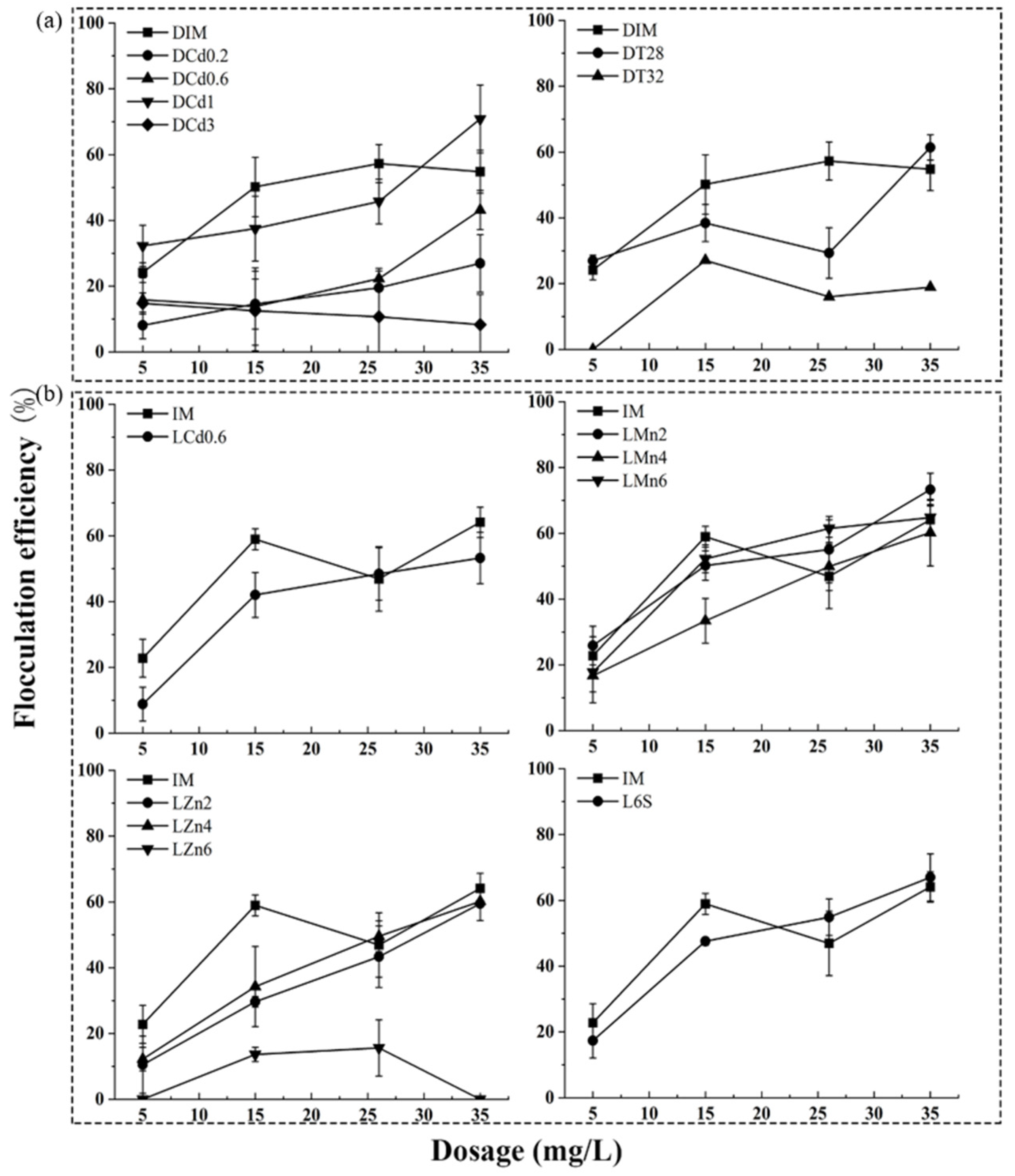
3.2. Bio-Flocculation Ability Analysis of Freeze-Dried Algae Powder of UTEX 2341
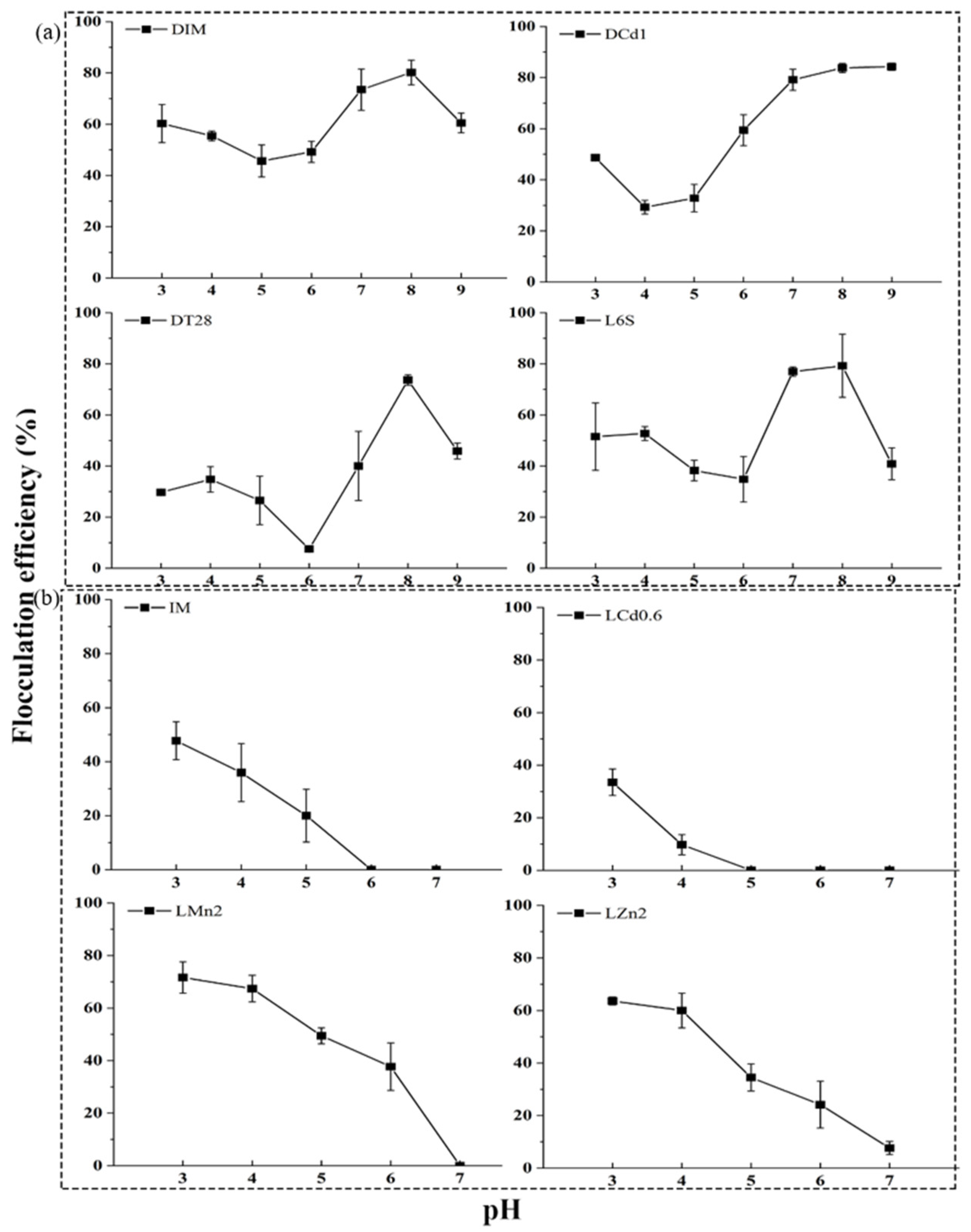
3.3. Effects of pH on Flocculation Efficiency
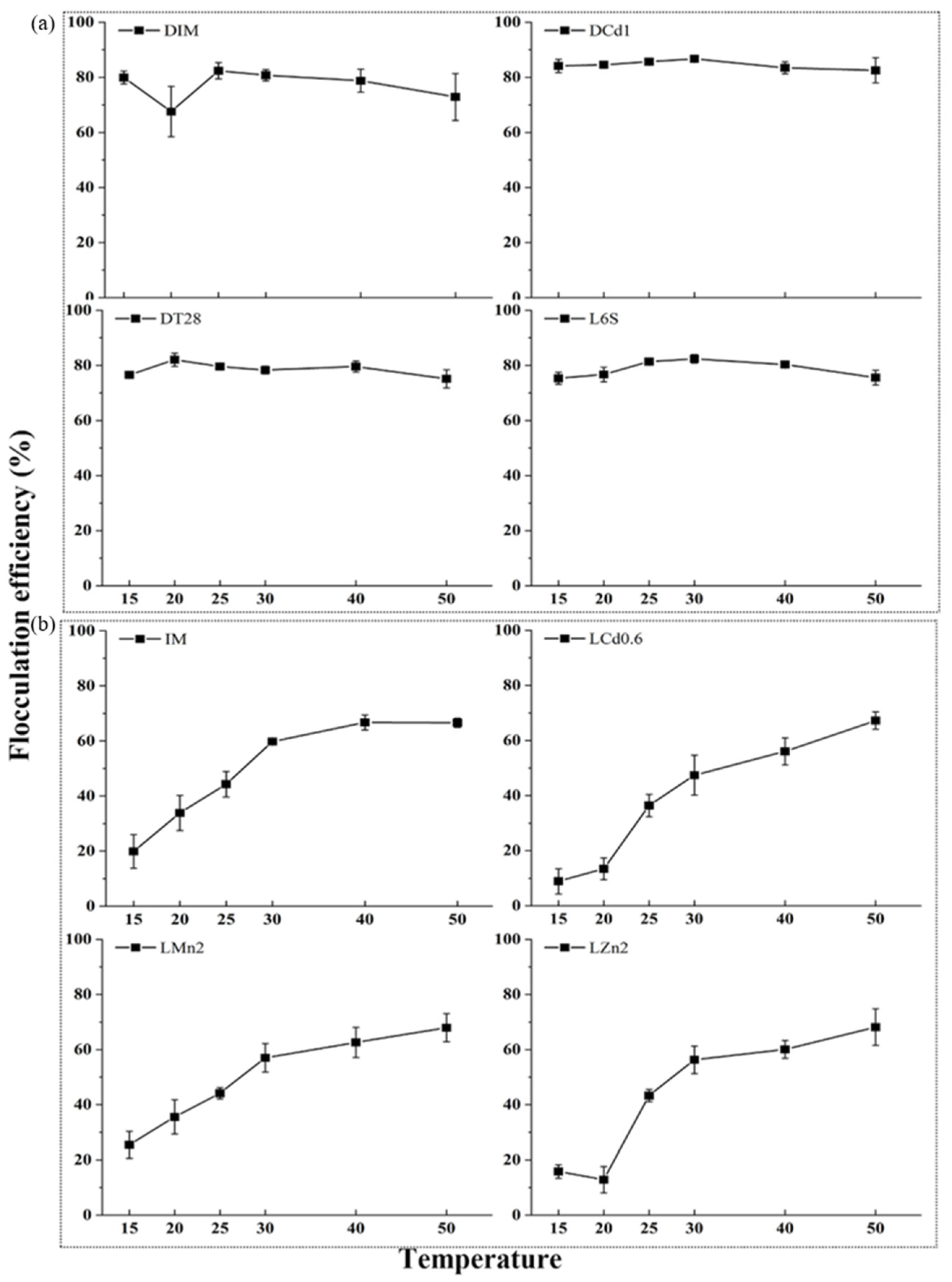
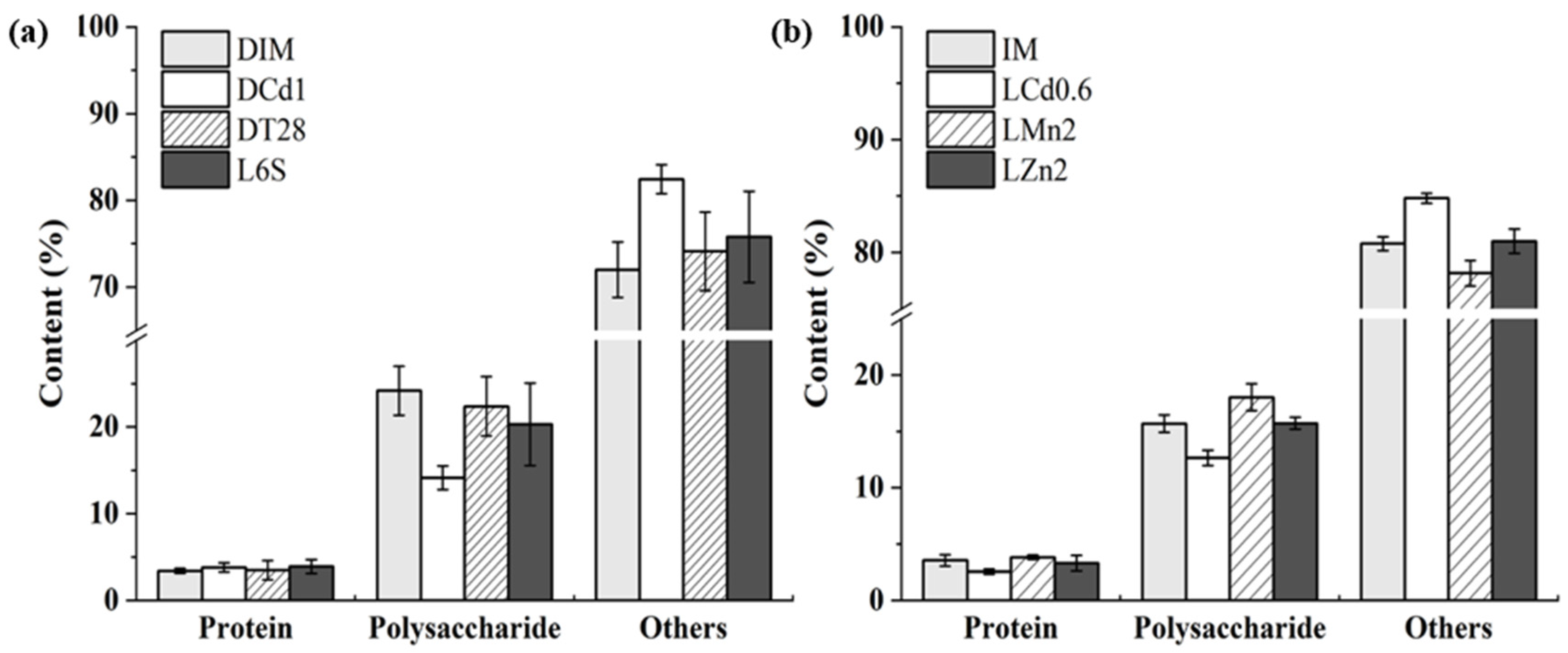
3.4. Effects of Temperature on Flocculation Efficiency and the Analysis of Flocculant Composition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, M.; Sun, Y.; Rathour, R.; Pandey, A.; Thakur, I.S.; Tsang, D.C.W. Algae as potential feedstock for the production of biofuels and value-added products: Opportunities and challenges. Sci. Total Environ. 2020, 716, 137116. [Google Scholar] [CrossRef]
- Marinello, S.; Gamberini, R.; Bastone, L. Experimental plant for the cultivation of microalgae in photobioreactors for energy production. In Proceedings of the 24th Summer School Francesco Turco, Brescia, Italy, 11–13 September 2019. [Google Scholar]
- Liu, J.; Zhu, Y.; Tao, Y.; Zhang, Y.; Li, A.; Li, T.; Sang, M.; Zhang, C. Freshwater microalgae harvested via flocculation induced by pH decrease. Biotechnol. Biofuels. 2013, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, A.; García-Galán, M.J.; García, J.; Díez-Montero, R. Optimization and operation of a demonstrative full scale microalgae harvesting unit based on coagulation, flocculation and sedimentation. Sep. Purif. Technol. 2021, 259, 118171. [Google Scholar] [CrossRef]
- Vasistha, S.; Khanra, A.; Clifford, M.; Rai, M.P. Current advances in microalgae harvesting and lipid extraction processes for improved biodiesel production: A review. Renew. Sustain. Energy Rev. 2021, 137, 110498. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Y.; Liu, L.; Jiang, X.; Zhang, K.; Zheng, T.; Wang, H. First evidence of bioflocculant from Shinella albus with flocculation activity on harvesting of Chlorella vulgaris biomass. Bioresour. Technol. 2016, 218, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ryu, B.G.; Kim, K.; Kim, B.K.; Han, J.I.; Yang, J.W. Continuous microalgae recovery using electrolysis: Effect of different electrode pairs and timing of polarity exchange. Bioresour. Technol. 2012, 123, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, D.; Foubert, I.; Muylaert, K. Flocculation as a low-cost method for harvesting microalgae for bulk biomass production. Trends Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.C.; Kim, B.; Farooq, W.; Chung, J.; Han, J.I.; Shin, H.J.; Jeong, S.H.; Park, J.Y.; Lee, J.S.; Oh, Y.K. Harvesting of oleaginous Chlorella sp. by organoclays. Bioresour. Technol. 2013, 132, 440–445. [Google Scholar] [CrossRef]
- Schenk, P.M.; Thomas-Hall, S.R.; Stephens, E.; Marx, U.C.; Mussgnug, J.H.; Posten, C.; Kruse, O.; Hankamer, B. Second Generation Biofuels: High-Efficiency Microalgae for Biodiesel Production. Bioenergy Res. 2008, 1, 20–43. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Harrison, S.T.L. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Appl. Phycol. 2009, 21, 493–507. [Google Scholar] [CrossRef]
- Salim, S.; Vermue, M.H.; Wijffels, R.H. Ratio between autoflocculating and target microalgae affects the energy-efficient harvesting by bio-flocculation. Bioresour. Technol. 2012, 118, 49–55. [Google Scholar] [CrossRef]
- Wang, H.; Laughinghouse, H.D.T.; Anderson, M.A.; Chen, F.; Willliams, E.; Place, A.R.; Zmora, O.; Zohar, Y.; Zheng, T.; Hill, R.T. Novel bacterial isolate from Permian groundwater, capable of aggregating potential biofuel-producing microalga Nannochloropsis oceanica IMET1. Appl. Environ. Microbiol. 2012, 78, 1445–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Hu, B. A novel method to harvest microalgae via co-culture of filamentous fungi to form cell pellets. Bioresour. Technol. 2012, 114, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Xu, W.; Luan, T.; Pan, T.; Yang, L.; Lin, L. Comparative responses of cell growth and related extracellular polymeric substances in Tetraselmis sp. to nonylphenol, bisphenol A and 17α-ethinylestradiol. Environ. Pollut. 2021, 274, 116605. [Google Scholar] [CrossRef]
- Rashid, N.; Nayak, M.; Lee, B.; Chang, Y.-K. Efficient microalgae harvesting mediated by polysaccharides interaction with residual calcium and phosphate in the growth medium. J. Clean. Prod. 2019, 234, 150–156. [Google Scholar] [CrossRef]
- Ummalyma, S.B.; Gnansounou, E.; Sukumaran, R.K.; Sindhu, R.; Pandey, A.; Sahoo, D. Bioflocculation: An alternative strategy for harvesting of microalgae—An overview. Bioresour. Technol. 2017, 242, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yuan, H.; Li, B.; Yang, J. Significance evaluation of the effects of environmental factors on the lipid accumulation of Chlorella minutissima UTEX 2341 under low-nutrition heterotrophic condition. Bioresour. Technol. 2014, 152, 177–184. [Google Scholar] [CrossRef]
- Xing, G.; Yuan, H.; Yang, J.; Li, J.; Gao, Q.; Li, W.; Wang, E. Integrated analyses of transcriptome, proteome and fatty acid profilings of the oleaginous microalga Auxenochlorella protothecoides UTEX 2341 reveal differential reprogramming of fatty acid metabolism in response to low and high temperatures. Algal Res. 2018, 33, 16–27. [Google Scholar] [CrossRef]
- Lu, J.; Ma, Y.; Xing, G.; Li, W.; Kong, X.; Li, J.; Wang, L.; Yuan, H.; Yang, J. Revelation of microalgae’s lipid production and resistance mechanism to ultra-high Cd stress by integrated transcriptome and physiochemical analyses. Environ. Pollut. 2019, 250, 186–195. [Google Scholar] [CrossRef]
- Li, C.; Yang, J.; Wang, X.; Wang, E.; Li, B.; He, R.; Yuan, H. Removal of nitrogen by heterotrophic nitrification-aerobic denitrification of a phosphate accumulating bacterium Pseudomonas stutzeri YG-24. Bioresour. Technol. 2015, 182, 18–25. [Google Scholar] [CrossRef]
- Liu, W.; Wang, K.; Li, B.; Yuan, H.; Yang, J. Production and characterization of an intracellular bioflocculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresour. Technol. 2010, 101, 1044–1048. [Google Scholar] [CrossRef]
- Albalasmeh, A.A.; Berhe, A.A.; Ghezzehei, T.A. A new method for rapid determination of carbohydrate and total carbon concentrations using UV spectrophotometry. Carbohydr. Polym. 2013, 97, 253–261. [Google Scholar] [CrossRef]
- Zhu, N.; Liu, J.; Yang, J.; Lin, Y.; Yang, Y.; Ji, L.; Li, M.; Yuan, H. Comparative analysis of the secretomes of Schizophyllum commune and other wood-decay basidiomycetes during solid-state fermentation reveals its unique lignocellulose-degrading enzyme system. Biotechnol. Biofuels 2016, 9, 2–22. [Google Scholar] [CrossRef] [Green Version]
- Halfhide, T.; Dalrymple, O.K.; Wilkie, A.C.; Trimmer, J.; Gillie, B.; Udom, I.; Zhang, Q.; Ergas, S.J. Growth of an Indigenous Algal Consortium on Anaerobically Digested Municipal Sludge Centrate: Photobioreactor Performance and Modeling. Bioenergy Res. 2014, 8, 249–258. [Google Scholar] [CrossRef]
- Wang, M.; Yang, H.; Ergas, S.J.; van der Steen, P. A novel shortcut nitrogen removal process using an algal-bacterial consortium in a photo-sequencing batch reactor (PSBR). Water Res. 2015, 87, 38–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Guo, W.; Yen, H.-W.; Ho, S.-H.; Lo, Y.-C.; Cheng, C.-L.; Ren, N.; Chang, J.-S. Cultivation of Chlorella vulgaris JSC-6 with swine wastewater for simultaneous nutrient/COD removal and carbohydrate production. Bioresour. Technol. 2015, 198, 619–625. [Google Scholar] [CrossRef]
- Zuharlida, T.H.; Fatimah, M.Y.; Mohd, S.M.; Mohamed, S.M.D.; Arbakariya, B.A. Effect of different flocculants on the flocculation performance of microalgae, Chaetoceros calcitrans, cells. Afr. J. Biotechnol. 2009, 8, 5971–5978. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Zhang, R.; Cheng, L.; Chi, Z. A recycling culture of Neochloris oleoabundans in a bicarbonate-based integrated carbon capture and algae production system with harvesting by auto-flocculation. Biotechnol. Biofuels 2018, 11, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, L.; Salgueiro, J.L.; Maceiras, R.; Cancela, Á.; Sánchez, Á. An effective method for harvesting of marine microalgae: pH induced flocculation. Biomass Bioenergy 2017, 97, 20–26. [Google Scholar] [CrossRef]
- Ndikubwimana, T.; Zeng, X.; Murwanashyaka, T.; Manirafasha, E.; He, N.; Shao, W.; Lu, Y. Harvesting of freshwater microalgae with microbial bioflocculant: A pilot-scale study. Biotechnol. Biofuels 2016, 9, 47. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.L.; Rand, J.D.; Caldwell, G.S. Treatment with algae extracts promotes flocculation, and enhances growth and neutral lipid content in Nannochloropsis oculata--a candidate for biofuel production. Mar. Biotechnol. 2012, 14, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Lin, J.; Xia, M.; Luo, J.; Du, B.; Pang, H.; Yan, Z. Impact of Extracellular Polymeric Substance in the Inactivation of Harmful Algae by Ag2O/g-C3N4 under Visible Light. Part. Part. Syst. Charact. 2020, 38, 2000272. [Google Scholar] [CrossRef]
- Pirhanov, A.; Bridges, C.M.; Goodwin, R.A.; Guo, Y.-S.; Furrer, J.; Shor, L.M.; Gage, D.J.; Cho, Y.K. Optogenetics in Sinorhizobium meliloti Enables Spatial Control of Exopolysaccharide Production and Biofilm Structure. ACS Synth. Biol. 2021, 10, 345–356. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Ballesteros, M. Microalgae autoflocculation: An alternative to high-energy consuming harvesting methods. J. Appl. Phycol. 2012, 25, 991–999. [Google Scholar] [CrossRef]
- Hanumanth Rao, N.R.; Yap, R.; Whittaker, M.; Stuetz, R.M.; Jefferson, B.; Peirson, W.L.; Granville, A.M.; Henderson, R.K. The role of algal organic matter in the separation of algae and cyanobacteria using the novel “Posi”—Dissolved air flotation process. Water Res. 2018, 130, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, H.-M.; lEE, S.J.; Park, M.H.; Kim, H.S. Harvesting of Chlorella vulgaris using a bioflocculant from Paenibacillus sp. AM49. Biotechnol. Lett. 2001, 23, 1229–1234. [Google Scholar] [CrossRef]
- Díaz-Santos, E.; Vila, M.; de la Vega, M.; León, R.; Vigara, J. Study of bioflocculation induced by Saccharomyces bayanus var. uvarum and flocculating protein factors in microalgae. Algal Res. 2015, 8, 23–29. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, Y.; Huang, W.; Zhang, C.; Li, T.; Zhang, Y.; Li, A. Evaluation of flocculation induced by pH increase for harvesting microalgae and reuse of flocculated medium. Bioresour. Technol. 2012, 110, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Agbakpe, M.; Ge, S.; Zhang, W.; Zhang, X.; Kobylarz, P. Algae harvesting for biofuel production: Influences of UV irradiation and polyethylenimine (PEI) coating on bacterial biocoagulation. Bioresour. Technol. 2014, 166, 266–272. [Google Scholar] [CrossRef]
- Takagi, H.; Kadowaki, K. Flocculant Production by Paecilomyces sp. Taxonomic Studies and Culture Conditions for Production. Agric. Biol. Chem. 2014, 49, 3151–3157. [Google Scholar] [CrossRef] [Green Version]
- Andreadakis, A.D. Physical and chemical properties of activated sludge floc. Water Res. 1993, 27, 1707–1714. [Google Scholar] [CrossRef]
- Shen, Y.; Fan, Z.; Chen, C.; Xu, X. An auto-flocculation strategy for Chlorella vulgaris. Biotechnol. Lett. 2015, 37, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.L.; Zhao, X.Q.; Wan, C.; Huang, Z.Y.; Yang, Y.L.; Alam, M.A.; Ho, S.H.; Bai, F.W.; Chang, J.S. Characterization of flocculating agent from the self-flocculating microalga Scenedesmus obliquus AS-6-1 for efficient biomass harvest. Bioresour. Technol. 2013, 145, 285–289. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae Species | Flocculating Rate | Flocculation Time (h) | References |
|---|---|---|---|
| A. protothecoides UTEX 2341 | 92.82% (IM) | 2 | This study |
| 85% (municipal wastewater) | 2 | This study | |
| Chaetoceros minutissimus | 82% | 4 | [28] |
| C. vulgaris | 25% | 3 | [12] |
| N. oleoabundans (Alkaline flocculation added) | 40% | 3 | [12] |
| Ettlia sp. | 61% | 1 | [16] |
| Neochloris oleoabundans | 97.4% | 2 | [29] |
| Skeletonema Costatum (pH = 10) | 65% | 7.5 | [30] |
| Desmodesmus sp. F51 | 60–89% | 3 | [31] |
| Skeletonema marinoi to N. oculata | 95% | 6 | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Li, B.; Yang, J. Bio-Flocculation Property Analyses of Oleaginous Microalgae Auxenochlorella protothecoides UTEX 2341. Sustainability 2021, 13, 2885. https://doi.org/10.3390/su13052885
Li J, Li B, Yang J. Bio-Flocculation Property Analyses of Oleaginous Microalgae Auxenochlorella protothecoides UTEX 2341. Sustainability. 2021; 13(5):2885. https://doi.org/10.3390/su13052885
Chicago/Turabian StyleLi, Jinyu, Baozhen Li, and Jinshui Yang. 2021. "Bio-Flocculation Property Analyses of Oleaginous Microalgae Auxenochlorella protothecoides UTEX 2341" Sustainability 13, no. 5: 2885. https://doi.org/10.3390/su13052885
APA StyleLi, J., Li, B., & Yang, J. (2021). Bio-Flocculation Property Analyses of Oleaginous Microalgae Auxenochlorella protothecoides UTEX 2341. Sustainability, 13(5), 2885. https://doi.org/10.3390/su13052885